Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Horwitz, A.; Birk, R. Adipocyte Hyperplasia. Encyclopedia. Available online: https://encyclopedia.pub/entry/48267 (accessed on 26 July 2026).
Horwitz A, Birk R. Adipocyte Hyperplasia. Encyclopedia. Available at: https://encyclopedia.pub/entry/48267. Accessed July 26, 2026.
Horwitz, Avital, Ruth Birk. "Adipocyte Hyperplasia" Encyclopedia, https://encyclopedia.pub/entry/48267 (accessed July 26, 2026).
Horwitz, A., & Birk, R. (2023, August 21). Adipocyte Hyperplasia. In Encyclopedia. https://encyclopedia.pub/entry/48267
Horwitz, Avital and Ruth Birk. "Adipocyte Hyperplasia." Encyclopedia. Web. 21 August, 2023.
Copy Citation
Adipose tissue comprises various cell types, including mature adipocytes, stromal cells, fibroblasts, macrophages, blood cells, endothelial cells, smooth muscle cells, mesenchymal stem cells (MSCs), and APCs. APCs, resembling fibroblasts, can differentiate into different preadipocyte lineages (e.g., beige and white adipocytes) in response to genetic and environmental factors, contributing to adipose tissue hyperplasia expansion.
obesity
Bardet–Biedl syndrome (BBS)
hyperplasia
hypertrophy
1. Introduction
Obesity is a widespread chronic disease classified as a global pandemic. The prevalence of adult overweight and obesity has risen to nearly 40% and 10–15%, respectively, over the past 50 years [1][2]. The development of obesity involves a combination of genetic predisposition and environmental factors, where an individual’s unique genetic background interacts with obesogenic environments [3]. The primary cause of obesity is an imbalance between energy intake and expenditure, disrupting energy homeostasis and resulting in initial fat accumulation in white adipose tissue (WAT) and subsequent accumulation in other tissues. This fat accumulation leads to increased stress and dysfunction in tissues, contributing to the development of obesity-related comorbidities such as type 2 diabetes (T2D), cardiovascular disease, hypertension, liver dysfunction, and cancer, which collectively account for over 70% of premature deaths worldwide [3][4][5][6][7]. The economic burden of obesity includes both direct and indirect costs, including medical expenses, unemployment, and reduced socio-economic productivity [3]. The adipose tissue is distributed throughout the body in two main depots: subcutaneous WAT located beneath the skin, accounting for approximately 90% of total WAT, and visceral WAT located in the abdominal cavity, with the omental depot being the primary visceral fat depot in humans [8]. Central fat distribution around internal organs is associated with a higher risk of metabolic complications and inflammation.
2. Adipose tissue Hyperplasia and Hypertrophy
Obesity is a metabolic state generated by adipose tissue expansion, which can occur through hyperplasia (proliferation and differentiation of adipose precursor cells) and hypertrophy (increase in adipocyte size), as demonstrated in Scheme 1 [13]. The number of adipocytes in a specific fat depot is primarily established early in life and tends to remain stable throughout adulthood [14]. In contrast, adipocyte hypertrophy, caused by a constant surplus of energy, potential is remarkable and can reach a size increase of several hundred micrometers in diameter. Thus, adipose tissue expansion involves a complex interplay of factors including energy balance, genetics, and developmental processes [15]. Adipose tissue expansion can be divided into two main phases: prenatal and postnatal. During the prenatal phase, adipose tissue undergoes dynamic changes characterized by both hyperplasia and hypertrophy processes [16]. Upon gestational week 14, an early fat depot is starting to emerge from connective tissue. The primitive fat depots are composed of primary vessels and proliferating mesenchymal cells that subsequently will differentiate into preadipocytes [17]. Adipocytes begin to appear by the 23rd gestation week, and by the 28th gestation week, discernible fat lobule structures are formed. At the end of pregnancy, fat depots contain and encompass diverse groups of adipocyte subpopulations, primarily distinguished by their fat content and, consequently, their size [18][19]. The postnatal phase begins after birth and continues throughout life, with adipose tissue undergoing dynamic changes in response to energy balance and physiological needs by hyperplasia or hypertrophy [14]. Adipose tissue hyperplasia and hypertrophy depend on and vary with age, with rapid hyperplasia and hypertrophy during early childhood (0–2 years) and adolescence (12–18 years), and with relative hyperplasia stabilization at adulthood, as was shown in longitudinal and cross-sectional studies by Knittle et al. and others [20][21]. The rapid increase in both the hyperplasia and hypertrophy of adipose tissue occurs particularly in subcutaneous depots and is driven mainly by a growth hormone, insulin-like growth factor 1, sex steroids, and nutritional factors [22][23]. Notably, childhood obesity is characterized by accelerated adipose tissue hyperplasia, resulting in the estimated doubling of the adipocyte number compared to normal-weight counterparts, which significantly elevate the risk for the development of obesity in later life as well [23][24]. At normal weight states, the number of adipocytes peaks around puberty, and then stabilizes in adulthood. During adulthood, there is a gradual decline in adipose tissue hyperplasia potential with hypertrophy becoming predominant especially in visceral depots.
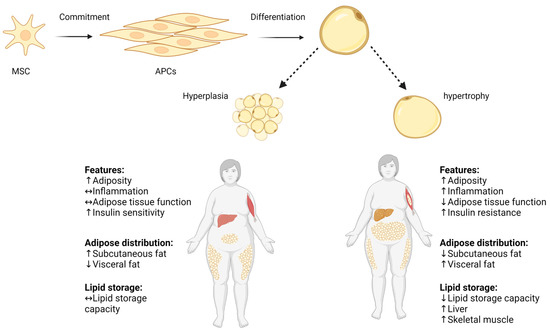
Scheme 1. Hyperplasia and hypertrophy in adipose tissue. Pluripotent MSC commit to differentiate into APC lineage located in the stromovascular fraction of adipose tissue, and APCs differentiate into mature adipocyte. Adipose tissue expands by increasing the volume of pre-existing adipocytes (adipose hypertrophy), and by generating new small adipocytes (hyperplasia). Increased adipocyte size correlates with impaired adipose tissue function, increased local inflammation, and decreased lipid storage capacity. ↑ increase, ↓ decrease, ↔ no change. Adapted from “Metabolically Unhealthy Obesity” and “Metabolically Healthy Obesity” by BioRender.com (2022). Retrieved from https://app.biorender.com/biorender-templates (accessed on 16 July 2023).
