Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Alexis Murillo Carrasco | -- | 3086 | 2023-08-10 12:09:51 | | | |
| 2 | Fanny Huang | Meta information modification | 3086 | 2023-08-11 08:11:16 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Murillo Carrasco, A.G.; Giovanini, G.; Ramos, A.F.; Chammas, R.; Bustos, S.O. Clustering Solid Tumors Based on Autophagy-Related Genes. Encyclopedia. Available online: https://encyclopedia.pub/entry/47898 (accessed on 25 July 2026).
Murillo Carrasco AG, Giovanini G, Ramos AF, Chammas R, Bustos SO. Clustering Solid Tumors Based on Autophagy-Related Genes. Encyclopedia. Available at: https://encyclopedia.pub/entry/47898. Accessed July 25, 2026.
Murillo Carrasco, Alexis Germán, Guilherme Giovanini, Alexandre Ferreira Ramos, Roger Chammas, Silvina Odete Bustos. "Clustering Solid Tumors Based on Autophagy-Related Genes" Encyclopedia, https://encyclopedia.pub/entry/47898 (accessed July 25, 2026).
Murillo Carrasco, A.G., Giovanini, G., Ramos, A.F., Chammas, R., & Bustos, S.O. (2023, August 10). Clustering Solid Tumors Based on Autophagy-Related Genes. In Encyclopedia. https://encyclopedia.pub/entry/47898
Murillo Carrasco, Alexis Germán, et al. "Clustering Solid Tumors Based on Autophagy-Related Genes." Encyclopedia. Web. 10 August, 2023.
Copy Citation
There has been a boost in autophagy reports due to its role in cancer progression and its association with tumor resistance to treatment. The availability of large cancer datasets has provided an extensive evidence-based approach to understanding the role of autophagy-related genes in various human cancers and their clinical implications, including cancer progression, development, and treatment response.
autophagy
cancer
gene
Data analysis
1. Introduction
To assess the clustering of solid tumors, researchers utilized the expression levels of autophagy-related genes. To ensure robust observations, researchers focused on solid tumor types with more than 100 tumor and control samples. Employing this approach, researchers generated a UMAP plot that revealed three distinct clusters among the sixteen solid tumors analyzed (Figure 1A). The identified clusters were as follows: Cluster 0 comprised BRCA, KIRC, KIRP, LGG, KIHC, LUAD, LUSC, and PRAD; Cluster 1 included COAD, ESCA, PAAD, and STAD; and Cluster 2 consisted of GBM, OV, SKCM, and TGCT. Notably, cluster 0 grouped tissues with similar genetic or anatomical profiles, such as BRCA-PRAD, KIRC-KIRP, or LUAD-LUSC. Cluster 1 predominantly encompassed gastrointestinal tumors, while Cluster 2 included TGCT and OV, which are tumors from reproductive organs. To identify differentially expressed genes characterizing these clusters, researchers identified 18 genes that primarily distinguished Clusters 0 and 1. Cluster 2 exhibited decreased levels of these markers (Figure 1B), and therefore it will not be analyzed in further detail herein. Nevertheless, it is important to note that autophagy profiles have been induced and studied on SKCM, GBM, and OV models with anti-tumoral effects [1][2][3][4]. In addition, for GBM and SKCM, there is possible to suggest that expression similarities in non-pivotal genes could be originated at their division from the ectoderm, as was demonstrated for the P2X7 receptor [5]. Then, these findings could support the evolutive hypothesis of cancer as an embryological phenomenon [6].
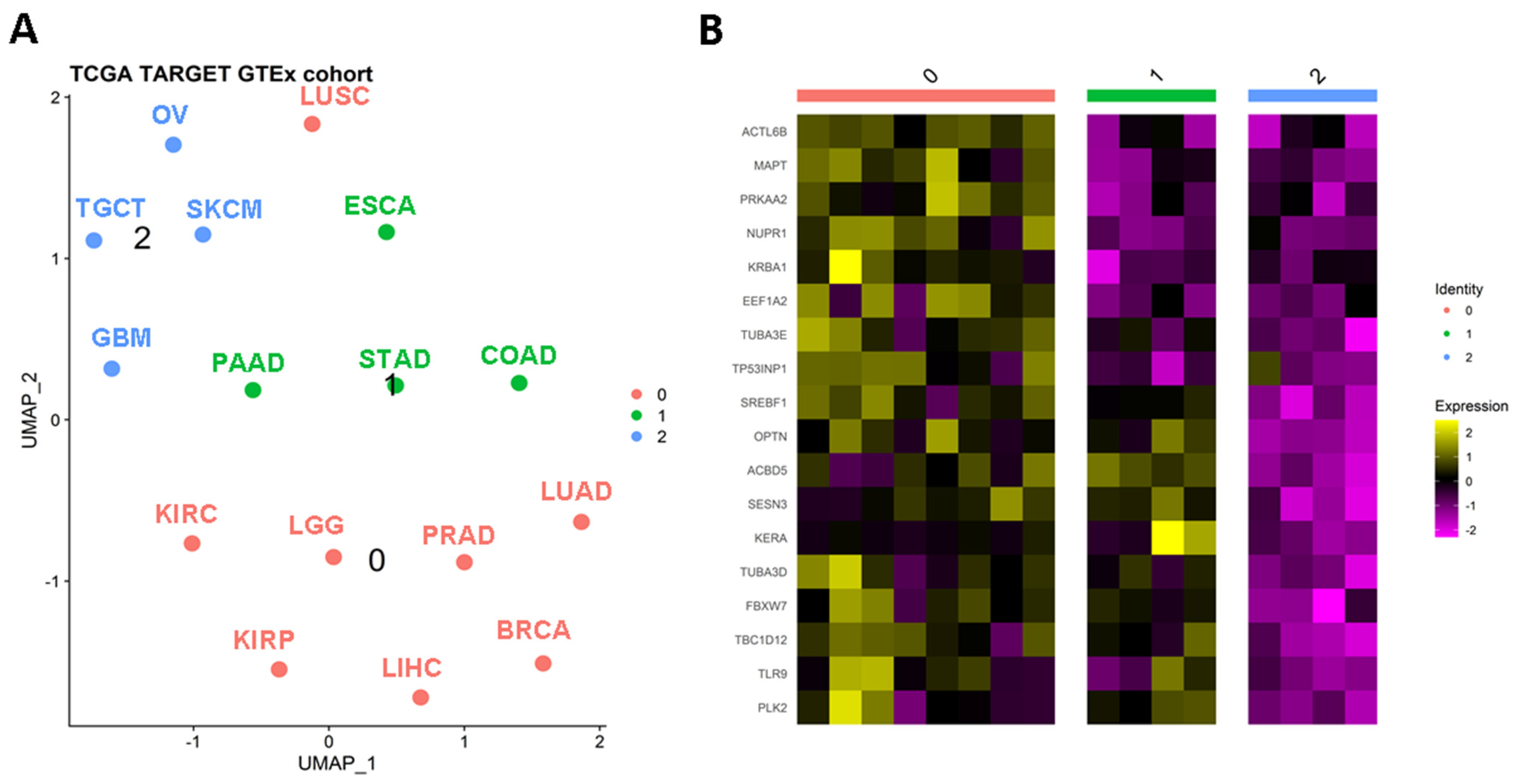
Figure 1. Autophagy-related genes can stratify solid tumors. (A) Clusterization of solid tumors based on the differential expression of autophagy genes. After a UMAP analysis, it is possible to recognize three classifications (B) of relevant tumors based on the expression of autophagy genes. BRCA: breast invasive carcinoma; COAD: colon adenocarcinoma; ESCA: esophageal carcinoma; GBM: glioblastoma multiforme; KIRC: kidney renal clear cell carcinoma; KIRP: kidney renal papillary cell carcinoma; LGG: brain lower grade glioma; LIHC: liver hepatocellular carcinoma; LUAD: lung adenocarcinoma; LUSC: lung squamous cell carcinoma; OV: ovarian serous cystadenocarcinoma; PAAD: pancreatic adenocarcinoma; PRAD: prostate adenocarcinoma; SKCM: skin cutaneous melanoma; STAD: stomach adenocarcinoma; TGCT: testicular germ cell tumors.
2. Autophagy Regulators Specific to Cluster 0
Researchers' analysis revealed that the genes ACTL6B, MAPT, PRKAA2, NUPR1, KRBA1, EEF1A2, TUBA3E, and TP53INP1 specifically characterized cluster “0” through their overexpression. Furthermore, researchers observed that these genes exhibited upregulated levels in tumors belonging to Cluster “0” compared to their normal adjacent tissues. While the limited number of normal-adjacent samples in the TCGA data introduces potential biases and limitations, the majority of the putative markers for Cluster “0” could be validated using the UALCAN tool [7].
2.1. Protein and Mutational Features of Relevant Genes for Cluster 0
To gain insight into the protein products of these genes, researchers utilized the UALCAN tool to examine their change in proteins with data from the Clinical Proteomic Tumor Analysis Consortium (CPTAC) cohort. In the LIHC dataset, researchers found a comparable group of normal-adjacent samples, which demonstrated the upregulation of EEF1A2, NUPR1, and MAPT proteins in the tumor group. However, there were some inconsistencies. For instance, while the PRKAA2 gene exhibited notable overexpression in LIHC, its corresponding protein was significantly downregulated. This disparity suggests the importance of considering the mutational profile of these genes or post-translational events on the produced proteins. Based on data from cBioPortal [8] of the TCGA datasets belonging to Cluster “0”, these genes exhibited a low frequency of mutations (1.1–5%), implying that somatic mutations may not play a significant role in dysregulating the relationship between autophagy-related genes and their corresponding proteins.
2.2. Previous Research on Relevant Genes for Cluster 0
According to the MSigDB, the NUPR1 and PRKAA2 genes participate in macroautophagy and its regulation, while MAPT serves as an autophagy regulator, TP53INP1 contributes to autophagosome organization, and ACTL6B and KRBA1 are involved in the autophagy-related network. Additionally, other genes are associated with less-studied forms of autophagy. For example, EEF1A2 is linked to chaperone-mediated autophagy and its regulation, PRKAA2 is involved in lipophagy-related pathways, and TUBA3E is associated with aggrephagy.
Previous studies have highlighted the significant role of NUPR1 in macroautophagy and its impact on the aggressiveness and treatment resistance of specific tumors such as BRCA, LUAD, LUSC, LIHC, and LGG [9][10][11][12][13][14][15]. NUPR1, also known as p8, is a transcriptional regulator that has been shown to reduce apoptosis caused by dihydroartemisinin (DHA), sorafenib, or ionizing radiation (IR) in LIHC tumor cells [9][10][11]. However, opposing effects have been observed in osteosarcoma and non-tumor cells [16]. Additionally, research has demonstrated that Δ9-tetrahydrocannabinol (THC) induces autophagy-mediated apoptosis in an LGG model [12]. Despite autophagy-related pathways being upregulated in LIHC and LGG tumors, it remains uncertain whether these pathways promote tumor growth or tumor suppression, necessitating further investigation. In lung and breast cancers (LUAD, LUSC, and BRCA), repression of NUPR1, in combination with conventional anticancer therapies, has been proven to control tumor growth [13][14][15]. Another study supports the inhibition of NUPR1 using microRNA-637 (hsa-miR-637) as a promising option for this purpose [17].
Controversial results have emerged regarding the expression of the PRKAA2 gene (Protein Kinase AMP-Activated Catalytic Subunit α 2, AMPKα2) in gastrointestinal malignancies [18][19][20][21]. Some studies suggest that repression of PRKAA2 promotes tumor growth in gastrointestinal cancer by suppressing ferroptosis, an autophagy-dependent form of cell death [18]. On the other hand, other studies propose that PRKAA2 activates autophagy-related pathways, leading to treatment resistance, and that its activation can be triggered by the gastrin hormone [19][20][21]. In LIHC, inhibiting PRKAA2 has been shown to downregulate autophagy rates, and metformin has been identified as a potential PRKAA agonist for controlling hepatitis C virus (HCV) replication [22]. In glioma, low expression of PRKAA2 has been associated with a better prognosis [23]. Although these findings may seem contradictory, especially considering that LGG belongs to Cluster “0”, they underscore the importance of including autophagy-related factors in the intra- and inter-individual heterogeneity of tumors.
The MAPT gene encodes the microtubule-associated protein tau, which has been extensively studied in Alzheimer’s disease (AD) [24][25]. Recent research has explored the interplay between autophagy and MAPT in AD and has demonstrated that overexpression of MAPT/tau inhibits the fusion of autophagosomes with lysosomes, leading to autophagosome accumulation through increased levels of LC3 protein [26]. Although direct links between MAPT and autophagy in cancer remain limited, the high expression levels of Tau protein in glioblastoma, a tumor with enhanced autophagy activity, have raised questions about its possible role in oncogenesis and its implications for cancer therapy [27].
In BRCA cohorts, a long non-coding RNA (lncRNA) for the MAPT gene called MAPT-AS1 has been found to be overexpressed in tumor tissues [28][29][30]. This is noteworthy because lncRNAs, which are usually located in antisense strands of DNA from original genes, can also be affected by somatic mutations irrespective of their canonical effects. Thus, the combination of somatic mutations and non-coding RNA as potential prognostic markers deserves further attention, as demonstrated in COAD [31].
TP53INP1 gene exhibits inconsistent findings across experiments and tumor tissues. Some researchers have identified hsa-miR-106a as an oncomiR that targets TP53INP1 in metastatic lung cancer [32], indicating its involvement in tumor suppression. Increasing the levels of TP53INP1 could be crucial in controlling tumor growth through autophagy-dependent cell death. In the case of PRAD, hsa-miR-30a and hsa-miR-205 have been suggested as potential therapeutic options for suppressing TP53INP1 [33][34]. However, it has been explained that TP53INP1 is overexpressed as a response to ionizing radiation, which confers resistance [33][34]. Therefore, suppressing this gene could potentially resensitize tumor cells to standard treatment protocols. Like other representative genes in this cluster, TP53INP1 exhibits a dual function. According to Peuget et al. (2021), oxidative stress induces the expression of TP53INP1 [35]. This stress can trigger autophagy by interacting with LC3 in the cytoplasm or apoptosis by interacting with P53 in the nucleus, and the role of mitochondria and their metabolism in this process is also implicated [36]. Thus, an additional factor to consider in researchers' analysis is the localization of autophagy-related transcripts and proteins. Unfortunately, there is insufficient information available to conduct this type of comparison.
In summary, NUPR1, PRKAA2, TP53INP1, ACTL6B, KRBA1, EEF1A2, and MAPT genes are coexpressed with 17 other genes (ANK2, ST8SIA1, GUCY2F, HERC1, TRHR, COL11A1, CHRM3, CNR2, KITLG, ROR1, CDKL5, PPOX, IGF2R, DDIT3, OPCML, ELOVL5, and BRINP2) according to the GeneMania database [37]. These genes are enriched in the MAPK pathway (p = 0.004) [38], allowing us to associate cluster “0” with a MAPK-dependent macroautophagy-like process. However, it is important to note the significant heterogeneity observed in the samples, classifications, tumor tissues, and other forms of autophagy.
3. Tumors Balancing Macro- and Micro-Autophagy Processes (Clusters 0 and 1)
Clusters “0” and “1” in Figure 1 represent a distinct group of genes associated with tumors that exhibit a balance between macroautophagy and microautophagy processes. Notably, the genes SREBF1, OPTN, ACBD5, SESN3, KERA, TUBA3D, FBXW7, TBC1D12, TLR9, and PLK2 show high expression levels in various tumors such as ESCA, PAAD, STAD, COAD, LUAD, LUSC, KIRC, LGG, PRAD, KIRP, LIHC, and BRCA.
In addition, after performing a random forest Gini importance analysis, researchers observed that KERA, TP53INP1, SREBF1, and TUBA3E showed great accuracy (above 75%) and over 75% of Gini contribution. It suggests the potential contribution of these autophagy-related genes in the classification of tissues regarding their dysregulation between tumor and normal samples.
Of particular interest are the TUBA3D and FBXW7 genes, which are associated with the chaperone-mediated protein folding pathway (R-HSA-390466) according to the Enrichr database [38]. This suggests their potential involvement in chaperone-mediated autophagy. Supporting this idea, these genes have also been implicated in certain forms of microautophagy, such as aggrephagy and mitophagy, as indicated by the MSigDB. Additionally, these genes are part of the regulatory pathways of macroautophagy along with the other eight genes that cluster these tumors. ACBD5, SREBF1, and OPTN genes are also involved in microautophagy pathways, including aggrephagy, mitophagy, and xenophagy.
3.1. Accumulation of ACBD5 Is Found in Tumors from Cluster 0 and 1
Notably, the ACBD5 gene is interesting in autophagy-related studies as its deregulation can induce their accumulation at protein levels. This gene has been associated with peroxisome maintenance, lipid exchange, and homeostasis, which are crucial processes for lipid and carbohydrate metabolism reorganization in tumor cells [39][40]. These processes involve microautophagy pathways such as pexophagy, aggrephagy, and mitophagy [41].
3.2. Previous Research on Overexpressed Genes in Tumors of Clusters 0 and 1
Other genes related to microautophagy processes include PLK2, SESN3, TLR9, OPTN, and SREBF1. Independent research has demonstrated that the PLK2 gene controls α-Synuclein aggregation in an autophagy-dependent context [42]. Although this process is dependent on macroautophagy and regulated by mTORC1 inhibition, it appears to be a microautophagy pathway that is specifically activated in the presence of its substrate, α-Synuclein [42][43]. An interesting regulatory axis involves the lncRNA OIP5-AS1, which targets hsa-miR-126 to prevent α-Synuclein aggregation in autophagy-activated cells [43].
Regarding the SESN3 gene, recent studies have identified its role as an autophagy activator in tumor cells by repressing mTORC1 [44]. However, this gene has also been associated with other autophagy pathways such as chaperone-mediated autophagy [45]. Overexpression of SESN3 has been observed in LUAD [45] and ESCA [46] models, suggesting its potential involvement in promoting pro-tumor autophagy pathways. Expression levels of this gene can be regulated by specific miRNAs, such as hsa-miR-194-3p [45] or hsa-miR-429 [46].
About mitophagy, several reports have described the upregulation of the TLR9 gene in tumors belonging to Clusters “0” and “1” [47][48][49], indicating its involvement in inducing this form of autophagy. In BRCA, it has been reported that this gene plays a role in the rewiring of doxorubicin and may explain the cardiomyocyte death and systolic dysfunction observed in patients undergoing this tumor treatment [50]. Consistent with these findings, TLR9 was found to be upregulated in aggressive versions of LIHC, LUAD, LUSC, and COAD models [51][52][53]. Consequently, various regulatory pathways have been proposed to control TLR9 expression. For example, hsa-miR-30a has been shown to sensitize LIHC cells to a combined therapy of hydroxychloroquine and sorafenib by repressing TLR9 [51]. On the other hand, inducing TLR9 expression in dendritic cells has been suggested as a potential therapeutic strategy, as demonstrated in PAAD cases [54]. It is important to note that bulk analyses using next-generation sequencing (NGS) do not differentiate between the origins of cells within tumors, which can lead to different interpretations of the results. Therefore, researchers are increasingly turning to single-cell sequencing to differentiate immune cells, tumor cells, and normal-adjacent cells with varying autophagy-related profiles within the same tumor pool.
In addition to TLR9, OPTN has been extensively studied in the context of mitophagy. PINK1 and PRKN, which are highly studied autophagy-related genes, are also involved in this process. The PHB2 gene stabilizes PINK1 in mitochondria, facilitating the recruitment of Parkin (the product of PRKN), ubiquitin, and optineurin (the product of OPTN) to promote mitophagy [55][56][57][58]. However, a recent study challenges the necessity of PINK1 and PRKN for initiating mitophagy [59]. Consequently, it has been suggested that OPTN may have tumor suppressor functions by activating suppressor autophagy mediated by HACE1, a tumor suppressor [60][61][62], or by repressing the pro-oncogenic transforming growth factor-β (TGFβ) signaling in triple-negative breast cancer (TNBC) cells, a subtype of BRCA [63]. Importantly, OPTN has been found to be downregulated in GBM tumor samples, which has been corroborated by independent studies [64]. The same study proposes that inducing OPTN expression in GBM cells could help control tumor growth, supporting a suppressive role for this gene, although the underlying mechanism remains unknown.
In terms of the application of OPTN in the context of mitophagy and the tumor environment, several studies have identified OPTN as a potential therapeutic target. For instance, it has been observed that OPTN induces pro-tumor mitochondrial-related autophagy, reducing the efficacy of combined treatments involving pemetrexed, cisplatin, and MEK inhibitors or anti-PD-L1 in a LUSC model [52]. In a PAAD model, repression of OPTN leads to apoptosis through chaperone-mediated autophagy [65]. Similar to TLR9, understanding the function of OPTN allows us to differentiate its contribution to tumor growth based on its expression in surrounding cells. In LUAD models, higher expression of OPTN in fibroblasts surrounding the tumor contributes to tumor invasiveness [66].
SREBF1 upregulation has been linked to mTORC1-dependent autophagy, which may be induced by leptins to suppress ferroptosis in BRCA, LIHC, PRAD, and LUAD models [67][68][69][70]. Additionally, SREBF1 levels were found to be elevated in PAAD tissues, regulated by high glucose concentrations. In PAAD models, the upregulation of SREBF1 helps control autophagy levels [71]. This gene may act as a negative regulator of mTORC1-dependent autophagy, favoring pro-tumor microautophagy pathways. It is worth noting that SREBF1 can function as both a protein and a transcription factor. Studies have demonstrated that genes upregulated by the SREBF1 transcription factor can be altered in the presence of cisplatin, inducing treatment resistance in a LUSC model [72]. This evidence highlights the importance of carefully analyzing autophagy-related genes with dual functions to enhance researchers' understanding of this process. A study proposed that mTORC2 stabilizes SREBF1 through FBXW7-mediated regulation to integrate autophagy and lipid metabolism processes, leading to the downregulation of target genes such as acetyl-CoA carboxylase and fatty-acid synthase [73].
Considering the combined findings of two genes involved in tumor clusterization, FBXW7 and SREBF1, it is hypothesized that these tumors exchange autophagy-related processes and large-scale technologies based on their aggressiveness and treatment sensitivity or resistance. However, conducting large-scale high-throughput analyses in mass groups could obscure specific autophagy pathways in certain tumor subtypes or patients. Therefore, the current perspective is to compare global observations with focused research. Nevertheless, the scientific community is moving towards a comprehensive analysis of tumors, considering their heterogeneity and subclonal profile, which will allow us to confirm researchers' current hypotheses about autophagy-related processes in the tumor environment in the future.
Regarding macroautophagy, the FBXW7 gene has been the focus of numerous studies aiming to characterize its function. This gene is known as a tumor suppressor as it is frequently mutated or suppressed in human tumors [74]. However, its dysregulation in chemoresistance remains controversial, suggesting that its behavior depends on the context. It has been observed to be upregulated in resistant gastric cells [75] and downregulated in chemoresistant models of BRCA [76].
Interestingly, FBXW7 has been found to induce the expression of ATG16L1, an important gene involved in LC3 lipidation and autophagosome formation, while not affecting the levels of other autophagy-related genes (ATG) [77]. Moreover, FBXW7 suppresses mTORC1, thereby activating autophagy pathways [78][79]. FBXW7 participates in different molecular axes, resulting in different effects on tumor cells. For instance, the GSK3-FBXW7 interaction leads to the ubiquitination and degradation of Rictor, increasing cellular ROS (reactive oxygen species) in an autophagy-activated context [80]. On the other hand, interactions between FBXW7 and oncogenes such as SHOC2 or LSD1 can reduce the expression of autophagy-related pathways [78][79][81]. In conditions where tumors are growing, cisplatin treatments have been shown to induce the degradation of the MRE11-RAD50-NBS1 (MRN) complex by FBXW7 and lysosomes [74]. As a result, the overexpression of the MRN complex or the suppression of the FBXW7 gene can lead to cisplatin-resistant tumors and a poor prognosis. In relation to this, hsa-miR-25 and hsa-miR-223 have been shown to suppress FBXW7 levels, promoting autophagy and treatment resistance in LIHC [82] and LUAD [83] models, respectively. Anti-miRs could be used to counteract the suppression of FBXW7 levels, but it is important to better understand the specific context in which this strategy would be applicable.
Lastly, three genes (TBC1D12, KERA, and TUBA3D) that contribute to tumor clustering in groups “0” and “1” have not been previously associated with the tumor-related autophagy process. It is important to emphasize that, in researchers' analysis, the KERA gene was the top gene in Gini relevance and accuracy in tissue pooling of groups between 0 + 1 vs. 2. Studies on mutations in the TBC1D12 gene (TBC1 Domain Family Member 12) have been conducted in urological tumors, suggesting that alterations in its mutational profile could be linked to worse patient survival [84]. Interestingly, this gene exhibits a higher mutation frequency in PRAD samples compared to other patients. The KERA (Keratan Sulfate Proteoglycan Keratocan) gene has been found to have lower levels in cisplatin and paclitaxel-resistant OV models [85], partially aligning with observations in the entire dataset (Cluster “2”). The expression levels of the TUBA3D (Tubulin α-3D Chain) gene in BRCA (upregulated) and OV (downregulated) have been validated [86][87]. Notably, in BRCA models, TUBA3D was shown to be downregulated in paclitaxel-resistant cells compared to parental cells [88].
In summary, the findings presented in this discussion suggest that all the aforementioned genes may make significant contributions to tumor-related autophagy through their expression in tumors and the surrounding cells, warranting further attention in future research.
References
- Chatterjee, S.J.; Pandey, S. Chemo-resistant melanoma sensitized by tamoxifen to low dose curcumin treatment through induction of apoptosis and autophagy. Cancer Biol. Ther. 2011, 11, 216–228.
- Matteoni, S.; Matarrese, P.; Ascione, B.; Ricci-Vitiani, L.; Pallini, R.; Villani, V.; Pace, A.; Paggi, M.G.; Abbruzzese, C. Chlorpromazine induces cytotoxic autophagy in glioblastoma cells via endoplasmic reticulum stress and unfolded protein response. J. Exp. Clin. Cancer Res. 2021, 40, 347.
- Zhan, L.; Zhang, Y.; Wang, W.; Song, E.; Fan, Y.; Li, J.; Wei, B. Autophagy as an emerging therapy target for ovarian carcinoma. Oncotarget 2016, 7, 83476–83487.
- Al-Qatati, A.; Aliwaini, S. Combined pitavastatin and dacarbazine treatment activates apoptosis and autophagy resulting in synergistic cytotoxicity in melanoma cells. Oncol. Lett. 2017, 14, 7993–7999.
- Li, X.; Qi, X.; Zhou, L.; Fu, W.; Abdul-Karim, F.W.; MacLennan, G.; Gorodeski, G.I. P2X7 receptor expression is decreased in epithelial cancer cells of ectodermal, uro-genital sinus, and distal paramesonephric duct origin. Purinergic Signal. 2009, 5, 351–368.
- Cofre, J.; Abdelhay, E. Cancer Is to Embryology as Mutation Is to Genetics: Hypothesis of the Cancer as Embryological Phenomenon. Sci. World J. 2017, 2017, 3578090.
- Chandrashekar, D.S.; Karthikeyan, S.K.; Korla, P.K.; Patel, H.; Shovon, A.R.; Athar, M.; Netto, G.J.; Qin, Z.S.; Kumar, S.; Manne, U.; et al. UALCAN: An update to the integrated cancer data analysis platform. Neoplasia 2022, 25, 18–27.
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio Cancer Genomics Portal: An Open Platform for Exploring Multidimensional Cancer Genomics Data. Cancer Discov. 2012, 2, 401–404.
- Augello, G.; Emma, M.R.; Azzolina, A.; Puleio, R.; Condorelli, L.; Cusimano, A.; Giannitrapani, L.; McCubrey, J.A.; Iovanna, J.L.; Cervello, M. The NUPR1/p73 axis contributes to sorafenib resistance in hepatocellular carcinoma. Cancer Lett. 2021, 519, 250–262.
- Zhan, Y.; Zhang, Z.; Liu, Y.; Fang, Y.; Xie, Y.; Zheng, Y.; Li, G.; Liang, L.; Ding, Y. NUPR1 contributes to radiation resistance by maintaining ROS homeostasis via AhR/CYP signal axis in hepatocellular carcinoma. BMC Med. 2022, 20, 365.
- Chen, S.-S.; Hu, W.; Wang, Z.; Lou, X.-E.; Zhou, H.-J. p8 attenuates the apoptosis induced by dihydroartemisinin in cancer cells through promoting autophagy. Cancer Biol. Ther. 2015, 16, 770–779.
- Salazar, M.; Carracedo, A.; Salanueva, Í.J.; Hernández-Tiedra, S.; Lorente, M.; Egia, A.; Vázquez, P.; Blázquez, C.; Torres, S.; García, S.; et al. Cannabinoid action induces autophagy-mediated cell death through stimulation of ER stress in human glioma cells. J. Clin. Investig. 2009, 119, 1359–1372.
- Wang, L.; Sun, J.; Yin, Y.; Sun, Y.; Ma, J.; Zhou, R.; Chang, X.; Li, D.; Yao, Z.; Tian, S.; et al. Transcriptional coregualtor NUPR1 maintains tamoxifen resistance in breast cancer cells. Cell Death Dis. 2021, 12, 149.
- Xiao, H.; Long, J.; Chen, X.; Tan, M.-D. NUPR1 promotes the proliferation and migration of breast cancer cells by activating TFE3 transcription to induce autophagy. Exp. Cell Res. 2022, 418, 113234.
- Li, Y.; Yin, Y.; Ma, J.; Sun, Y.; Zhou, R.; Cui, B.; Zhang, Y.; Yang, J.; Yan, X.; Liu, Z.; et al. Combination of AAV-mediated NUPR1 knockdown and trifluoperazine induces premature senescence in human lung adenocarcinoma A549 cells in nude mice. Oncol. Rep. 2020, 43, 681–688.
- Kong, D.K.; Georgescu, S.P.; Cano, C.; Aronovitz, M.J.; Iovanna, J.L.; Patten, R.D.; Kyriakis, J.M.; Goruppi, S. Deficiency of the Transcriptional Regulator p8 Results in Increased Autophagy and Apoptosis, and Causes Impaired Heart Function. Mol. Biol. Cell 2010, 21, 1335–1349.
- Chen, X.; Li, A.; Zhan, Q.; Jing, Z.; Chen, Y.; Chen, J. microRNA-637 promotes apoptosis and suppresses proliferation and autophagy in multiple myeloma cell lines via NUPR1. FEBS Open Bio 2021, 11, 519–528.
- Song, X.; Zhu, S.; Chen, P.; Hou, W.; Wen, Q.; Liu, J.; Xie, Y.; Liu, J.; Klionsky, D.J.; Kroemer, G.; et al. AMPK-Mediated BECN1 Phosphorylation Promotes Ferroptosis by Directly Blocking System Xc– Activity. Curr. Biol. 2018, 28, 2388–2399.e5.
- Matrood, S.; Melms, L.E.; Bartsch, D.K.; Fazio, P. Di The Expression of Autophagy-Associated Genes Represents a Valid Footprint for Aggressive Pancreatic Neuroendocrine Neoplasms. Int. J. Mol. Sci. 2023, 24, 3636.
- Rao, S.V.; Solum, G.; Niederdorfer, B.; Nørsett, K.G.; Bjørkøy, G.; Thommesen, L. Gastrin activates autophagy and increases migration and survival of gastric adenocarcinoma cells. BMC Cancer 2017, 17, 68.
- Fang, L.; Lv, J.; Xuan, Z.; Li, B.; Li, Z.; He, Z.; Li, F.; Xu, J.; Wang, S.; Xia, Y.; et al. Circular CPM promotes chemoresistance of gastric cancer via activating PRKAA2-mediated autophagy. Clin. Transl. Med. 2022, 12, e708.
- Huang, H.; Kang, R.; Wang, J.; Luo, G.; Yang, W.; Zhao, Z. Hepatitis C virus inhibits AKT-tuberous sclerosis complex (TSC), the mechanistic target of rapamycin (MTOR) pathway, through endoplasmic reticulum stress to induce autophagy. Autophagy 2013, 9, 175–195.
- Sun, W.; Yan, J.; Ma, H.; Wu, J.; Zhang, Y. Autophagy-Dependent Ferroptosis-Related Signature is Closely Associated with the Prognosis and Tumor Immune Escape of Patients with Glioma. Int. J. Gen. Med. 2022, 15, 253–270.
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550.
- Wani, A.; Rihani, S.B.A.; Sharma, A.; Weadick, B.; Govindarajan, R.; Khan, S.U.; Sharma, P.R.; Dogra, A.; Nandi, U.; Reddy, C.N.; et al. Crocetin promotes clearance of amyloid-β by inducing autophagy via the STK11/LKB1-mediated AMPK pathway. Autophagy 2021, 17, 3813–3832.
- Feng, Q.; Luo, Y.; Zhang, X.-N.; Yang, X.-F.; Hong, X.-Y.; Sun, D.-S.; Li, X.-C.; Hu, Y.; Li, X.-G.; Zhang, J.-F.; et al. MAPT/Tau accumulation represses autophagy flux by disrupting IST1-regulated ESCRT-III complex formation: A vicious cycle in Alzheimer neurodegeneration. Autophagy 2020, 16, 641–658.
- Hedna, R.; Kovacic, H.; Pagano, A.; Peyrot, V.; Robin, M.; Devred, F.; Breuzard, G. Tau Protein as Therapeutic Target for Cancer? Focus on Glioblastoma. Cancers 2022, 14, 5386.
- Chen, J.; Li, X.; Yan, S.; Li, J.; Zhou, Y.; Wu, M.; Ding, J.; Yang, J.; Yuan, Y.; Zhu, Y.; et al. An autophagy-related long non-coding RNA prognostic model and related immune research for female breast cancer. Front. Oncol. 2022, 12, 929240.
- Wu, Q.; Li, Q.; Zhu, W.; Zhang, X.; Li, H. Identification of autophagy-related long non-coding RNA prognostic signature for breast cancer. J. Cell. Mol. Med. 2021, 25, 4088–4098.
- Luo, Z.; Nong, B.; Ma, Y.; Fang, D. Autophagy related long non-coding RNA and breast cancer prognosis analysis and prognostic risk model establishment. Ann. Transl. Med. 2022, 10, 58.
- Segura, A.V.C.; Sotomayor, M.B.V.; Román, A.I.F.G.; Rojas, C.A.O.; Carrasco, A.G.M. Impact of mini-driver genes in the prognosis and tumor features of colorectal cancer samples: A novel perspective to support current biomarkers. PeerJ 2023, 11, e15410.
- Han, L.; Huang, Z.; Liu, Y.; Ye, L.; Li, D.; Yao, Z.; Wang, C.; Zhang, Y.; Yang, H.; Tan, Z.; et al. MicroRNA-106a regulates autophagy-related cell death and EMT by targeting TP53INP1 in lung cancer with bone metastasis. Cell Death Dis. 2021, 12, 1037.
- Xu, C.-G.; Yang, M.-F.; Fan, J.-X.; Wang, W. MiR-30a and miR-205 are downregulated in hypoxia and modulate radiosensitivity of prostate cancer cells by inhibiting autophagy via TP53INP1. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 1501–1508.
- Wang, W.; Liu, J.; Wu, Q. MiR-205 suppresses autophagy and enhances radiosensitivity of prostate cancer cells by targeting TP53INP1. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 92–100.
- Peuget, S.; Bonacci, T.; Soubeyran, P.; Iovanna, J.; Dusetti, N.J. Oxidative stress-induced p53 activity is enhanced by a redox-sensitive TP53INP1 SUMOylation. Cell Death Differ. 2014, 21, 1107–1118.
- Seillier, M.; Pouyet, L.; N’Guessan, P.; Nollet, M.; Capo, F.; Guillaumond, F.; Peyta, L.; Dumas, J.; Varrault, A.; Bertrand, G.; et al. Defects in mitophagy promote redox-driven metabolic syndrome in the absence of TP53 INP1. EMBO Mol. Med. 2015, 7, 802–818.
- Warde-Farley, D.; Donaldson, S.L.; Comes, O.; Zuberi, K.; Badrawi, R.; Chao, P.; Franz, M.; Grouios, C.; Kazi, F.; Lopes, C.T.; et al. The GeneMANIA prediction server: Biological network integration for gene prioritization and predicting gene function. Nucleic Acids Res. 2010, 38, W214–W220.
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128.
- Hua, R.; Cheng, D.; Coyaud, É.; Freeman, S.; Pietro, E.D.; Wang, Y.; Vissa, A.; Yip, C.M.; Fairn, G.D.; Braverman, N.; et al. VAPs and ACBD5 tether peroxisomes to the ER for peroxisome maintenance and lipid homeostasis. J. Cell Biol. 2017, 216, 367–377.
- Kalayou, S.; Hamre, A.G.; Ndossi, D.; Connolly, L.; Sørlie, M.; Ropstad, E.; Verhaegen, S. Using SILAC proteomics to investigate the effect of the mycotoxin, alternariol, in the human H295R steroidogenesis model. Cell Biol. Toxicol. 2014, 30, 361–376.
- Alizadeh, J.; Kavoosi, M.; Singh, N.; Lorzadeh, S.; Ravandi, A.; Kidane, B.; Ahmed, N.; Mraiche, F.; Mowat, M.R.; Ghavami, S. Regulation of Autophagy via Carbohydrate and Lipid Metabolism in Cancer. Cancers 2023, 15, 2195.
- Dahmene, M.; Bérard, M.; Oueslati, A. Dissecting the Molecular Pathway Involved in PLK2 Kinase-mediated α-Synuclein-selective Autophagic Degradation. J. Biol. Chem. 2017, 292, 3919–3928.
- Song, Z.; Xie, B. LncRNA OIP5-AS1 reduces α-synuclein aggregation and toxicity by targeting miR-126 to activate PLK2 in human neuroblastoma SH-SY5Y cells. Neurosci. Lett. 2021, 740, 135482.
- Zhang, J.; Ng, S.; Wang, J.; Zhou, J.; Tan, S.-H.; Yang, N.; Lin, Q.; Xia, D.; Shen, H.-M. Histone deacetylase inhibitors induce autophagy through FOXO1-dependent pathways. Autophagy 2015, 11, 629–642.
- Fan, L.; Li, B.; Li, Z.; Sun, L. Identification of Autophagy Related circRNA-miRNA-mRNA-Subtypes Network With Radiotherapy Responses and Tumor Immune Microenvironment in Non-small Cell Lung Cancer. Front. Genet. 2021, 12, 730003.
- Li, Z.-W.; Zhang, T.-Y.; Yue, G.-J.; Tian, X.; Wu, J.-Z.; Feng, G.-Y.; Wang, Y.-S. Small nucleolar RNA host gene 22 (SNHG22) promotes the progression of esophageal squamous cell carcinoma by miR-429/SESN3 axis. Ann. Transl. Med. 2020, 8, 1007.
- Sipos, F.; Barta, B.B.; Simon, Á.; Nagy, L.; Dankó, T.; Raffay, R.E.; Petővári, G.; Zsiros, V.; Wichmann, B.; Sebestyén, A.; et al. Survival of HT29 Cancer Cells Is Affected by IGF1R Inhibition via Modulation of Self-DNA-Triggered TLR9 Signaling and the Autophagy Response. Pathol. Oncol. Res. 2022, 28, 1610322.
- Sipos, F.; Kiss, A.L.; Constantinovits, M.; Tulassay, Z.; Műzes, G. Modified Genomic Self-DNA Influences In Vitro Survival of HT29 Tumor Cells via TLR9- and Autophagy Signaling. Pathol. Oncol. Res. 2019, 25, 1505–1517.
- Barta, B.B.; Simon, Á.; Nagy, L.; Dankó, T.; Raffay, R.E.; Petővári, G.; Zsiros, V.; Sebestyén, A.; Sipos, F.; Műzes, G. Survival of HT29 cancer cells is influenced by hepatocyte growth factor receptor inhibition through modulation of self-DNA-triggered TLR9-dependent autophagy response. PLoS ONE 2022, 17, e0268217.
- Li, M.; Sala, V.; Santis, M.C.D.; Cimino, J.; Cappello, P.; Pianca, N.; Bona, A.D.; Margaria, J.P.; Martini, M.; Lazzarini, E.; et al. Phosphoinositide 3-Kinase γ Inhibition Protects From Anthracycline Cardiotoxicity and Reduces Tumor Growth. Circulation 2018, 138, 696–711.
- Chen, M.-Y.; Yadav, V.K.; Chu, Y.C.; Ong, J.R.; Huang, T.-Y.; Lee, K.-F.; Lee, K.-H.; Yeh, C.-T.; Lee, W.-H. Hydroxychloroquine (HCQ) Modulates Autophagy and Oxidative DNA Damage Stress in Hepatocellular Carcinoma to Overcome Sorafenib Resistance via TLR9/SOD1/hsa-miR-30a-5p/Beclin-1 Axis. Cancers 2021, 13, 3227.
- Limagne, E.; Nuttin, L.; Thibaudin, M.; Jacquin, E.; Aucagne, R.; Bon, M.; Revy, S.; Barnestein, R.; Ballot, E.; Truntzer, C.; et al. MEK inhibition overcomes chemoimmunotherapy resistance by inducing CXCL10 in cancer cells. Cancer Cell 2022, 40, 136–152.e12.
- Anunobi, R.; Boone, B.A.; Cheh, N.; Tang, D.; Kang, R.; Loux, T.; Lotze, M.T.; Zeh, H.J. Extracellular DNA promotes colorectal tumor cell survival after cytotoxic chemotherapy. J. Surg. Res. 2018, 226, 181–191.
- Zhao, X.; Dong, Y.; Zhang, J.; Chen, C.; Gao, L.; Shi, C.; Fu, Z.; Han, M.; Tang, C.; Sun, P.; et al. Reversing immune evasion using a DNA nano-orchestrator for pancreatic cancer immunotherapy. Acta Biomater. 2023, 166, 512–523.
- Jiang, H.; Chiang, C.Y.; Chen, Z.; Nathan, S.; D’Agostino, G.; Paulo, J.A.; Song, G.; Zhu, H.; Gabelli, S.B.; Cole, P.A. Enzymatic analysis of WWP2 E3 ubiquitin ligase using protein microarrays identifies autophagy-related substrates. J. Biol. Chem. 2022, 298, 101854.
- Inokuchi, S.; Yoshizumi, T.; Toshima, T.; Itoh, S.; Yugawa, K.; Harada, N.; Mori, H.; Fukuhara, T.; Matsuura, Y.; Mori, M. Suppression of optineurin impairs the progression of hepatocellular carcinoma through regulating mitophagy. Cancer Med. 2021, 10, 1501–1514.
- Zhang, Z.; Wang, N.; Ma, Q.; Chen, Y.; Yao, L.; Zhang, L.; Li, Q.; Shi, M.; Wang, H.; Ying, Z. Somatic and germline mutations in the tumor suppressor gene PARK2 impair PINK1/Parkin-mediated mitophagy in lung cancer cells. Acta Pharmacol. Sin. 2020, 41, 93–100.
- Yan, C.; Gong, L.; Chen, L.; Xu, M.; Abou-Hamdan, H.; Tang, M.; Désaubry, L.; Song, Z. PHB2 (prohibitin 2) promotes PINK1-PRKN/Parkin-dependent mitophagy by the PARL-PGAM5-PINK1 axis. Autophagy 2020, 16, 419–434.
- Yamano, K.; Kikuchi, R.; Kojima, W.; Hayashida, R.; Koyano, F.; Kawawaki, J.; Shoda, T.; Demizu, Y.; Naito, M.; Tanaka, K.; et al. Critical role of mitochondrial ubiquitination and the OPTN–ATG9A axis in mitophagy. J. Cell Biol. 2020, 219, e201912144.
- Liu, Z.; Chen, P.; Gao, H.; Gu, Y.; Yang, J.; Peng, H.; Xu, X.; Wang, H.; Yang, M.; Liu, X.; et al. Ubiquitylation of Autophagy Receptor Optineurin by HACE1 Activates Selective Autophagy for Tumor Suppression. Cancer Cell 2014, 26, 106–120.
- Li, S.; Yang, H.; Zhao, M.; Gong, L.; Wang, Y.; Lv, Z.; Quan, Y.; Wang, Z. Demethylation of HACE1 gene promoter by propofol promotes autophagy of human A549 cells. Oncol. Lett. 2020, 20, 12143.
- Yu, Z.; Li, Y.; Han, T.; Liu, Z. Demethylation of the HACE1 gene promoter inhibits the proliferation of human liver cancer cells. Oncol. Lett. 2019, 17, 4361–4368.
- Liu, S.; van Dinther, M.; Hagenaars, S.C.; Gu, Y.; Kuipers, T.B.; Mei, H.; Gomez-Puerto, M.C.; Mesker, W.E.; ten Dijke, P. Differential optineurin expression controls TGFβ signaling and is a key determinant for metastasis of triple negative breast cancer. Int. J. Cancer 2023, 152, 2594–2606.
- Lv, D.; Yang, K.; Rich, J.N. Growth factor receptor signaling induces mitophagy through epitranscriptomic regulation. Autophagy 2023, 19, 1034–1035.
- Ali, D.M.; Ansari, S.S.; Zepp, M.; Knapp-Mohammady, M.; Berger, M.R. Optineurin downregulation induces endoplasmic reticulum stress, chaperone-mediated autophagy, and apoptosis in pancreatic cancer cells. Cell Death Discov. 2019, 5, 128.
- Hou, H.; Pan, H.; Liao, W.; Lee, C.; Yu, C. Autophagy in fibroblasts induced by cigarette smoke extract promotes invasion in lung cancer cells. Int. J. Cancer 2020, 147, 2587–2596.
- Yi, J.; Zhu, J.; Wu, J.; Thompson, C.B.; Jiang, X. Oncogenic activation of PI3K-AKT-mTOR signaling suppresses ferroptosis via SREBP-mediated lipogenesis. Proc. Natl. Acad. Sci. USA 2020, 117, 31189–31197.
- Pham, D.; Pun, N.T.; Park, P. Autophagy activation and SREBP-1 induction contribute to fatty acid metabolic reprogramming by leptin in breast cancer cells. Mol. Oncol. 2021, 15, 657–678.
- Eguchi, A.; Mizukami, S.; Nakamura, M.; Masuda, S.; Murayama, H.; Kawashima, M.; Inohana, M.; Nagahara, R.; Kobayashi, M.; Yamashita, R.; et al. Metronidazole enhances steatosis-related early-stage hepatocarcinogenesis in high fat diet-fed rats through DNA double-strand breaks and modulation of autophagy. Environ. Sci. Pollut. Res. 2022, 29, 779–789.
- Guan, M.; Fousek, K.; Chow, W.A. Nelfinavir inhibits regulated intramembrane proteolysis of sterol regulatory element binding protein-1 and activating transcription factor 6 in castration-resistant prostate cancer. FEBS J. 2012, 279, 2399–2411.
- Zhou, C.; Qian, W.; Li, J.; Ma, J.; Chen, X.; Jiang, Z.; Cheng, L.; Duan, W.; Wang, Z.; Wu, Z.; et al. High glucose microenvironment accelerates tumor growth via SREBP1-autophagy axis in pancreatic cancer. J. Exp. Clin. Cancer Res. 2019, 38, 302.
- Huang, Y.; Bell, L.N.; Okamura, J.; Kim, M.S.; Mohney, R.P.; Guerrero-Preston, R.; Ratovitski, E.A. Phospho-ΔNp63α/SREBF1 protein interactions: Bridging cell metabolism and cisplatin chemoresistance. Cell Cycle 2012, 11, 3810–3827.
- Li, S.; Oh, Y.-T.; Yue, P.; Khuri, F.R.; Sun, S.-Y. Inhibition of mTOR complex 2 induces GSK3/FBXW7-dependent degradation of sterol regulatory element-binding protein 1 (SREBP1) and suppresses lipogenesis in cancer cells. Oncogene 2016, 35, 642–650.
- Belmonte-Fernández, A.; Herrero-Ruíz, J.; Galindo-Moreno, M.; Limón-Mortés, M.C.; Mora-Santos, M.; Sáez, C.; Japón, M.Á.; Tortolero, M.; Romero, F. Cisplatin-induced cell death increases the degradation of the MRE11-RAD50-NBS1 complex through the autophagy/lysosomal pathway. