Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Muhammad Asad Farooq | -- | 2039 | 2023-08-31 09:14:46 | | | |
| 2 | Peter Tang | Meta information modification | 2039 | 2023-08-31 10:29:08 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Farooq, M.A.; Ajmal, I.; Hui, X.; Chen, Y.; Ren, Y.; Jiang, W. Eminent Role of β2-AR in T Cells. Encyclopedia. Available online: https://encyclopedia.pub/entry/48674 (accessed on 26 June 2026).
Farooq MA, Ajmal I, Hui X, Chen Y, Ren Y, Jiang W. Eminent Role of β2-AR in T Cells. Encyclopedia. Available at: https://encyclopedia.pub/entry/48674. Accessed June 26, 2026.
Farooq, Muhammad Asad, Iqra Ajmal, Xinhui Hui, Yiran Chen, Yaojun Ren, Wenzheng Jiang. "Eminent Role of β2-AR in T Cells" Encyclopedia, https://encyclopedia.pub/entry/48674 (accessed June 26, 2026).
Farooq, M.A., Ajmal, I., Hui, X., Chen, Y., Ren, Y., & Jiang, W. (2023, August 31). Eminent Role of β2-AR in T Cells. In Encyclopedia. https://encyclopedia.pub/entry/48674
Farooq, Muhammad Asad, et al. "Eminent Role of β2-AR in T Cells." Encyclopedia. Web. 31 August, 2023.
Copy Citation
Beta2-adrenergic receptors (β2-ARs) are an emerging class of receptors that are capable of modulating the functioning of immune cells. β2-AR is reported to activate regulatory immune cells and inhibit effector immune cells. Blocking β2-AR increases activation, proliferation, and cytokine release of T lymphocytes. Moreover, β2-AR deficiency during metabolic reprogramming of T cells increases mitochondrial membrane potential and biogenesis.
β2-adrenergic receptor
CAR-T therapy
1. Introduction
Neurotransmitters play a bridging role between the nervous system and the body. Catecholamines are important set of neurotransmitters released by supra-renal glands and comprise adrenaline/epinephrine (A/E) and nor-adrenaline/nor-epinephrine (NA/NE). Adrenergic receptors are activated in response to stimulation by adrenaline and nor-adrenaline to chemically coordinate the signals/messages from nervous system to the target tissues [1][2]. Adrenergic receptors (ARs) belong to the G-protein coupled, seven transmembrane receptor (GPCR) family and constitute alpha-adrenergic receptor (α-AR) and beta-adrenergic receptor (β-AR) subtypes, which further have been classified as Alpha-1 adrenergic receptors (α1-ARs), Alpha-2 adrenergic receptors (α2-ARs), Beta-1 adrenergic receptors (β1-ARs), Beta-2 adrenergic receptors (β2-ARs) and Beta-3 adrenergic receptors (β3-ARs). In short-term or acute stress conditions like fear or exercise, the body releases catecholamines via activation of the sympathetic nervous system (SNS) that bind with adrenergic receptors on various organs, resulting in increased heart rate, dilation of pupils, mobilization of energy, and diversion of the blood flow from other body organs to the skeletal muscles to cope with the aforementioned stress [3].
However, in chronic stress conditions, the body retains a persistently higher concentration of catecholamines and sustained activation of SNS, leading to the initiation and progression of cancer [4]. Many studies have demonstrated that this cancer progression is due to catecholamines triggering β-adrenergic receptors, more specifically, downstream signaling of β2-Adrenergic receptors in cancer cells [5]. Later on, it was demonstrated that psychological factors and depressive disorders can lead to cancer incidence. During stress, elevated levels of catecholamines can lead to cancer initiation by making the genome unstable, rendering them exposed to environmental carcinogens [6]. Because of this, more studies were carried out aiming to unearth the role of β2-AR in tumor initiation and progression. Recently, the role of β2-AR in immunity has been the focus of many studies. β2-Adrenergic receptors can modulate the functions of various immune cells as the corresponding receptors are displayed on the surface of T cells, natural killer (NK) cells, and dendritic cells (DCs) [7][8]. The ligand for β2-AR is released inside the tumor microenvironment (TME) and leads to inhibitory signaling in T lymphocytes [9][10].