Earlier studies suggested that adulthood adipose tissue expansion is only through hypertrophic mechanisms [25][26]. However, recent lineage-tracing models in rodents indicate that under an excess caloric intake, preadipocyte cells can also differentiate into new adipocytes, contributing to adipose tissue expansion via hyperplasia [27][28]. Adulthood hyperplasia activation is possibly driven by the mature hypertrophic adipocytes’ limited oxygen diffusion capacity (which is at most 100 μm), and the neighboring cells’ mechanical stress [29].
Adipose tissue growth, development, and distribution are also sex-dependently mediated by sex hormones such as estrogen and testosterone [30][31]. For example, apart from estradiol fluctuation during the menstrual cycle, impacting appetite, caloric intake, and energy expenditure, it promotes subcutaneous fat accumulation, particularly in the gluteo-femoral region, resulting in the characteristic female pear-shaped body composition [32][33][34][35]. Conversely, testosterone favors the deposition of visceral fat, leading to the typical male apple-shaped body composition [36][37]. Moreover, subcutaneous adipose tissue usually has smaller more ‘plastic’ adipocytes, whereas the visceral adipose tissue is characterized by larger adipocytes encased in fibrotic tissues. Studies of rodents suggest that female mice present more adipose progenitor cells (APCs) in inguinal and gonadal depots compared to males, which respond to diet-induced obesity by elevating APCs’ hyperplasia (probably mediated by estrogen), while visceral fat expands with hypertrophy [8][28][38]. The precise mechanism by which sex hormones determine adipocyte hyperplasia or hypertrophy is still not fully known [30].
Hypoplasia is also a phenomenon related to adipose tissue, characterized by the underdevelopment or reduced growth of adipose tissue. Hypoplasia results from various factors, including genetic mutations, hormonal imbalances, ageing, or inadequate nutrient availability [39][40]. At the molecular level, adipose tissue hypoplasia can arise from differentiation, proliferation, and survival dysregulation through several key molecular regulators such as peroxisome-proliferator-activated receptor-gamma (PPARγ), CCAAT-Enhancer-Binding Proteins (C/EBPs), insulin-like growth factor 1 (IGF-1), and a fibroblast-growth-factor type (FGF). Additionally, inadequate nutrient availability, particularly essential fatty acids, can also limit adipose tissue growth [41][42][43][44].
3. Adipocyte Hyperplasia
Adipose tissue comprises various cell types, including mature adipocytes, stromal cells, fibroblasts, macrophages, blood cells, endothelial cells, smooth muscle cells, mesenchymal stem cells (MSCs), and APCs. APCs, resembling fibroblasts, can differentiate into different preadipocyte lineages (e.g., beige and white adipocytes) in response to genetic and environmental factors, contributing to adipose tissue hyperplasia expansion [45][46][47][48]. The pool of proliferating APCs is located in the stromovascular fraction of adipose tissue and is a depot and sex- and age-dependent [31][40][45][49][50]. Ex vivo and in vivo human studies have identified APC types among CD31− and CD34+ stromal vascular fractions [51][52][53][54][55][56][57]. Similarly, in a mice-specific sub-population characterized by cell surface immune markers was identified to possess proliferation and differentiation pro-adipogenic functional capabilities [51][53][54]. Yet, the final commitment to the adipogenic linkage is not fully known. Adipocytes’ steady-state turnover rate has been under investigation with no consensus to date. In a constant nutritional condition, the suggested estimate is a stable 10% turnover rate yearly [14]. This rate indicates that the maintenance of the adipocyte number normally involves a tightly regulated balance of adipogenesis and adipocyte death [58]. However, shown by several elegant studies, at any life stage, states of over-nutrition lead to adipose tissue hyperplasia and, consequently, to an increased mass [14][50][51][59]. Depot-dependent hyperplasia is a challenging research area due to technical issues; however, generally, it is accepted that a subcutaneous abdominal depot has a higher proliferative and differentiation capacity than a subcutaneous femoral depot and visceral adipose depot [60][61][62]. WAT depot distribution plays a critical role and is a stronger predictor of metabolic health risks than overall obesity. The accumulation of fat in the visceral adipose depot and subcutaneous abdominal depot confers a higher risk of developing T2D and cardiovascular disease, while the accumulation of subcutaneous gluteal and subcutaneous femoral fat may be metabolically protective [59][63][64].
Genetics also plays a role in an individual’s predisposition to adipose tissue hyperplasia, although the understanding on the subject is still evolving, and it is mostly investigated in animal models [65][66][67]. Genetic factors contribute to the development and maintenance of adipose tissue by controlling processes such as cell proliferation, differentiation, and apoptosis. One of the key genetic factors playing a role in adipose tissue hyperplasia regulation is leptin. Leptin-recessive mutated mice (Ob/Ob) lack leptin, consequently exhibiting hyper-phagic, obesity, hyper-insulinemia, and hyper-glycemia [68]. Furthermore, Ob/Ob mice present a larger number of adipose cells compared to the control, particularly being pronounced in the female mice [65]. Findings in other genetically obese strains, such as New Zealand (NZO), yellow (aAV), intermediate yellow (aAfY), and (db/db), in states of a high- and low-fat diet show variation in obesity development, suggesting that diet-induced adipose tissue hyperplasia is strain-dependent, and indicating an interaction between genetics and nutrition [69].
