 +1 credit
+1 credit
Video Upload Options
After sperm enter the female reproductive tract, the physicochemical and biochemical microenvironment undergoes significant changes. In particular, the large changes in various ions encountered by sperm may alter the physiology of sperm, ultimately compromising capacitation and fertilization. Thus, the rapid response to environmental variations is vital for sperm functions. For example, Calcium, the most crucial ion for sperm functions, enters into sperm via Ca2+ permeable ion channels. The cation channel of sperm (CatSper) is a sperm-specific, pH-sensitive, and Ca2+-permeable ion channel. It is responsible for the predominant Ca2+ entry in mammalian sperm and is involved in nearly every event of sperm to acquire fertilizing capability. In addition, CatSper also serves as a pivotal polymodal chemosensor in mammalian sperm by responding to multiple chemical cues. Physiological chemicals (such as progesterone, prostaglandins, β-defensins, and odorants) provoke Ca2+ entry into sperm by activating CatSper and thus triggering sperm functions.
1. Introduction
Specifically, the cation channel of sperm (CatSper) is a sperm-specific, pH-sensitive, and Ca2+-permeable ion channel [7]. Crucially, this channel is responsible for the predominant Ca2+ entry in mammalian sperm and is involved in nearly every event by which sperm acquire their fertilizing capability. Additionally, ion channels are capable of transporting ions faster than transporters. This allows sperm to respond quickly to guidance cues within the female reproductive tract. Consequently, CatSper enables the translation of large changes in the microenvironment into changes of [Ca2+]i [8]. Although fertilization is at the center of creating new life, it is still a long way from being fully understood. A better understanding of the CatSper channel is important, not only to advance knowledge of the cause of male infertility but also to inspire improvement in the development of male contraceptives. On one hand, the knockout of genes encoding the CatSper channel in male mice, as well as genetic mutations in CatSper genes in humans, lead to male infertility and the inability of sperm to undergo hyperactivation and to penetrate oocytes. On the other hand, CatSper plays a pivotal role in responding to multiple chemical cues, including physiological chemicals (such as progesterone [P4] and prostaglandins [PGs]), and synthetic and natural chemicals (such as medicines and endocrine disrupting chemicals [EDCs]). Therefore, CatSper is also a pivotal polymodal chemosensor in mammalian sperm [9].
2. CatSper
3. CatSper and Physiological Chemicals
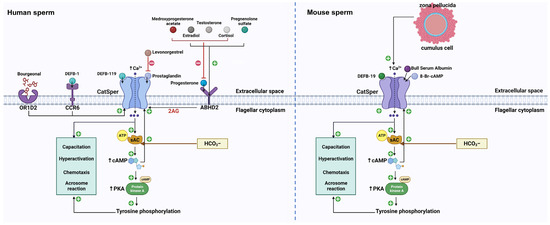
3.1. CatSper and Endogenous Steroids
3.2. CatSper and PGs
3.3. CatSper and cAMP
3.4. CatSper and ZP Glycoproteins
3.5. CatSper and Bovine Serum Albumin (BSA)
3.6. CatSper and DEFBs
3.7. CatSper and Neurotransmitters
3.8. CatSper and Odorant Attractants
4. CatSper and Medicines
4.1. CatSper and Traditional Medicine
4.2. CatSper and Anti-Depressants
4.3. CatSper and 5-Alpha Reductase Inhibitors
4.4. CatSper and Analgesics
4.5. CatSper and Ca2+ Channel Blockers
4.6. CatSper and Phosphodiesterase (PDE)-Inhibitors
4.7. CatSper and Anti-Inflammatory Drugs
5. CatSper and EDCs
5.1. CatSper and Environmental Estrogens
5.2. CatSper and Pesticides
5.3. CatSper and Chemical Ultraviolet (UV) Filters
6. CatSper and Drugs of Abuse
7. CatSper and Antioxidants
References
- Gervasi, M.G.; Visconti, P.E. Molecular changes and signaling events occurring in spermatozoa during epididymal maturation. Andrology 2017, 5, 204–218.
- TMortimer, S. A critical review of the physiological importance and analysis of sperm movement in mammals. Hum. Reprod. 1997, 3, 403–439.
- Perez-Cerezales, S.; Boryshpolets, S.; Eisenbach, M. Behavioral mechanisms of mammalian sperm guidance. Asian J. Androl. 2015, 17, 628–632.
- Costello, S.; Michelangeli, F.; Nash, K.; Lefievre, L.; Morris, J.; Machado-Oliveira, G.; Barratt, C.; Kirkman-Brown, J.; Publicover, S. Ca2+-stores in sperm: Their identities and functions. Reproduction 2009, 138, 425–437.
- Molina, L.C.P.; Luque, G.M.; Balestrini, P.A.; Marín-Briggiler, C.I.; Romarowski, A.; Buffone, M.G. Molecular Basis of Human Sperm Capacitation. Front. Cell Dev. Biol. 2018, 6, 72.
- Finkelstein, M.; Etkovitz, N.; Breitbart, H. Ca(2+) signaling in mammalian spermatozoa. Mol. Cell. Endocrinol. 2020, 516, 110953.
- Singh, A.P.; Rajender, S. CatSper channel, sperm function and male fertility. Reprod. Biomed. Online 2015, 30, 28–38.
- Jeschke, J.K.; Biagioni, C.; Schierling, T.; Wagner, I.V.; Börgel, F.; Schepmann, D.; Schüring, A.; Kulle, A.E.; Holterhus, P.M.; von Wolff, M.; et al. The Action of Reproductive Fluids and Contained Steroids, Prostaglandins, and Zn(2+) on CatSper Ca(2+) Channels in Human Sperm. Front. Cell Dev. Biol. 2021, 9, 699554.
- Brenker, C.; Goodwin, N.; Weyand, I.; Kashikar, N.D.; Naruse, M.; Krähling, M.; Müller, A.; Kaupp, U.B.; Strünker, T. The CatSper channel: A polymodal chemosensor in human sperm. EMBO J. 2012, 31, 1654–1665.
- Huang, X.; Miyata, H.; Wang, H.; Mori, G.; Iida-Norita, R.; Ikawa, M.; Percudani, R.; Chung, J.J. A CUG-initiated CATSPERθ functions in the CatSper channel assembly and serves as a checkpoint for flagellar trafficking. bioRxiv 2023.
- Yang, F.; Gracia Gervasi, M.; Orta, G.; Tourzani, D.A.; De la Vega-Beltran, J.L.; Ruthel, G.; Darszon, A.; Visconti, P.E.; Wang, P.J. C2CD6 regulates targeting and organization of the CatSper calcium channel complex in sperm flagella. Development 2022, 149, dev199988.
- Lin, S.; Ke, M.; Zhang, Y.; Yan, Z.; Wu, J. Structure of a mammalian sperm cation channel complex. Nature 2021, 595, 746–750.
- Zhao, Y.; Wang, H.; Wiesehoefer, C.; Shah, N.B.; Reetz, E.; Hwang, J.Y.; Huang, X.; Wang, T.E.; Lishko, P.V.; Davies, K.M.; et al. 3D structure and in situ arrangements of CatSper channel in the sperm flagellum. Nat. Commun. 2022, 13, 3439.
- Avenarius, M.R.; Hildebrand, M.S.; Zhang, Y.; Meyer, N.C.; Smith, L.L.; Kahrizi, K.; Najmabadi, H.; Smith, R.J. Human male infertility caused by mutations in the CATSPER1 channel protein. Am. J. Hum. Genet. 2009, 84, 505–510.
- Luo, T.; Chen, H.Y.; Zou, Q.X.; Wang, T.; Cheng, Y.M.; Wang, H.F.; Wang, F.; Jin, Z.L.; Chen, Y.; Weng, S.Q.; et al. A novel copy number variation in CATSPER2 causes idiopathic male infertility with normal semen parameters. Hum. Reprod. 2019, 34, 414–423.
- Wang, J.; Tang, H.; Zou, Q.; Zheng, A.; Li, H.; Yang, S.; Xiang, J. Patient with CATSPER3 mutations-related failure of sperm acrosome reaction with successful pregnancy outcome from intracytoplasmic sperm injection (ICSI). Mol. Genet. Genom. Med. 2021, 9, e1579.
- Chung, J.J.; Navarro, B.; Krapivinsky, G.; Krapivinsky, L.; Clapham, D.E. A novel gene required for male fertility and functional CATSPER channel formation in spermatozoa. Nat. Commun. 2011, 2, 153.
- Carlson, A.E.; Quill, T.A.; Westenbroek, R.E.; Schuh, S.M.; Hille, B.; Babcock, D.F. Identical phenotypes of CatSper1 and CatSper2 null sperm. J. Biol. Chem. 2005, 280, 32238–32244.