The immune system plays a critical role in identifying and eliminating cells that undergo malignant transformation, in addition to its primary function of eradicating infectious agents. If the inflammatory response remains unresolved, the affected cells undergo transformation and become cancerous [11]. However, at this stage, the immune system identifies the tumor cells by recognizing the tumor-specific antigens displayed on their surfaces [12], followed by effector immune responses primarily mediated by CD8+ T cells and NK cells. The activation of these immune responses forms the basis for immunotherapy, as it halts tumorigenesis [13][14].
Due to the remarkable success of the initial clinical trials of chimeric antigen receptor (CAR)-T therapy, particularly in pediatric patients with hematological malignancies, the clinical response rates in leukemia patients have reached as high as 90% [15][16]. As a result, the number of successful clinical trials for CAR-T therapy targeting hematological malignancies which direct numerous antigens has upsurged significantly, yet the parallel success of CAR-T therapy in solid tumors is still being awaited [17][18]. The number of ongoing clinical trials in solid tumors is far less than in liquid tumors, courtesy of the toxic side effects and suboptimal therapeutic outcomes which are achieved almost every time. There can be several explanations for this, including the following. Hematological cancers commonly exhibit similar antigens, and their distribution across various types of hematological cancers is generally similar, with some exceptions [19]. On the contrary, antigens related to solid tumors vary greatly, not just from tumor to tumor, but also between different forms of a similar tumor, i.e., primary and metastatic forms [20]. Moreover, hematological cancers are widespread in the circulatory system, which makes them less dense and easily targetable as compared to solid tumors. The solid tumors are denser and are concentrated on a single site, creating a physical barrier for CAR-T cells by developing extensive vasculature to supply ample nutrients to the fast-growing tumor, thus inhibiting the chemo-attractive signals that are necessary for CAR-T cells to reach the tumor site [21]. The immune-suppressive characteristics of solid tumors, which arise due to the presentation of checkpoint inhibitory ligands and metabolites from diverse metabolic pathways collectively create a tumor microenvironment (TME). This microenvironment makes it exceedingly challenging for CAR-T cells to infiltrate and effectively eliminate solid tumors, leading to significant barriers to CAR-T cell therapy’s success [22]. Adrenergic stress is one of the culprits, among others, responsible for immunosuppression inside the TME. Previously, the researchers have attempted to target immunosuppressive factors inside the TME in an attempt to increase the efficacy of CAR-T therapy in the prostate and pancreatic tumor microenvironments [23][24].
2. Nor-Adrenaline in Tumor Microenvironment
The production of adrenaline and nor-adrenaline within the TME can arise from various sources, including the tumor cells themselves, the nerve fibers that innervate the tumors, and the surrounding stromal cells such as fibroblasts and immune cells. It seems that NA, mainly from the innervating nerves, plays a role in the incidence and initial progression of the cancer, as TME is absent initially. After TME is established, it is mainly the TME that pours sufficient amounts of NA into the tumor surroundings that carry its immunosuppressive roles in CD8+ T cells. This NA comes mainly from two sources. Firstly, the tumor itself contains all the necessary machinery to synthesize NA. Secondly, many tumor types are innervated by sympathetic nerve fibers [25]. Neuronal progenitor cells migrate towards the tumor tissue during the process of neurogenesis and adapt a sympathetic tone while innervating tumor tissue. Moreover, the sensory neurons innervating tumor tissues are also reprogrammed to sympathetic nerve phenotypes [26]. Altogether, this creates hyperactive SNS signaling inside the TME. Growing evidence suggest that an immunosuppressive role of the adrenergic system inside the TME could possibly hinder the antitumor functions of immunotherapy [6][27].
3. Adrenergic Stress Endorses Immunosuppressive Tumor Milieu Development
The catecholamines bind to adrenergic receptors on target cells, including immune cells, and trigger a range of biological responses. In the context of the immune system, adrenergic stress can change the activities of different immune cell subsets due to its wide expression. One of the key mechanisms by which adrenergic stress promotes immunosuppression is through the recruitment and activation of myeloid-derived suppressor cells (MDSCs) [28]. MDSCs are regulatory immune cells that have the skill to inhibit T cells and promote tumor growth. Adrenergic stress has been shown to promote the expansion and activation of MDSCs in the tumor microenvironment through the activation of adrenergic receptors on MDSCs themselves, as well as on other cells that produce cytokines and chemokines which promote the recruitment and activation of MDSCs [29]. β2-AR activation on MDSCs has been shown to enhance the expression of arginase-1, an enzyme that can deplete arginine, an amino acid that is important for T cell function. Additionally, adrenergic stress inhibits T cell functions by upregulating interleukin-10 (IL-10) and transforming growth factor-beta (TGF-β) [30][31]. The production of these anti-inflammatory cytokines, owing to the activation of β2-AR on the surfaces of tumor-associated macrophages (TAMS), also demonstrated as M2-phenotype. Adrenergic signaling in cancer plays a significant role in fostering the release of vascular endothelial growth factor (VEGF), a crucial mediator of angiogenesis, from both tumor cells and M2-macrophages residing in the tumor microenvironment [32]. VEGF facilitates the recruitment of endothelial cells, leading to the formation of new blood vessels, which in turn support the delivery of oxygen and nutrients to the tumor [27]. Additionally, the activation of β2-adrenergic receptors (β2-AR) can enhance the activity of regulatory T cells (Tregs), thereby amplifying the immunosuppressive environment around the tumor [33][34].
According to the literature, adrenergic stress has been shown to inhibit T cell activation and proliferation, and to promote apoptosis. Similarly, it can inhibit NK cell and B cell activation.
The observed effects are facilitated through the stimulation of adrenergic receptors, leading to the secretion of cytokines and chemokines that contribute to the impairment of immune cell function [35]. In conclusion, adrenergic stress plays a key role in the development of the TME by promoting the recruitment and activation of immune-regulatory cells, inhibiting the function of pro-inflammatory immune cells (Figure 1), endorsing the production of immunosuppressive cytokines and chemokines, and promoting tumor angiogenesis [36]. Here, the researchers discuss the role of adrenergic stress, specifically in T lymphocytes.
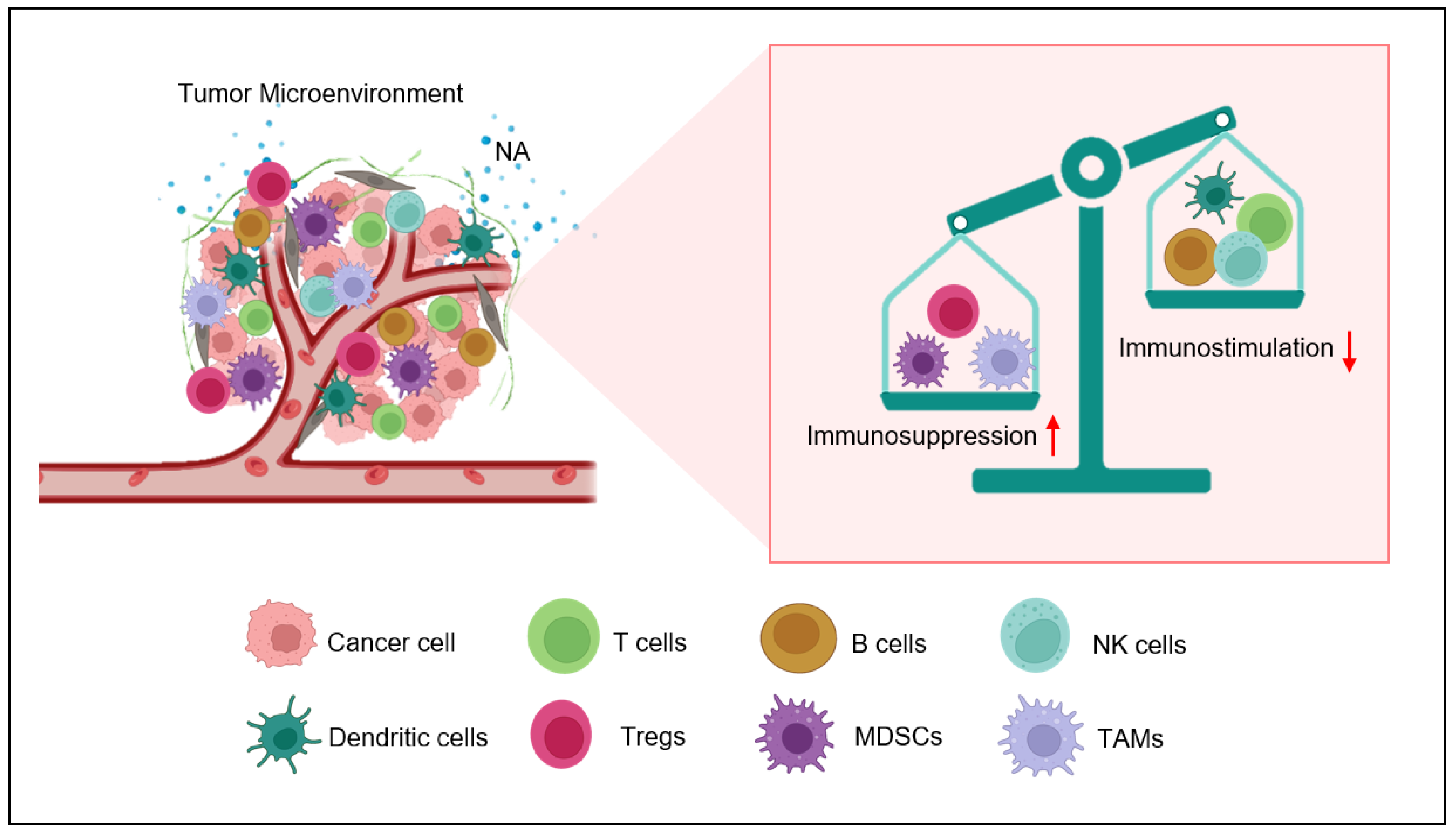
Figure 1. Nor-adrenaline (NA)-mediated immunosuppression inside the tumor microenvironment. Nor-adrenaline released in the tumor microenvironment activates β2-AR on the surfaces of various immune cells. The activation of these adrenergic receptors activates myeloid-derived suppresser cells (MDSCs) and recruits T-regulatory cells (Tregs) and M2 tumor-associated macrophages (TAMS). On the other hand, the activity of natural killer (NK) cells, T cells, and dendritic cells (DCs) is inhibited by β2-AR signaling.
4. Eminent Role of β2-AR in T Cells
Catecholamines can activate five distinct types of adrenergic receptors. Some of them have been reported to be expressed in immune cells. T lymphocytes express both α-AR and β-AR receptors [37][38]. The expression of α-AR on T lymphocytes has remained controversial, and there are various conflicting data on this issue. There are some studies which have completely denied the possibility of the presence of α-AR in T cell populations, while others have reported the expression of α-AR in lymphocyte populations upon LPS stimulation [39][40]. The conflict in these studies might be due to lack of availability of specific antibodies. The data regarding α-AR expression mostly rely on RT-PCR analysis, which is prone to contamination. Whatever the reason might be, the fact is that evidence regarding the role of α-AR in T lymphocytes is insufficient and less documented [41]. On the other hand, the role of β2-AR in T lymphocytes is well documented and has been the focus of much recent research [21][42][43].
The effects of NA on CD8+ T cells are primarily mediated by β2-adrenergic receptors (β2-AR) rather than other receptors of the same class. It was found that the α-AR antagonist phentolamine and the β1-AR-specific antagonist atenolol were unable to reverse the effects of NA in CD8+ T cells. This was demonstrated by the fact that phentolamine did not alter the expression of IL-1 and IL-6 genes under the influence of NA. Therefore, this research suggests that β2-AR plays a more exclusive role in the suppression of T cell receptor (TCR)-mediated human and mouse CD8+ T cell effector functions [44][45]. Moreover, the β2-AR specific blocker nadolol can abolish NA-mediated effects on CD8+ T cells. On the other hand, β2-AR-specific agonist terbutaline mimics the inhibitory actions of NA [45]. In a model of influenza virus, the role of beta-adrenergic receptors was assessed by pharmacological inhibition of β2-AR as well as chemical inhibition of SNS. Both of the aforementioned systems mimicked each other, signifying the importance and vitality of β2-AR signaling in T cell populations [46].
Pharmacological inhibition or genetic deletion of β2-AR in mice can improve anti-tumor immunity in mice models of colon and breast cancer under stress conditions. Inhibition of β2-AR signaling led to an improved number and function of CD8+ T cells, owing to better tumor control as compared to the control group [47]. The beneficial effects of β2-AR blockade (pharmacological or genetic) in the case of cancer depend upon CD8+ T cells [47][48]. CD8+ T cells, being the backbone of CAR-T therapy, are vital for anti-tumor immune responses. Certain studies have demonstrated that blocking β2-AR signaling can halt the growth of tumors. The mechanism of this phenomenon is relatively complex, and often, intrinsic signaling pathways of cancer cells are held responsible for adrenergic blockade-mediated anti-tumor responses. The poor prognosis of cancer might be due to adrenergic stress-driven loss of T cell activity. In the absence of CD8+ T cells, propranolol and β2-AR knockout (KO) mice do not exert their anti-tumor effects, which signifies that the β2-AR blockade, apart from affecting tumor intrinsic pathways, improves the status of anti-tumor immunity and, hence, leads to better anti-tumor control [49]. Overall, the evidence suggests that the inhibitory effects of NA on T cells are mainly due to the exclusive involvement of β2-AR, highlighting the importance of this receptor subtype in regulating immune responses.
References
- Lymperopoulos, A. Physiology and pharmacology of the cardiovascular adrenergic system. Front. Physiol. 2013, 4, 240.
- Behl, T.; Kaur, I.; Sehgal, A.; Singh, S.; Makeen, H.A.; Albratty, M.; Alhazmi, H.A.; Bhatia, S.; Bungau, S. The Locus Coeruleus—Noradrenaline system: Looking into Alzheimer’s therapeutics with rose coloured glasses. Biomed. Pharmacother. 2022, 151, 113179.
- Molina, P.E. Neurobiology of the stress response: Contribution of the sympathetic nervous system to the neuroimmune axis in traumatic injury. Shock 2005, 24, 3–10.
- Chrousos, G.P. Stress and disorders of the stress system. Nat. Rev. Endocrinol. 2009, 5, 374–381.
- Ouyang, X.; Zhu, Z.; Yang, C.; Wang, L.; Ding, G.; Jiang, F. Epinephrine increases malignancy of breast cancer through p38 MAPK signaling pathway in depressive disorders. Int. J. Clin. Exp. Pathol. 2019, 12, 1932–1946.
- Irwin, M.R.; Miller, A.H. Depressive disorders and immunity: 20 years of progress and discovery. Brain Behav. Immun. 2007, 21, 374–383.
- Graff, R.M.; Kunz, H.E.; Agha, N.H.; Baker, F.L.; Laughlin, M.; Bigley, A.B.; Markofski, M.M.; LaVoy, E.C.; Katsanis, E.; Bond, R.A.; et al. β2-Adrenergic receptor signaling mediates the preferential mobilization of differentiated subsets of CD8+ T-cells, NK-cells and non-classical monocytes in response to acute exercise in humans. Brain Behav. Immun. 2018, 74, 143–153.
- Takenaka, M.C.; Araujo, L.P.; Maricato, J.T.; Nascimento, V.M.; Guereschi, M.G.; Rezende, R.M.; Quintana, F.J.; Basso, A.S. Norepinephrine Controls Effector T Cell Differentiation through β2-Adrenergic Receptor–Mediated Inhibition of NF-κB and AP-1 in Dendritic Cells. J. Immunol. 2016, 196, 637–644.
- Xiao, L.; Li, X.; Fang, C.; Yu, J.; Chen, T. Neurotransmitters: Promising immune modulators in the tumor microenvironment. Front. Immunol. 2023, 14, 1118637.
- Silva, D.; Quintas, C.; Gonçalves, J.; Fresco, P. Contribution of adrenergic mechanisms for the stress-induced breast cancer carcinogenesis. J. Cell. Physiol. 2022, 237, 2107–2127.
- Croasdell, A.; Duffney, P.F.; Kim, N.; Lacy, S.H.; Sime, P.J.; Phipps, R.P. PPARγ and the innate immune system mediate the resolution of inflammation. PPAR Res. 2015, 2015, 549691.
- Apavaloaei, A.; Hardy, M.P.; Thibault, P.; Perreault, C. The origin and immune recognition of tumor-specific antigens. Cancers 2020, 12, 2607.
- Shimasaki, N.; Jain, A.; Campana, D. NK cells for cancer immunotherapy. Nat. Rev. Drug Discov. 2020, 19, 200–218.
- Ellis, G.I.; Sheppard, N.C.; Riley, J.L. Genetic engineering of T cells for immunotherapy. Nat. Rev. Genet. 2021, 22, 427–447.
- Maude, S.L.; Frey, N.; Shaw, P.A.; Aplenc, R.; Barrett, D.M.; Bunin, N.J.; Chew, A.; Gonzalez, V.E.; Zheng, Z.; Lacey, S.F.; et al. Chimeric antigen receptor T cells for sustained remissions in leukemia. N. Engl. J. Med. 2014, 371, 1507–1517.
- Zhang, C.; He, J.; Liu, L.; Wang, J.; Wang, S.; Liu, L.; Gao, L.; Gao, L.; Liu, Y.; Kong, P.; et al. CD19-directed fast CART therapy for relapsed/refractory acute lymphoblastic leukemia: From bench to bedside. Blood 2019, 134, 1340.
- Marofi, F.; Motavalli, R.; Safonov, V.A.; Thangavelu, L.; Yumashev, A.V.; Alexander, M.; Shomali, N.; Chartrand, M.S.; Pathak, Y.; Jarahian, M.; et al. CAR T cells in solid tumors: Challenges and opportunities. Stem Cell Res. Ther. 2021, 12, 1–16.
- Rahman, M.M.; Behl, T.; Islam, M.R.; Alam, M.N.; Islam, M.M.; Albarrati, A.; Albratty, M.; Meraya, A.M.; Bungau, S.G. Emerging Management Approach for the Adverse Events of Immunotherapy of Cancer. Molecules 2022, 27, 3798.
- Dogan, A.; Siegel, D.; Tran, N.; Fu, A.; Fowler, J.; Belani, R.; Landgren, O. B-cell maturation antigen expression across hematologic cancers: A systematic literature review. Blood Cancer J. 2020, 10, 73.
- Yan, T.; Zhu, L.; Chen, J. Current advances and challenges in CAR T-Cell therapy for solid tumors: Tumor-associated antigens and the tumor microenvironment. Exp. Hematol. Oncol. 2023, 12, 14.
- El-Sayes, N.; Vito, A.; Mossman, K. Tumor heterogeneity: A great barrier in the age of cancer immunotherapy. Cancers 2021, 13, 806.
- Li, W.; Song, X.; Jin, Y.; Li, F.; Yu, H.; Cao, C.; Jiang, Q. Carts for solid tumors: Feasible or infeasible? Oncol. Res. Treat. 2017, 40, 540–546.
- He, C.; Zhou, Y.; Li, Z.; Farooq, M.A.; Ajmal, I.; Zhang, H.; Zhang, L.; Tao, L.; Yao, J.; Du, B.; et al. Co-expression of IL-7 improves NKG2D-based CAR T cell therapy on prostate cancer by enhancing the expansion and inhibiting the apoptosis and exhaustion. Cancers 2020, 12, 1969.
- Gao, Y.; Lin, H.; Guo, D.; Cheng, S.; Zhou, Y.; Zhang, L.; Yao, J.; Farooq, M.A.; Ajmal, I.; Duan, Y.; et al. Suppression of 4.1 R enhances the potency of NKG2D-CAR T cells against pancreatic carcinoma via activating ERK signaling pathway. Oncogenesis 2021, 10, 62.
- Wrobel, L.J.; Gayet-Ageron, A.; Le Gal, F.A. Effects of beta-blockers on melanoma microenvironment and disease survival in human. Cancers 2020, 12, 1094.
- Lorton, D.; Bellinger, D.L. Molecular mechanisms underlying β-adrenergic receptor-mediated cross-talk between sympathetic neurons and immune cells. Int. J. Mol. Sci. 2015, 16, 5635–5665.
- Wang, W.; Cao, X. Beta-adrenergic signaling in tumor immunology and immunotherapy. Crit. Rev. Immunol. 2019, 39, 93–103.
- Kumar, V.; Patel, S.; Tcyganov, E.; Gabrilovich, D.I. The nature of myeloid-derived suppressor cells in the tumor microenvironment. Trends Immunol. 2016, 37, 208–220.
- Ammons, D.T.; MacDonald, C.R.; Chow, L.; Repasky, E.A.; Dow, S. Chronic Adrenergic Stress and Generation of Myeloid-Derived Suppressor Cells: Implications for Cancer Immunotherapy in Dogs. Vet. Comp. Oncol. 2023, 21, 159–165.
- Koinis, F.; Xagara, A.; Chantzara, E.; Leontopoulou, V.; Aidarinis, C.; Kotsakis, A. Myeloid-derived suppressor cells in prostate cancer: Present knowledge and future perspectives. Cells 2022, 11, 20.
- Ağaç, D.; Estrada, L.D.; Maples, R.; Hooper, L.V.; Farrar, J.D. The β2-adrenergic receptor controls inflammation by driving rapid IL-10 secretion. Brain Behav. Immun. 2018, 74, 176–185.
- Zahalka, A.H.; Arnal-Estapé, A.; Maryanovich, M.; Nakahara, F.; Cruz, C.D.; Finley, L.W.; Frenette, P.S. Adrenergic nerves activate an angio-metabolic switch in prostate cancer. Science 2017, 358, 321–326.
- Gardner, K.P.; Cristofanilli, M.; Chumsri, S.; Lapidus, R.; Tang, C.M.; Raghavakaimal, A.; Adams, D.L. Beta 2-Adrenergic Receptor in Circulating Cancer-Associated Cells Predicts for Increases in Stromal Macrophages in Circulation and Patient Survival in Metastatic Breast Cancer. Int. J. Mol. Sci. 2022, 23, 7299.
- Lamkin, D.M.; Ho, H.Y.; Ong, T.H.; Kawanishi, C.K.; Stoffers, V.L.; Ahlawat, N.; Ma, J.C.; Arevalo, J.M.; Cole, S.W.; Sloan, E.K. β-Adrenergic-stimulated macrophages: Comprehensive localization in the M1-M2 spectrum. Brain Behav. Immun. 2016, 57, 338–346.
- Thapa, S.; Cao, X. Nervous regulation: Beta-2-adrenergic signaling in immune homeostasis, cancer immunotherapy, and autoimmune diseases. Cancer Immunol. Immunother. 2023, 72, 2549–2556.
- Qiao, G. Understanding the Mechanisms by Which β-Adrenergic Stress Signaling Impairs T-Cell Immune Responses. Ph.D. Dissertation, State University of New York at Buffalo, Buffalo, NY, USA, 2021.
- Kavelaars, A. Regulated expression of α-1 adrenergic receptors in the immune system. Brain Behav. Immun. 2002, 16, 799–807.
- Kalinichenko, V.V.; Mokyr, M.B.; Graf, L.H.; Cohen, R.L.; Chambers, D.A. Norepinephrine-mediated inhibition of antitumor cytotoxic T lymphocyte generation involves a β-adrenergic receptor mechanism and decreased TNF-α gene expression. J. Immunol. 1999, 163, 2492–2499.
- Casale, T.B.; Kaliner, M. Demonstration that circulating human blood cells have no detectable alpha1-adrenergic receptors by radioligand binding analysis. J. Allergy Clin. Immunol. 1984, 74, 812–818.
- Van Der Voort, C.R.; Kavelaars, A.; Van De Pol, M.; Heijnen, C.J. Noradrenaline induces phosphorylation of ERK-2 in human peripheral blood mononuclear cells after induction of α1-adrenergic receptors. J. Neuroimmunol. 2000, 108, 82–91.
- Grisanti, L.A.; Perez, D.M.; Porter, J.E. Modulation of immune cell function by α1-adrenergic receptor activation. Curr. Top. Membr. 2011, 67, 113–138.
- Nissen, M.D.; Sloan, E.K.; Mattarollo, S.R. β-Adrenergic Signaling Impairs Antitumor CD8+ T-cell Responses to B-cell Lymphoma ImmunotherapyβAR Signaling Impairs T cell–Mediated Immunotherapy. Cancer Immunol. Res. 2018, 6, 98–109.
- Yang, C.; He, Y.; Chen, F.; Zhang, F.; Shao, D.; Wang, Z. Leveraging β-Adrenergic Receptor Signaling Blockade for Improved Cancer Immunotherapy Through Biomimetic Nanovaccine. Small 2023, 19, e2207029.
- Estrada, L.D.; Ağaç, D.; Farrar, J.D. Sympathetic neural signaling via the β2-adrenergic receptor suppresses T-cell receptor-mediated human and mouse CD8+ T-cell effector function. Eur. J. Immunol. 2016, 46, 1948–1958.
- Slota, C.; Shi, A.; Chen, G.; Bevans, M.; Weng, N.P. Norepinephrine preferentially modulates memory CD8 T cell function inducing inflammatory cytokine production and reducing proliferation in response to activation. Brain Behav. Immun. 2015, 46, 168–179.
- Grebe, K.M.; Hickman, H.D.; Irvine, K.R.; Takeda, K.; Bennink, J.R.; Yewdell, J.W. Sympathetic nervous system control of anti-influenza CD8+ T cell responses. Proc. Natl. Acad. Sci. USA 2009, 106, 5300–5305.
- Chen, M.; Qiao, G.; Hylander, B.L.; Mohammadpour, H.; Wang, X.Y.; Subjeck, J.R.; Singh, A.K.; Repasky, E.A. Adrenergic stress constrains the development of anti-tumor immunity and abscopal responses following local radiation. Nat. Commun. 2020, 11, 1821.
- Bucsek, M.J.; Qiao, G.; MacDonald, C.R.; Giridharan, T.; Evans, L.; Niedzwecki, B.; Liu, H.; Kokolus, K.M.; Eng, J.W.L.; Messmer, M.N.; et al. β-Adrenergic Signaling in Mice Housed at Standard Temperatures Suppresses an Effector Phenotype in CD8+ T Cells and Undermines Checkpoint Inhibitor Therapy. Cancer Res. 2017, 77, 5639–5651.
- Qiao, G.; Bucsek, M.J.; Winder, N.M.; Chen, M.; Giridharan, T.; Olejniczak, S.H.; Hylander, B.L.; Repasky, E.A. β-Adrenergic signaling blocks murine CD8+ T-cell metabolic reprogramming during activation: A mechanism for immunosuppression by adrenergic stress. Cancer Immunol. Immunother. 2019, 68, 11–22.
More
Information
Subjects:
Immunology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
712
Revisions:
2 times
(View History)
Update Date:
31 Aug 2023
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No