4. Adipocyte Hyperplasia in Obesity
During life, to replace mature adipocytes and under the condition of an excess energy flow, APC proliferate to generate new adipocytes [70][71]. The proliferation of other stromal cells parallels this process to ensure a sufficient blood flow to supply oxygen and nutrients to the growing tissue [72][73]. The number and size of adipocytes are critical factors in metabolic health. Many smaller adipocytes characterize a better healthy metabolic status, expressed by a better insulin sensitivity, a lower inflammation level, and less ectopic lipid accumulation [74][75][76]. For example, in a prospective study of Pima Indians with either a normal, impaired, or diabetic glucose tolerance, Weyer et al. demonstrated that a smaller adipocyte cell size was positively correlated with an improved glucose tolerance [76]. Enlarged adipocytes are typical of an obese state and correlate with the risk of metabolic syndrome (independent of BMI) [77][78][79]. For example, the overexpression of insulin-responsive glucose transporter (GLUT4) in mice results in obesity and the expansion of body fat only with hyperplasia, accompanied by an improved glucose tolerance [80]. Additionally, the collagen-IV-knock-out (KO) mice, characterized by unrestricted WAT hyperplasia under a positive energy balance due to the weakening of adipocytes’ extracellular scaffold, exhibit a significant weight gain yet an improved insulin sensitivity and inflammatory profile [81]. WAT hyperplasia occurs in a depot-specific manner, as was shown in diet-induced obese mice models [27][82][83]. WAT expansion in an intra-abdominal inguinal fat depot occurs almost exclusively through adipocyte hypertrophy [28], possibly due to the microenvironmental conditions that suppress the APCs’ potential to undergo adipogenesis [84]. In contrast, gonadal WAT can expand in both a hypertrophic and APC hyperplasic manner, triggered by nutrition and specific dietary lipids rather than the total caloric intake [22][82]. Hyperplasia involves complex, sequential, and molecular mechanisms and can be divided into two distinguished processes:
The commitment step—the multipotent precursor mesenchymal stem cells (MSC) differentiate into APCs, which are committed to differentiate into preadipocytes’ lineage. The commitment step is regulated by several signaling pathways and cytokine, as well as epigenetic modifications such as DNA hypomethylation, insulin, glucocorticoids, transforming growth factor β (TGFβ) superfamily members, bone morphogenetic proteins (BMPs), and wingless (WNT) family members [85][86][87]. The commitment stage is a complex process in which gene expression is precisely regulated and is extensively reviewed elsewhere [88][89][90].
The differentiation step—preadipocytes undergo growth arrest, accumulate lipids, and form functional insulin-responsive mature adipocytes [91]. The differentiation transcriptional regulation is a tightly regulated process, accompanied by transit and sequential expression at the level of different transcripts and proteins. This results in the progressive acquisition of morphological and biochemical characteristics of mature adipocytes [92]. Preadipocytes respond to combined mitogenic and adipogenic signals necessary for the subsequent differentiation steps. In the early stage of adipocyte differentiation, the expression of C/EBPβ and C/EBPδ increases, which upregulates C/EBPα expression, further activating PPARγ, the master regulator of adipocytes’ differentiation. PPARγ binds to the retinoic acid X receptor (RXR) to form heterodimers that bind to the PPARγ response element (PPRE) and initiate the transcription of downstream genes, including C/EBPα (positive feedback), to obtain mature adipocytes’ phenotype with the ability to accumulate fat and secret adipokines, such as leptin and adiponectin [93][94].
Adipose tissue hyperplasic expansion is a process regulated by hormones through endocrine, paracrine, autocrine, and neural systems [95][96][97]. Centrally, hormones and cytokines regulate satiety/hunger, metabolic, and activity states in a complex net of interactions. Major central players include glucagon-like peptide-1 (GLP-1), neuropeptide Y, leptin, ghrelin, and Cholecystokinin (CCK) [98]. Peripherally, molecular regulators of the adipose cell number include insulin, PPARγ ligands, retinoids, corticosteroids, and tumor necrosis factor-alpha (TNFα) [92][99][100]. Additionally, adipose tissue is innervated by sympathetic neurons, where APCs’ proliferation is highly responsive to β-adrenergic signaling [101].
States of dysregulation or unbalanced regulation of the complex cascade manifesting adipose tissue development can lead to accelerated adipocyte differentiation and hyperplasia [92][102]. For example, adipocytes are among the most insulin-responsive cell types, thus critically contributing to whole-body insulin sensitivity and energy homeostasis [103][104]. Furthermore, insulin is an obligatory hormone in preadipocyte differentiation to mature adipocytes in both in vivo and in vitro models [105][106]. Obesity induces an insulin resistance state, which is characterized by an adipocyte expansion blockage, possible death, and hyperplasia contributing to obesity co-morbidities, including T2D [107][108]. Growth factors such as insulin-like growth factor 1 (IGF-1) and fibroblast growth factor type 1 (FGF1) are crucial regulators of cell proliferation, survival, and differentiation and participate in the regulation of hyperplasia in adipose tissue [109][110][111]. Alteration in growth hormone levels can lead to the dysregulation of the activation of downstream signaling pathways such as the phosphatidylinositol 3-kinase (PI3K)/AKT and the mitogen-activated protein kinase (MAPK) pathways, which are known to stimulate cell growth and proliferation [112]. Several studies have shown negative correlations between BMI and absolute IGF-1 levels, while others reported increased circulating IGF-1 in obesity and, specifically, abdominal obesity due to elevated portal insulin levels [113][114][115][116][117].
References
- Seong, J.; Kang, J.Y.; Sun, J.S.; Kim, K.W. Hypothalamic Inflammation and Obesity: A Mechanistic Review. Arch. Pharm. Res. 2019, 42, 383–392.
- Hill, J.O.; Wyatt, H.R.; Peters, J.C. Energy Balance and Obesity. Circulation 2012, 126, 126–132.
- Apovian, C.M. Obesity: Definition, Comorbidities, Causes, and Burden. Am. J. Manag. Care 2016, 22, s176–s185.
- Cnop, M.; Foufelle, F.; Velloso, L.A. Endoplasmic Reticulum Stress, Obesity and Diabetes. Trends Mol. Med. 2012, 18, 59–68.
- Jung, U.; Choi, M.-S. Obesity and Its Metabolic Complications: The Role of Adipokines and the Relationship between Obesity, Inflammation, Insulin Resistance, Dyslipidemia and Nonalcoholic Fatty Liver Disease. Int. J. Mol. Sci. 2014, 15, 6184–6223.
- Haczeyni, F.; Wang, H.; Barn, V.; Mridha, A.R.; Yeh, M.M.; Haigh, G.W.; Ioannou, G.N.; Choi, Y.-J.; McWherter, C.A.; Teoh, N.C.-H.; et al. The Selective Peroxisome Proliferator–Activated Receptor-Delta Agonist Seladelpar Reverses Nonalcoholic Steatohepatitis Pathology by Abrogating Lipotoxicity in Diabetic Obese Mice. Hepatol. Commun. 2017, 1, 663–674.
- Bhaskaran, K.; Douglas, I.; Forbes, H.; dos-Santos-Silva, I.; Leon, D.A.; Smeeth, L. Body-Mass Index and Risk of 22 Specific Cancers: A Population-Based Cohort Study of 5·24 Million UK Adults. Lancet 2014, 384, 755–765.
- Wu, Y.; Lee, M.-J.; Ido, Y.; Fried, S.K. High-Fat Diet-Induced Obesity Regulates MMP3 to Modulate Depot- and Sex-Dependent Adipose Expansion in C57BL/6J Mice. Am. J. Physiol.-Endocrinol. Metab. 2017, 312, E58–E71.
- Tchernof, A.; Després, J.-P. Pathophysiology of Human Visceral Obesity: An Update. Physiol. Rev. 2013, 93, 359–404.
- Fried, S.K.; Bunkin, D.A.; Greenberg, A.S. Omental and Subcutaneous Adipose Tissues of Obese Subjects Release Interleukin-6: Depot Difference and Regulation by Glucocorticoid. J. Clin. Endocrinol. Metab. 1998, 83, 847–850.
- Mittelman, S.D.; Van Citters, G.W.; Kirkman, E.L.; Bergman, R.N. Extreme Insulin Resistance of the Central Adipose Depot In Vivo. Diabetes 2002, 51, 755–761.
- Gyllenhammer, L.E.; Lam, J.; Alderete, T.L.; Allayee, H.; Akbari, O.; Katkhouda, N.; Goran, M.I. Lower Omental T-Regulatory Cell Count Is Associated with Higher Fasting Glucose and Lower β-Cell Function in Adults with Obesity: Lower Omental Tregs and Type 2 Diabetes Risk. Obesity 2016, 24, 1274–1282.
- Arner, E.; Westermark, P.O.; Spalding, K.L.; Britton, T.; Rydén, M.; Frisén, J.; Bernard, S.; Arner, P. Adipocyte Turnover: Relevance to Human Adipose Tissue Morphology. Diabetes 2010, 59, 105–109.
- Spalding, K.L.; Arner, E.; Westermark, P.O.; Bernard, S.; Buchholz, B.A.; Bergmann, O.; Blomqvist, L.; Hoffstedt, J.; Näslund, E.; Britton, T.; et al. Dynamics of Fat Cell Turnover in Humans. Nature 2008, 453, 783–787.
- Kiess, W.; Petzold, S.; Töpfer, M.; Garten, A.; Blüher, S.; Kapellen, T.; Körner, A.; Kratzsch, J. Adipocytes and Adipose Tissue. Best Pract. Res. Clin. Endocrinol. Metab. 2008, 22, 135–153.
- Poissonnet, C.M.; Burdi, A.R.; Bookstein, F.L. Growth and Development of Human Adipose Tissue during Early Gestation. Early Hum. Dev. 1983, 8, 1–11.
- Votino, C.; Verhoye, M.; Segers, V.; Cannie, M.; Bessieres, B.; Cos, T.; Lipombi, D.; Jani, J. Fetal Organ Weight Estimation by Postmortem High-Field Magnetic Resonance Imaging before 20 Weeks’ Gestation. Ultrasound Obstet. Gynecol. 2012, 39, 673–678.
- Ali, A.T.; Hochfeld, W.E.; Myburgh, R.; Pepper, M.S. Adipocyte and Adipogenesis. Eur. J. Cell Biol. 2013, 92, 229–236.
- Scheidl, T.B.; Brightwell, A.L.; Easson, S.H.; Thompson, J.A. Maternal Obesity and Programming of Metabolic Syndrome in the Offspring: Searching for Mechanisms in the Adipocyte Progenitor Pool. BMC Med. 2023, 21, 50.
- Knittle, J.L.; Timmers, K.; Ginsberg-Fellner, F.; Brown, R.E.; Katz, D.P. The Growth of Adipose Tissue in Children and Adolescents. Cross-Sectional and Longitudinal Studies of Adipose Cell Number and Size. J. Clin. Investig. 1979, 63, 239–246.
- Sakers, A.; De Siqueira, M.K.; Seale, P.; Villanueva, C.J. Adipose-Tissue Plasticity in Health and Disease. Cell 2022, 185, 419–446.
- Meln, I.; Wolff, G.; Gajek, T.; Koddebusch, J.; Lerch, S.; Harbrecht, L.; Hong, W.; Bayindir-Buchhalter, I.; Krunic, D.; Augustin, H.G.; et al. Dietary Calories and Lipids Synergistically Shape Adipose Tissue Cellularity during Postnatal Growth. Mol. Metab. 2019, 24, 139–148.
- Landgraf, K.; Rockstroh, D.; Wagner, I.V.; Weise, S.; Tauscher, R.; Schwartze, J.T.; Löffler, D.; Bühligen, U.; Wojan, M.; Till, H.; et al. Evidence of Early Alterations in Adipose Tissue Biology and Function and Its Association with Obesity-Related Inflammation and Insulin Resistance in Children. Diabetes 2015, 64, 1249–1261.
- Efrat, M.; Tepper, S.; Birk, R.Z. From Fat Cell Biology to Public Health Preventive Strategies—Pinpointing the Critical Period for Obesity Prevention. J. Pediatr. Endocrinol. Metab. 2013, 26, 197–209.
- Salans, L.B.; Horton, E.S.; Sims, E.A.H. Experimental Obesity in Man: Cellular Character of the Adipose Tissue. J. Clin. Investig. 1971, 50, 1005–1011.
- Björntorp, P.; Sjöström, L. Number and Size of Adipose Tissue Fat Cells in Relation to Metabolism in Human Obesity. Metabolism 1971, 20, 703–713.
- Vishvanath, L.; MacPherson, K.A.; Hepler, C.; Wang, Q.A.; Shao, M.; Spurgin, S.B.; Wang, M.Y.; Kusminski, C.M.; Morley, T.S.; Gupta, R.K. Pdgfrβ+ Mural Preadipocytes Contribute to Adipocyte Hyperplasia Induced by High-Fat-Diet Feeding and Prolonged Cold Exposure in Adult Mice. Cell Metab. 2016, 23, 350–359.
- Wang, Q.A.; Tao, C.; Gupta, R.K.; Scherer, P.E. Tracking Adipogenesis during White Adipose Tissue Development, Expansion and Regeneration. Nat. Med. 2013, 19, 1338–1344.
- Trayhurn, P. Hypoxia and Adipose Tissue Function and Dysfunction in Obesity. Physiol. Rev. 2013, 93, 1–21.
- Palmer, B.F.; Clegg, D.J. The Sexual Dimorphism of Obesity. Mol. Cell. Endocrinol. 2015, 402, 113–119.
- Goossens, G.H.; Jocken, J.W.E.; Blaak, E.E. Sexual Dimorphism in Cardiometabolic Health: The Role of Adipose Tissue, Muscle and Liver. Nat. Rev. Endocrinol. 2021, 17, 47–66.
- Schreiner, P.J.; Terry, J.G.; Evans, G.W.; Hinson, W.H.; Crouse, J.R.; Heiss, G. Sex-Specific Associations of Magnetic Resonance Imaging-Derived Intra-Abdominal and Subcutaneous Fat Areas with Conventional Anthropometric Indices: The Atherosclerosis Risk in Communities Study. Am. J. Epidemiol. 1996, 144, 335–345.
- Lissner, L.; Stevens, J.; Levitsky, D.; Rasmussen, K.; Strupp, B. Variation in Energy Intake during the Menstrual Cycle: Implications for Food-Intake Research. Am. J. Clin. Nutr. 1988, 48, 956–962.
- Xu, Y.; Nedungadi, T.P.; Zhu, L.; Sobhani, N.; Irani, B.G.; Davis, K.E.; Zhang, X.; Zou, F.; Gent, L.M.; Hahner, L.D.; et al. Distinct Hypothalamic Neurons Mediate Estrogenic Effects on Energy Homeostasis and Reproduction. Cell Metab. 2011, 14, 453–465.
- Musatov, S.; Chen, W.; Pfaff, D.W.; Mobbs, C.V.; Yang, X.-J.; Clegg, D.J.; Kaplitt, M.G.; Ogawa, S. Silencing of Estrogen Receptor α in the Ventromedial Nucleus of Hypothalamus Leads to Metabolic Syndrome. Proc. Natl. Acad. Sci. USA 2007, 104, 2501–2506.
- Andersson, D.P.; Arner, E.; Hogling, D.E.; Rydén, M.; Arner, P. Abdominal Subcutaneous Adipose Tissue Cellularity in Men and Women. Int. J. Obes. 2017, 41, 1564–1569.
- Ramirez, M.E.; McMurry, M.P.; Wiebke, G.A.; Felten, K.J.; Ren, K.; Meikle, A.W.; Iverius, P.-H. Evidence for Sex Steroid Inhibition of Lipoprotein Lipase in Men: Comparison of Abdominal and Femoral Adipose Tissue. Metabolism 1997, 46, 179–185.
- Joe, A.W.B.; Yi, L.; Even, Y.; Vogl, A.W.; Rossi, F.M.V. Depot-Specific Differences in Adipogenic Progenitor Abundance and Proliferative Response to High-Fat Diet. Stem Cells 2009, 27, 2563–2570.
- Hammarstedt, A.; Gogg, S.; Hedjazifar, S.; Nerstedt, A.; Smith, U. Impaired Adipogenesis and Dysfunctional Adipose Tissue in Human Hypertrophic Obesity. Physiol. Rev. 2018, 98, 1911–1941.
- Kirkland, J.L.; Dobson, D.E. Preadipocyte Function and Aging: Links between Age-Related Changes in Cell Dynamics and Altered Fat Tissue Function. J. Am. Geriatr. Soc. 1997, 45, 959–967.
- Karagiannides, I.; Tchkonia, T.; Dobson, D.E.; Steppan, C.M.; Cummins, P.; Chan, G.; Salvatori, K.; Hadzopoulou-Cladaras, M.; Kirkland, J.L. Altered Expression of C/EBP Family Members Results in Decreased Adipogenesis with Aging. Am. J. Physiol. -Regul. Integr. Comp. Physiol. 2001, 280, R1772–R1780.
- Tchkonia, T.; Pirtskhalava, T.; Thomou, T.; Cartwright, M.J.; Wise, B.; Karagiannides, I.; Shpilman, A.; Lash, T.L.; Becherer, J.D.; Kirkland, J.L. Increased TNFα and CCAAT/Enhancer-Binding Protein Homologous Protein with Aging Predispose Preadipocytes to Resist Adipogenesis. Am. J. Physiol.-Endocrinol. Metab. 2007, 293, E1810–E1819.
- Xu, L.; Ma, X.; Verma, N.K.; Wang, D.; Gavrilova, O.; Proia, R.L.; Finkel, T.; Mueller, E. Ablation of PPARγ in Subcutaneous Fat Exacerbates Age-Associated Obesity and Metabolic Decline. Aging Cell 2018, 17, e12721.
- Ou, M.-Y.; Zhang, H.; Tan, P.-C.; Zhou, S.-B.; Li, Q.-F. Adipose Tissue Aging: Mechanisms and Therapeutic Implications. Cell Death Dis. 2022, 13, 300.
- Hepler, C.; Shan, B.; Zhang, Q.; Henry, G.H.; Shao, M.; Vishvanath, L.; Ghaben, A.L.; Mobley, A.B.; Strand, D.; Hon, G.C.; et al. Identification of Functionally Distinct Fibro-Inflammatory and Adipogenic Stromal Subpopulations in Visceral Adipose Tissue of Adult Mice. eLife 2018, 7, e39636.
- Iwayama, T.; Steele, C.; Yao, L.; Dozmorov, M.G.; Karamichos, D.; Wren, J.D.; Olson, L.E. PDGFRα Signaling Drives Adipose Tissue Fibrosis by Targeting Progenitor Cell Plasticity. Genes Dev. 2015, 29, 1106–1119.
- Sun, K.; Gao, Z.; Kolonin, M.G. Transient Inflammatory Signaling Promotes Beige Adipogenesis. Sci. Signal. 2018, 11, eaat3192.
- Sun, K.; Kusminski, C.M.; Scherer, P.E. Adipose Tissue Remodeling and Obesity. J. Clin. Investig. 2011, 121, 2094–2101.
- Sepe, A.; Tchkonia, T.; Thomou, T.; Zamboni, M.; Kirkland, J.L. Aging and Regional Differences in Fat Cell Progenitors—A Mini-Review. Gerontology 2011, 57, 66–75.
- Prins, J.B.; O’Rahilly, S. Regulation of Adipose Cell Number in Man. Clin. Sci. 1997, 92, 3–11.
- Berry, R.; Jeffery, E.; Rodeheffer, M.S. Weighing in on Adipocyte Precursors. Cell Metab. 2014, 19, 8–20.
- Sengenès, C.; Lolmède, K.; Zakaroff-Girard, A.; Busse, R.; Bouloumié, A. Preadipocytes in the Human Subcutaneous Adipose Tissue Display Distinct Features from the Adult Mesenchymal and Hematopoietic Stem Cells. J. Cell. Physiol. 2005, 205, 114–122.
- Rodeheffer, M.S.; Birsoy, K.; Friedman, J.M. Identification of White Adipocyte Progenitor Cells in Vivo. Cell 2008, 135, 240–249.
- Berry, R.; Rodeheffer, M.S. Characterization of the Adipocyte Cellular Lineage in Vivo. Nat. Cell Biol. 2013, 15, 302–308.
- Macdougald, O.A. Methods of Adipose Tissue Biology. In Methods in Enzymology, 1st ed.; Academic Press: Amsterdam, The Netherlands, 2014; ISBN 978-0-12-411619-1.
- Zenić, L.; Polančec, D.; Hudetz, D.; Jeleč, Z.; Rod, E.; Vidović, D.; Starešinić, M.; Sabalić, S.; Vrdoljak, T.; Petrović, T.; et al. Medicinal Signaling Cells Niche in Stromal Vascular Fraction from Lipoaspirate and Microfragmented Counterpart. Croat. Med. J. 2022, 63, 265–272.
- Frias, F.; Matos, B.; Jarnalo, M.; Freitas-Ribeiro, S.; Reis, R.L.; Pirraco, R.P.; Horta, R. Stromal Vascular Fraction Obtained from Subcutaneous Adipose Tissue: Ex-Obese and Older Population as Main Clinical Targets. J. Surg. Res. 2023, 283, 632–639.
- Arner, P.; Spalding, K.L. Fat Cell Turnover in Humans. Biochem. Biophys. Res. Commun. 2010, 396, 101–104.
- White, U.; Ravussin, E. Dynamics of Adipose Tissue Turnover in Human Metabolic Health and Disease. Diabetologia 2019, 62, 17–23.
- Hauner, H.; Entenmann, G. Regional Variation of Adipose Differentiation in Cultured Stromal-Vascular Cells from the Abdominal and Femoral Adipose Tissue of Obese Women. Int. J. Obes. 1991, 15, 121–126.
- Tchoukalova, Y.D.; Koutsari, C.; Votruba, S.B.; Tchkonia, T.; Giorgadze, N.; Thomou, T.; Kirkland, J.L.; Jensen, M.D. Sex- and Depot-Dependent Differences in Adipogenesis in Normal-Weight Humans. Obesity 2010, 18, 1875–1880.
- Tchkonia, T.; Giorgadze, N.; Pirtskhalava, T.; Tchoukalova, Y.; Karagiannides, I.; Forse, R.A.; DePonte, M.; Stevenson, M.; Guo, W.; Han, J.; et al. Fat Depot Origin Affects Adipogenesis in Primary Cultured and Cloned Human Preadipocytes. Am. J. Physiol. -Regul. Integr. Comp. Physiol. 2002, 282, R1286–R1296.
- Duque, A.P.; Rodrigues Junior, L.F.; Mediano, M.F.F.; Tibiriça, E.; De Lorenzo, A. Emerging Concepts in Metabolically Healthy Obesity. Am. J. Cardiovasc. Dis. 2020, 10, 48–61.
- Manolopoulos, K.N.; Karpe, F.; Frayn, K.N. Gluteofemoral Body Fat as a Determinant of Metabolic Health. Int. J. Obes. 2010, 34, 949–959.
- Johnson, P.R.; Hirsch, J. Cellularity of Adipose Depots in Six Strains of Genetically Obese Mice. J. Lipid Res. 1972, 13, 2–11.
- Johnson, P.R.; Zucker, L.M.; Cruce, J.A.F.; Hirsch, J. Cellularity of Adipose Depots in the Genetically Obese Zucker Rat. J. Lipid Res. 1971, 12, 706–714.
- Hirsch, J.; Batchelor, B. Adipose Tissue Cellularity in Human Obesity. Clin. Endocrinol. Metab. 1976, 5, 299–311.
- Szabadfi, K.; Pinter, E.; Reglodi, D.; Gabriel, R. Neuropeptides, Trophic Factors, and Other Substances Providing Morphofunctional and Metabolic Protection in Experimental Models of Diabetic Retinopathy. In International Review of Cell and Molecular Biology; Elsevier: Amsterdam, The Netherlands, 2014; Volume 311, pp. 1–121. ISBN 978-0-12-800179-0.
- Jo, J.; Gavrilova, O.; Pack, S.; Jou, W.; Mullen, S.; Sumner, A.E.; Cushman, S.W.; Periwal, V. Hypertrophy and/or Hyperplasia: Dynamics of Adipose Tissue Growth. PLoS Comput. Biol. 2009, 5, e1000324.
- Eckel-Mahan, K.; Ribas Latre, A.; Kolonin, M.G. Adipose Stromal Cell Expansion and Exhaustion: Mechanisms and Consequences. Cells 2020, 9, 863.
- Cinti, S. Between Brown and White: Novel Aspects of Adipocyte Differentiation. Ann. Med. 2011, 43, 104–115.
- Eto, H.; Suga, H.; Inoue, K.; Aoi, N.; Kato, H.; Araki, J.; Doi, K.; Higashino, T.; Yoshimura, K. Adipose Injury-Associated Factors Mitigate Hypoxia in Ischemic Tissues through Activation of Adipose-Derived Stem/Progenitor/Stromal Cells and Induction of Angiogenesis. Am. J. Pathol. 2011, 178, 2322–2332.
- Zhang, Y. Adipose Tissue-Derived Progenitor Cells and Cancer. World J. Stem Cells 2010, 2, 103.
- Strissel, K.J.; Stancheva, Z.; Miyoshi, H.; Perfield, J.W.; DeFuria, J.; Jick, Z.; Greenberg, A.S.; Obin, M.S. Adipocyte Death, Adipose Tissue Remodeling, and Obesity Complications. Diabetes 2007, 56, 2910–2918.
- Nishimura, S.; Manabe, I.; Nagasaki, M.; Seo, K.; Yamashita, H.; Hosoya, Y.; Ohsugi, M.; Tobe, K.; Kadowaki, T.; Nagai, R.; et al. In Vivo Imaging in Mice Reveals Local Cell Dynamics and Inflammation in Obese Adipose Tissue. J. Clin. Investig. 2008, 118, JCI33328.
- Weyer, C.; Foley, J.E.; Bogardus, C.; Tataranni, P.A.; Pratley, R.E. Enlarged Subcutaneous Abdominal Adipocyte Size, but Not Obesity Itself, Predicts Type II Diabetes Independent of Insulin Resistance. Diabetologia 2000, 43, 1498–1506.
- Gustafson, B.; Hedjazifar, S.; Gogg, S.; Hammarstedt, A.; Smith, U. Insulin Resistance and Impaired Adipogenesis. Trends Endocrinol. Metab. 2015, 26, 193–200.
- Hardy, O.T.; Perugini, R.A.; Nicoloro, S.M.; Gallagher-Dorval, K.; Puri, V.; Straubhaar, J.; Czech, M.P. Body Mass Index-Independent Inflammation in Omental Adipose Tissue Associated with Insulin Resistance in Morbid Obesity. Surg. Obes. Relat. Dis. 2011, 7, 60–67.
- Klöting, N.; Fasshauer, M.; Dietrich, A.; Kovacs, P.; Schön, M.R.; Kern, M.; Stumvoll, M.; Blüher, M. Insulin-Sensitive Obesity. Am. J. Physiol. Endocrinol. Metab. 2010, 299, E506–E515.
- Shepherd, P.R.; Gnudi, L.; Tozzo, E.; Yang, H.; Leach, F.; Kahn, B.B. Adipose Cell Hyperplasia and Enhanced Glucose Disposal in Transgenic Mice Overexpressing GLUT4 Selectively in Adipose Tissue. J. Biol. Chem. 1993, 268, 22243–22246.
- Khan, T.; Muise, E.S.; Iyengar, P.; Wang, Z.V.; Chandalia, M.; Abate, N.; Zhang, B.B.; Bonaldo, P.; Chua, S.; Scherer, P.E. Metabolic Dysregulation and Adipose Tissue Fibrosis: Role of Collagen VI. Mol. Cell Biol. 2009, 29, 1575–1591.
- Gao, Z.; Daquinag, A.C.; Su, F.; Snyder, B.; Kolonin, M.G. PDGFRα/PDGFRβ Signaling Balance Modulates Progenitor Cell Differentiation into White and Beige Adipocytes. Development 2017, 145, dev155861.
- Kim, S.M.; Lun, M.; Wang, M.; Senyo, S.E.; Guillermier, C.; Patwari, P.; Steinhauser, M.L. Loss of White Adipose Hyperplastic Potential Is Associated with Enhanced Susceptibility to Insulin Resistance. Cell Metab. 2014, 20, 1049–1058.
- Jeffery, E.; Wing, A.; Holtrup, B.; Sebo, Z.; Kaplan, J.L.; Saavedra-Peña, R.; Church, C.D.; Colman, L.; Berry, R.; Rodeheffer, M.S. The Adipose Tissue Microenvironment Regulates Depot-Specific Adipogenesis in Obesity. Cell Metab. 2016, 24, 142–150.
- Konieczny, S.F.; Emerson, C.P. 5-Azacytidine Induction of Stable Mesodermal Stem Cell Lineages from 10T1/2 Cells: Evidence for Regulatory Genes Controlling Determination. Cell 1984, 38, 791–800.
- Bowers, R.R.; Kim, J.W.; Otto, T.C.; Lane, M.D. Stable Stem Cell Commitment to the Adipocyte Lineage by Inhibition of DNA Methylation: Role of the BMP-4 Gene. Proc. Natl. Acad. Sci. USA 2006, 103, 13022–13027.
- Huang, H.; Song, T.-J.; Li, X.; Hu, L.; He, Q.; Liu, M.; Lane, M.D.; Tang, Q.-Q. BMP Signaling Pathway Is Required for Commitment of C3H10T1/2 Pluripotent Stem Cells to the Adipocyte Lineage. Proc. Natl. Acad. Sci. USA 2009, 106, 12670–12675.
- Farmer, S.R. Transcriptional Control of Adipocyte Formation. Cell Metab. 2006, 4, 263–273.
- Cawthorn, W.P.; Scheller, E.L.; MacDougald, O.A. Adipose Tissue Stem Cells Meet Preadipocyte Commitment: Going Back to the Future. J. Lipid Res. 2012, 53, 227–246.
- Li, S.-N.; Wu, J.-F. TGF-β/SMAD Signaling Regulation of Mesenchymal Stem Cells in Adipocyte Commitment. Stem Cell Res. Ther. 2020, 11, 41.
- Karastergiou, K.; Fried, S.K. Cellular Mechanisms Driving Sex Differences in Adipose Tissue Biology and Body Shape in Humans and Mouse Models. In Sex and Gender Factors Affecting Metabolic Homeostasis, Diabetes and Obesity; Mauvais-Jarvis, F., Ed.; Advances in Experimental Medicine and Biology; Springer International Publishing: Cham, Switzerland, 2017; Volume 1043, pp. 29–51. ISBN 978-3-319-70177-6.
- Jakab, J.; Miškić, B.; Mikšić, Š.; Juranić, B.; Ćosić, V.; Schwarz, D.; Včev, A. Adipogenesis as a Potential Anti-Obesity Target: A Review of Pharmacological Treatment and Natural Products. Diabetes Metab. Syndr. Obes. 2021, 14, 67–83.
- Ghaben, A.L.; Scherer, P.E. Adipogenesis and Metabolic Health. Nat. Rev. Mol. Cell Biol. 2019, 20, 242–258.
- Barilla, S.; Treuter, E.; Venteclef, N. Transcriptional and Epigenetic Control of Adipocyte Remodeling during Obesity. Obesity 2021, 29, 2013–2025.
- Lefterova, M.I.; Lazar, M.A. New Developments in Adipogenesis. Trends Endocrinol. Metab. 2009, 20, 107–114.
- Rosen, E.D.; Walkey, C.J.; Puigserver, P.; Spiegelman, B.M. Transcriptional Regulation of Adipogenesis. Genes Dev. 2000, 14, 1293–1307.
- Sarjeant, K.; Stephens, J.M. Adipogenesis. Cold Spring Harb. Perspect. Biol. 2012, 4, a008417.
- Cifuentes, L.; Acosta, A. Homeostatic Regulation of Food Intake. Clin. Res. Hepatol. Gastroenterol. 2022, 46, 101794.
- González-Casanova, J.E.; Pertuz-Cruz, S.L.; Caicedo-Ortega, N.H.; Rojas-Gomez, D.M. Adipogenesis Regulation and Endocrine Disruptors: Emerging Insights in Obesity. BioMed Res. Int. 2020, 2020, 7453786.
- Feinle-Bisset, C.; Horowitz, M. Appetite and Satiety Control—Contribution of Gut Mechanisms. Nutrients 2021, 13, 3635.
- Lee, Y.-H.; Petkova, A.P.; Mottillo, E.P.; Granneman, J.G. In Vivo Identification of Bipotential Adipocyte Progenitors Recruited by Β3-Adrenoceptor Activation and High-Fat Feeding. Cell Metab. 2012, 15, 480–491.
- Khan, F.; Khan, H.; Khan, A.; Yamasaki, M.; Moustaid-Moussa, N.; Al-Harrasi, A.; Rahman, S.M. Autophagy in Adipogenesis: Molecular Mechanisms and Regulation by Bioactive Compounds. Biomed. Pharmacother. 2022, 155, 113715.
- Newsholme, P.; Cruzat, V.; Arfuso, F.; Keane, K. Nutrient Regulation of Insulin Secretion and Action. J. Endocrinol. 2014, 221, R105–R120.
- Nahum, N.; Forti, E.; Aksanov, O.; Birk, R. Insulin Regulates Bbs4 during Adipogenesis: Insulin Regulates BBS Genes Transcripts. IUBMB Life 2017, 69, 489–499.
- Cho, H.J.; Park, J.; Lee, H.W.; Lee, Y.S.; Kim, J.B. Regulation of Adipocyte Differentiation and Insulin Action with Rapamycin. Biochem. Biophys. Res. Commun. 2004, 321, 942–948.
- Christoffersen, C.T.; Tornqvist, H.; Vlahos, C.J.; Bucchini, D.; Jami, J.; De Meyts, P.; Joshi, R.L. Insulin and Insulin-like Growth Factor-I Receptor Mediated Differentiation of 3T3-F442A Cells into Adipocytes: Effect of PI 3-Kinase Inhibition. Biochem. Biophys. Res. Commun. 1998, 246, 426–430.
- Sakaguchi, M.; Fujisaka, S.; Cai, W.; Winnay, J.N.; Konishi, M.; O’Neill, B.T.; Li, M.; García-Martín, R.; Takahashi, H.; Hu, J.; et al. Adipocyte Dynamics and Reversible Metabolic Syndrome in Mice with an Inducible Adipocyte-Specific Deletion of the Insulin Receptor. Cell Metab. 2017, 25, 448–462.
- Manna, P.; Jain, S.K. Obesity, Oxidative Stress, Adipose Tissue Dysfunction, and the Associated Health Risks: Causes and Therapeutic Strategies. Metab. Syndr. Relat. Disord. 2015, 13, 423–444.
- Grohmann, M.; Sabin, M.; Holly, J.; Shield, J.; Crowne, E.; Stewart, C. Characterization of Differentiated Subcutaneous and Visceral Adipose Tissue from Children. J. Lipid Res. 2005, 46, 93–103.
- Klöting, N.; Koch, L.; Wunderlich, T.; Kern, M.; Ruschke, K.; Krone, W.; Brüning, J.C.; Blüher, M. Autocrine IGF-1 Action in Adipocytes Controls Systemic IGF-1 Concentrations and Growth. Diabetes 2008, 57, 2074–2082.
- Salvatori, R. Growth Hormone Deficiency in Patients with Obesity. Endocrine 2015, 49, 304–306.
- Belfiore, A.; Frasca, F.; Pandini, G.; Sciacca, L.; Vigneri, R. Insulin Receptor Isoforms and Insulin Receptor/Insulin-like Growth Factor Receptor Hybrids in Physiology and Disease. Endocr. Rev. 2009, 30, 586–623.
- Sherlala, R.A.; Kammerer, C.M.; Kuipers, A.L.; Wojczynski, M.K.; Ukraintseva, S.V.; Feitosa, M.F.; Mengel-From, J.; Zmuda, J.M.; Minster, R.L. Relationship Between Serum IGF-1 and BMI Differs by Age. J. Gerontol. Ser. A 2021, 76, 1303–1308.
- Yamamoto, H.; Kato, Y. Relationship between Plasma Insulin-like Growth Factor I(IGF-I) Levels and Body Mass Index (BMI) in Adults. Endocr. J. 1993, 40, 41–45.
- Hjelholt, A.; Høgild, M.; Bak, A.M.; Arlien-Søborg, M.C.; Bæk, A.; Jessen, N.; Richelsen, B.; Pedersen, S.B.; Møller, N.; Lunde Jørgensen, J.O. Growth Hormone and Obesity. Endocrinol. Metab. Clin. N. Am. 2020, 49, 239–250.
- Serra, M.C.; Ryan, A.S.; Sorkin, J.D.; Favor, K.H.; Goldberg, A.P. High Adipose LPL Activity and Adipocyte Hypertrophy Reduce Visceral Fat and Metabolic Risk in Obese, Older Women: Adipocyte Hypertrophy and Metabolism. Obesity 2015, 23, 602–607.
- Rambold, A.S.; Cohen, S.; Lippincott-Schwartz, J. Fatty Acid Trafficking in Starved Cells: Regulation by Lipid Droplet Lipolysis, Autophagy, and Mitochondrial Fusion Dynamics. Dev. Cell 2015, 32, 678–692.
More
Information
Subjects:
Nutrition & Dietetics
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
3.1K
Revisions:
3 times
(View History)
Update Date:
28 Aug 2023
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No