- Jin, J.; Jin, N.; Zheng, H.; Ro, S.; Tafolla, D.; Sanders, K.M.; Yan, W. Catsper3 and Catsper4 are essential for sperm hyperactivated motility and male fertility in the mouse. Biol. Reprod. 2007, 77, 37–44.
- Qi, H.; Moran, M.M.; Navarro, B.; Chong, J.A.; Krapivinsky, G.; Krapivinsky, L.; Kirichok, Y.; Ramsey, I.S.; Quill, T.A.; Clapham, D.E. All four CatSper ion channel protein are required for male fertility and sperm cell hyperactivated motility. Proc. Natl. Acad. Sci. USA 2007, 104, 1219–1223.
- Avidan, N.; Tamary, H.; Dgany, O.; Cattan, D.; Pariente, A.; Thulliez, M.; Borot, N.; Moati, L.; Barthelme, A.; Shalmon, L.; et al. CATSPER2, a human autosomal nonsyndromic male infertility gene. Eur. J. Hum. Genet. 2003, 11, 497–502.
- Williams, H.L.; Mansell, S.; Alasmari, W.; Brown, S.G.; Wilson, S.M.; Sutton, K.A.; Miller, M.R.; Lishko, P.V.; Barratt, C.L.R.; Publicover, S.J.; et al. Specific loss of CatSper function is sufficient to compromise fertilizing capacity of human spermatozoa. Hum. Reprod. 2015, 30, 28–38.
- Brown, S.G.; Miller, M.R.; Lishko, P.V.; Lester, D.H.; Publicover, S.J.; Barratt, C.L.R.; Da Silva, S.M. Homozygous in-frame deletion in CATSPERE in a man producing spermatozoa with loss of CatSper function and compromised fertilizing capacity. Hum. Reprod. 2018, 33, 1812–1816.
- Wang, H.; McGoldrick, L.L.; Chung, J.-J. Sperm ion channels and transporters in male fertility and infertility. Nat. Rev. Urol. 2021, 18, 46–66.
- Navarro, B.; Kirichok, Y.; Clapham, D.E. KSper, a pH-sensitive K+ current that controls sperm membrane potential. Proc. Natl. Acad. Sci. USA 2007, 104, 7688–7692.
- Chavez, J.C.; Ferreira, J.J.; Butler, A.; De La Vega Beltran, J.L.; Trevino, C.L.; Darszon, A.; Salkoff, L.; Santi, C.M. SLO3K+ channels control calcium entry through CATSPER channels in sperm. J. Biol. Chem. 2014, 289, 32266–32275.
- Hwang, J.Y.; Chung, J.J. CatSper Calcium Channels: 20 Years On. Physiology 2023, 38, 125–140.
- Brown, S.G.; Costello, S.; Kelly, M.C.; Ramalingam, M.; Drew, E.; Publicover, S.J.; Barratt, C.L.R.; Da Silva, S.M. Complex CatSper-dependent and independent i signalling in human spermatozoa induced by follicular fluid. Hum. Reprod. 2017, 32, 1995–2006.
- Sumigama, S.; Mansell, S.; Miller, M.; Lishko, P.V.; Cherr, G.N.; Meyers, S.A.; Tollner, T. Progesterone Accelerates the Completion of Sperm Capacitation and Activates CatSper Channel in Spermatozoa from the Rhesus Macaque. Biol. Reprod. 2015, 93, 130.
- Lishko, P.V.; Botchkina, I.L.; Kirichok, Y. Progesterone activates the principal Ca2+ channel of human sperm. Nature 2011, 471, 387–391.
- Miller, M.R.; Mannowetz, N.; Iavarone, A.T.; Safavi, R.; Gracheva, E.O.; Smith, J.F.; Hill, R.Z.; Bautista, D.M.; Kirichok, Y.; Lishko, P.V. Unconventional endocannabinoid signaling governs sperm activation via the sex hormone progesterone. Science 2016, 352, 555–559.
- Uhler, M.L.; Leungt, A.; Chan, S.Y.W.; Wang, C. Direct effects of progesterone and antiprogesterone on human sperm hyperactivated motility and acrosome reaction. Fertil. Steril. 1992, 58, 1191–1198.
- Mannowetz, N.; Miller, M.R.; Lishko, P.V. Regulation of the sperm calcium channel CatSper by endogenous steroids and plant triterpenoids. Proc. Natl. Acad. Sci. USA 2017, 114, 5743–5748.
- Brenker, C.; Schiffer, C.; Wagner, I.V.; Tüttelmann, F.; Röpke, A.; Rennhack, A.; Kaupp, U.B.; Strünker, T. Action of steroids and plant triterpenoids on CatSper Ca(2+) channels in human sperm. Proc. Natl. Acad. Sci. USA 2018, 115, E344–E346.
- Rehfeld, A. Revisiting the action of steroids and triterpenoids on the human sperm Ca2+ channel CatSper. Mol. Hum. Reprod. 2020, 26, 816–824.
- Sánchez González, S.R.; Mata Martínez, E.; Torres Juárez, J.A.; Arias, R.J.; De Blas, G.A.; Sánchez Tusie, A.A. Cortisol modulates Ca(2+) signaling and acrosome reaction in human sperm. Andrology 2023, 11, 134–142.
- Carlson, E.J.; Georg, G.I.; Hawkinson, J.E. Steroidal Antagonists of Progesterone- and Prostaglandin E(1)-Induced Activation of the Cation Channel of Sperm. Mol. Pharmacol. 2022, 101, 56–67.
- Shimizu, Y.; Yorimitsu, A.; Maruyama, Y.; Kubota, T.; Aso, T.; ABronson, R. Prostaglandins induce calcium influx in human spermatozoa. Mol. Hum. Reprod. 1998, 4, 555–561.
- Strünker, T.; Goodwin, N.; Brenker, C.; Kashikar, N.D.; Weyand, I.; Seifert, R.; Kaupp, U.B. The CatSper channel mediates progesterone-induced Ca2+ influx in human sperm. Nature 2011, 471, 382–386.
- Xia, J.; Reigada, D.; Mitchell, C.H.; Ren, D. CATSPER channel-mediated Ca2+ entry into mouse sperm triggers a tail-to-head propagation. Biol. Reprod. 2007, 77, 551–559.
- Kobori, H.; Miyazaki, S.; Kuwabara, Y. Characterization of intracellular Ca(2+) increase in response to progesterone and cyclic nucleotides in mouse spermatozoa. Biol. Reprod. 2000, 63, 113–120.
- Hereng, T.H.; Elgstoen, K.B.; Eide, L.; Rosendal, K.R.; Skalhegg, B.S. Serum albumin and HCO3− regulate separate pools of ATP in human spermatozoa. Hum. Reprod. 2014, 29, 918–930.
- Boisen, I.M.; Rehfeld, A.; Mos, I.; Poulsen, N.N.; Nielsen, J.E.; Schwarz, P.; Rejnmark, L.; Dissing, S.; Bach-Mortensen, P.; Juul, A.; et al. The Calcium-Sensing Receptor Is Essential for Calcium and Bicarbonate Sensitivity in Human Spermatozoa. J. Clin. Endocrinol. Metab. 2021, 106, e1775–e1792.
- Ferreira, J.J.; Lybaert, P.; Puga-Molina, L.C.; Santi, C.M. Conserved Mechanism of Bicarbonate-Induced Sensitization of CatSper Channels in Human and Mouse Sperm. Front. Cell Dev. Biol. 2021, 9, 733653.
- Xia, J.; Ren, D. Egg coat proteins activate calcium entry into mouse sperm via CATSPER channels. Biol. Reprod. 2009, 80, 1092–1098.
- Balbach, M.; Hamzeh, H.; Jikeli, J.F.; Brenker, C.; Schiffer, C.; Hansen, J.N.; Neugebauer, P.; Trötschel, C.; Jovine, L.; Han, L.; et al. Molecular Mechanism Underlying the Action of Zona-pellucida Glycoproteins on Mouse Sperm. Front. Cell Dev. Biol. 2020, 8, 572735.
- Ren, D.; Navarro, B.; Perez, G.; Jackson, A.C.; Hsu, S.; Shi, Q.; Tilly, J.L.; Clapham, D.E. A sperm ion channel required for sperm motility and male fertility. Nature 2001, 413, 603–609.
- Xia, J.; Ren, D. The BSA-induced Ca2+ influx during sperm capacitation is CATSPER channel-dependent. Reprod. Biol. Endocrinol. 2009, 7, 119.
- Pazgier, M.; Hoover, D.M.; Yang, D.; Lu, W.; Lubkowski, J. Human beta-defensins. Cell. Mol. Life Sci. 2006, 63, 1294–1313.
- Diao, R.; Fok, K.L.; Chen, H.; Mei Kuen, Y.; Duan, Y.; Chung, C.M.; Li, Z.; Wu, H.; Li, Z.; Zhang, H.; et al. Deficient human b-defensin 1 underlies male infertility associated with poor sperm motility and genital tract infection. Sci. Transl. Med. 2014, 6, 249ra108.
- Diao, R.; Wang, T.; Fok, K.L.; Li, X.; Ruan, Y.; Yu, M.K.; Cheng, Y.; Chen, Y.; Chen, H.; Mou, L.; et al. CCR6 is required for ligand-induced CatSper activation in human sperm. Oncotarget 2017, 8, 91445–91458.
- Li, X.; Yuan, C.; Shi, J.; Kang, H.; Chen, Y.; Duan, Y.; Jin, J.; Cheung, L.P.; Li, T.C.; Liu, Y.; et al. β-Defensin 19/119 mediates sperm chemotaxis and is associated with idiopathic infertility. Cell Rep. Med. 2022, 3, 100825.
- Meizel, S. The sperm, a neuron with a tail: ‘Neuronal’ receptors in mammalian sperm. Biol. Rev. Camb. Philos. Soc. 2004, 79, 713–732.
- Ramirez-Reveco, A.; Villarroel-Espindola, F.; Rodriguez-Gil, J.E.; Concha, I.I. Neuronal signaling repertoire in the mammalian sperm functionality. Biol. Reprod. 2017, 96, 505–524.
- Sakamoto, C.; Fujinoki, M.; Kitazawa, M.; Obayashi, S. Serotonergic signals enhanced hamster sperm hyperactivation. J. Reprod. Dev. 2021, 67, 241–250.
- Yoshida, M.; Yoshida, K. Sperm chemotaxis and regulation of flagellar movement by Ca2+. Mol. Hum. Reprod. 2011, 17, 457–465.
- Cheng, Y.-M.; Luo, T.; Peng, Z.; Chen, H.-Y.; Zhang, J.; Zeng, X.-H. OR1D2 receptor mediates bourgeonal-induced human CatSper activation in a G-protein dependent manner. bioRxiv 2019, 757880.
- Sinding, C.; Kemper, E.; Spornraft-Ragaller, P.; Hummel, T. Decreased Perception of Bourgeonal May Be Linked to Male Idiopathic Infertility. Chem. Senses 2013, 38, 439–445.
- Wang, Y.N.; Wang, B.; Liang, M.; Han, C.Y.; Zhang, B.; Cai, J.; Sun, W.; Xing, G.G. Down-regulation of CatSper1 channel in epididymal spermatozoa contributes to the pathogenesis of asthenozoospermia, whereas up-regulation of the channel by Sheng-Jing-San treatment improves the sperm motility of asthenozoospermia in rats. Fertil. Steril. 2013, 99, 579–587.
- Sadogh, A.; Gorji, N.; Moeini, R. Herbal foodstuffs in Avicenna’s recommended diet to improve sperm quality and increase male fertility; an evidence-based approach. J. Complement. Integr. Med. 2021, 19, 47–70.
- Kim, D.R.; Kim, H.Y.; Kim, H.Y.; Chang, M.S.; Park, S.K. Trigonellae Semen Enhances Sperm Motility and the Expression of the Cation Sperm Channel Proteins in Mouse Testes. Evid.-Based Complement. Altern. Med. 2015, 2015, 817324.
- Mancuso, C.; Santangelo, R. Panax ginseng and Panax quinquefolius: From pharmacology to toxicology. Food Chem. Toxicol. 2017, 107, 362–372.
- Park, E.H.; Kim, D.R.; Kim, H.Y.; Park, S.K.; Chang, M.S. Panax ginseng induces the expression of CatSper genes and sperm hyperactivation. Asian J. Androl. 2014, 16, 845–851.
- Italiya, J.M.; Patel, M.R.; Golaviya, A.V.; Patel, S.S.; Thakkar, B.K.; Jakhesara, S.J.; Joshi, C.G.; Koringa, P.G. RNA-sequencing attest increased sperm motility in bovine spermatozoa treated with ethanolic extract of Putranjiva roxburghii. 3 Biotech 2023, 13, 33.
- Kiani, K.; Rudzitis-Auth, J.; Scheuer, C.; Movahedin, M.; Sadati Lamardi, S.N.; Malekafzali Ardakani, H.; Becker, V.; Moini, A.; Aflatoonian, R.; Ostad, S.N.; et al. Calligonum comosum (Escanbil) extract exerts anti-angiogenic, anti-proliferative and anti-inflammatory effects on endometriotic lesions. J. Ethnopharmacol. 2019, 239, 111918.
- Askari Jahromi, M.; Movahedin, M.; Mazaheri, Z.; Amanlu, M.; Mowla, S.J.; Batooli, H. Evaluating the effects of Escanbil (Calligonum) extract on the expression level of Catsper gene variants and sperm motility in aging male mice. Iran. J. Reprod. Med. 2014, 12, 459–466.
- Kumar, V.S.; Sharma, V.L.; Tiwari, P.; Singh, D.; Maikhuri, J.P.; Gupta, G.; Singh, M.M. The spermicidal and antitrichomonas activities of SSRI antidepressants. Bioorg. Med. Chem. Lett. 2006, 16, 2509–2512.
- Rahban, R.; Rehfeld, A.; Schiffer, C.; Brenker, C.; Egeberg Palme, D.L.; Wang, T.; Lorenz, J.; Almstrup, K.; Skakkebaek, N.E.; Strünker, T.; et al. The antidepressant Sertraline inhibits CatSper Ca2+ channels in human sperm. Hum. Reprod. 2021, 36, 2638–2648.
- Amory, J.K.; Wang, C.; Swerdloff, R.S.; Anawalt, B.D.; Matsumoto, A.M.; Bremner, W.J.; Walker, S.E.; Haberer, L.J.; Clark, R.V. The effect of 5alpha-reductase inhibition with dutasteride and finasteride on semen parameters and serum hormones in healthy men. J. Clin. Endocrinol. Metab. 2007, 92, 1659–1665.
- Birch, M.R.; Dissing, S.; Skakkebæk, N.E.; Rehfeld, A. Finasteride interferes with prostaglandin-induced CatSper signalling in human sperm. Reproduction 2021, 161, 561–572.
- Srivastav, A.; Changkija, B.; Sharan, K.; Nagar, G.K.; Bansode, F.W. Influence of antifertility agents Dutasteride and Nifedipine on CatSper gene level in epididymis during sperm maturation in BALB/c mice. Reproduction 2018, 155, 347–359.
- Smarr, M.M.; Kannan, K.; Chen, Z.; Kim, S.; Buck Louis, G.M. Male urinary paracetamol and semen quality. Andrology 2017, 5, 1082–1088.
- Rehfeld, A.; Frederiksen, H.; Rasmussen, R.H.; David, A.; Chaker, J.; Nielsen, B.S.; Nielsen, J.E.; Juul, A.; Skakkebæk, N.E.; Kristensen, D.M. Human sperm cells can form paracetamol metabolite AM404 that directly interferes with sperm calcium signalling and function through a CatSper-dependent mechanism. Hum. Reprod. 2022, 37, 922–935.
- Morakinyo, A.O.; Iranloye, B.O.; Adegoke, O.A. Antireproductive effect of calcium channel blockers on male rats. Reprod. Med. Biol. 2009, 8, 97–102.
- Rennhack, A.; Schiffer, C.; Brenker, C.; Fridman, D.; Nitao, E.T.; Cheng, Y.M.; Tamburrino, L.; Balbach, M.; Stolting, G.; Berger, T.K.; et al. A novel cross-species inhibitor to study the function of CatSper Ca(2+) channels in sperm. Br. J. Pharmacol. 2018, 175, 3144–3161.
- McBrinn, R.C.; Fraser, J.; Hope, A.G.; Gray, D.W.; Barratt, C.L.R.; da Silva, S.J.M.; Brown, S.G. Novel pharmacological actions of trequinsin hydrochloride improve human sperm cell motility and function. Br. J. Pharmacol. 2019, 176, 4521–4536.
- Howell, S.J.; Shalet, S.M. Spermatogenesis after cancer treatment: Damage and recovery. J. Natl. Cancer Inst. Monogr. 2005, 2005, 12–17.
- Razak, S.; Afsar, T.; Bibi, N.; Abulmeaty, M.; Qamar, W.; Almajwal, A.; Inam, A.; AI Disi, D.; Shabbir, M.; Bhat, M.A. Molecular docking, pharmacokinetic studies, and in vivo pharmacological study of indole derivative 2-(5-methoxy-2-methyl-1H-indole-3-yl)-N’- acetohydrazide as a promising chemoprotective agent against cisplatin induced organ damage. Sci. Rep. 2021, 11, 6245.
- Afsar, T.; Razak, S.; Trembley, J.H.; Khan, K.; Shabbir, M.; Almajwal, A.; Alruwaili, N.W.; Ijaz, M.U. Prevention of Testicular Damage by Indole Derivative MMINA via Upregulated StAR and CatSper Channels with Coincident Suppression of Oxidative Stress and Inflammation: In Silico and In Vivo Validation. Antioxidants 2022, 11, 2063.
- Yilmaz, B.; Terekeci, H.; Sandal, S.; Kelestimur, F. Endocrine disrupting chemicals: Exposure, effects on human health, mechanism of action, models for testing and strategies for prevention. Rev. Endocr. Metab. Disord. 2020, 21, 127–147.
- Shannon, M.; Rehfeld, A.; Frizzell, C.; Livingstone, C.; McGonagle, C.; Skakkebaek, N.E.; Wielogórska, E.; Connolly, L. In vitro bioassay investigations of the endocrine disrupting potential of steviol glycosides and their metabolite steviol, components of the natural sweetener Stevia. Mol. Cell. Endocrinol. 2016, 427, 65–72.
- Zhou, Y.; Xu, W.; Yuan, Y.; Luo, T. What is the Impact of Bisphenol A on Sperm Function and Related Signaling Pathways: A Mini-review? Curr. Pharm. Des. 2020, 26, 4822–4828.
- Le Fol, V.; Ait-Aissa, S.; Sonavane, M.; Porcher, J.M.; Balaguer, P.; Cravedi, J.P.; Zalko, D.; Brion, F. In vitro and in vivo estrogenic activity of BPA, BPF and BPS in zebrafish-specific assays. Ecotoxicol. Environ. Saf. 2017, 142, 150–156.
- Wang, H.F.; Liu, M.; Li, N.; Luo, T.; Zheng, L.P.; Zeng, X.H. Bisphenol A Impairs Mature Sperm Functions by a CatSper-Relevant Mechanism. Toxicol. Sci. 2016, 152, 145–154.
- Yuan, W.B.; Chen, H.Q.; Li, J.Z.; Zhou, S.M.; Zeng, Y.; Fan, J.; Zhang, Z.; Liu, J.Y.; Cao, J.; Liu, W.B. TET1 mediated male reproductive toxicity induced by Bisphenol A through Catsper-Ca(2+) signaling pathway. Environ. Pollut. 2022, 296, 118739.
- Rehfeld, A.; Andersson, A.M.; Skakkebaek, N.E. Bisphenol A Diglycidyl Ether (BADGE) and Bisphenol Analogs, but Not Bisphenol A (BPA), Activate the CatSper Ca(2+) Channel in Human Sperm. Front. Endocrinol. 2020, 11, 324.
- Zou, Q.X.; Peng, Z.; Zhao, Q.; Chen, H.Y.; Cheng, Y.M.; Liu, Q.; He, Y.Q.; Weng, S.Q.; Wang, H.F.; Wang, T.; et al. Diethylstilbestrol activates CatSper and disturbs progesterone actions in human spermatozoa. Hum. Reprod. 2017, 32, 290–298.
- Yuan, Y.; Ding, X.; Cheng, Y.; Kang, H.; Luo, T.; Zhang, X.; Kuang, H.; Chen, Y.; Zeng, X.; Zhang, D. PFOA evokes extracellular Ca(2+) influx and compromises progesterone-induced response in human sperm. Chemosphere 2020, 241, 125074.
- Diamanti-Kandarakis, E.; Bourguignon, J.P.; Giudice, L.C.; Hauser, R.; Prins, G.S.; Soto, A.M.; Zoeller, R.T.; Gore, A.C. Endocrine-disrupting chemicals: An Endocrine Society scientific statement. Endocr. Rev. 2009, 30, 293–342.
- Wang, H.F.; Chang, M.; Peng, T.T.; Yang, Y.; Li, N.; Luo, T.; Zhou, M.Z.; Zeng, X.H.; Zheng, L.P. Exposure to Cadmium Impairs Sperm Functions by Reducing CatSper in Mice. Cell. Physiol. Biochem. 2017, 42, 44–54.
- Tavares, R.S.; Mansell, S.; Barratt, C.L.; Wilson, S.M.; Publicover, S.J.; Ramalho-Santos, J. p,p′-DDE activates CatSper and compromises human sperm function at environmentally relevant concentrations. Hum. Reprod. 2013, 28, 3167–3177.
- Zhang, X.; Kang, H.; Peng, L.; Song, D.; Jiang, X.; Li, Y.; Chen, H.; Zeng, X. Pentachlorophenol inhibits CatSper function to compromise progesterone’s action on human sperm. Chemosphere 2020, 259, 127493.
- Birch, M.R.; Johansen, M.; Skakkebaek, N.E.; Andersson, A.M.; Rehfeld, A. In vitro investigation of endocrine disrupting effects of pesticides on Ca(2+)-signaling in human sperm cells through actions on the sperm-specific and steroid-activated CatSper Ca(2+)-channel. Environ. Int. 2022, 167, 107399.
- Schiffer, C.; Müller, A.; Egeberg, D.L.; Alvarez, L.; Brenker, C.; Rehfeld, A.; Frederiksen, H.; Wäschle, B.; Kaupp, U.B.; Balbach, M.; et al. Direct action of endocrine disrupting chemicals on human sperm. EMBO Rep. 2014, 15, 758–765.
- Rehfeld, A.; Dissing, S.; Skakkebaek, N.E. Chemical UV Filters Mimic the Effect of Progesterone on Ca(2+) Signaling in Human Sperm Cells. Endocrinology 2016, 157, 4297–4308.
- Alavi, S.H.; Taghavi, M.M.; Moallem, S.A. Evaluation of effects of methamphetamine repeated dosing on proliferation and apoptosis of rat germ cells. Syst. Biol. Reprod. Med. 2008, 54, 85–91.
- Allaeian Jahromi, Z.; Meshkibaf, M.H.; Naghdi, M.; Vahdati, A.; Makoolati, Z. Methamphetamine Downregulates the Sperm-Specific Calcium Channels Involved in Sperm Motility in Rats. ACS Omega 2022, 7, 5190–5196.
- Wang, H.; Liu, J.; Cho, K.H.; Ren, D. A novel, single, transmembrane protein CATSPERG is associated with CATSPER1 channel protein. Biol. Reprod. 2009, 81, 539–544.
- He, Y.; Zou, Q.; Li, B.; Chen, H.; Du, X.; Weng, S.; Luo, T.; Zeng, X. Ketamine inhibits human sperm function by Ca(2+)-related mechanism. Biochem. Biophys. Res. Commun. 2016, 478, 501–506.
- Chen, Y.; Xu, W.; Yuan, Y.; Chen, H.; Zheng, S.; He, Y.; Luo, T. N-Methyl-d-aspartic Acid (NMDA) Receptor Is Involved in the Inhibitory Effect of Ketamine on Human Sperm Functions. Int. J. Mol. Sci. 2021, 22, 12370.
- Wright, C.; Milne, S.; Leeson, H. Sperm DNA damage caused by oxidative stress: Modifiable clinical, lifestyle and nutritional factors in male infertility. Reprod. Biomed. Online 2014, 28, 684–703.
- Badouard, C.; Ménézo, Y.; Panteix, G.; Ravanat, J.L.; Douki, T.; Cadet, J.; Favier, A. Determination of new types of DNA lesions in human sperm. Zygote 2008, 16, 9–13.
- Gonzalez-Marin, C.; Gosalvez, J.; Roy, R. Types, causes, detection and repair of DNA fragmentation in animal and human sperm cells. Int. J. Mol. Sci. 2012, 13, 14026–14052.
- Tremellen, K. Oxidative stress and male infertility—A clinical perspective. Hum. Reprod. Update 2008, 14, 243–258.
- Mohammadi, S.; Jalali, M.; Nikravesh, M.R.; Fazel, A.; Ebrahimzadeh, A.; Gholamin, M.; Sankian, M. Effects of Vitamin-E treatment on CatSper genes expression and sperm quality in the testis of the aging mouse. Iran. J. Reprod. Med. 2013, 11, 989–998.
- Mohammadi, S.; Movahedin, M.; Mowla, S.J. Up-regulation of CatSper genes family by selenium. Reprod. Biol. Endocrinol. 2009, 7, 126.

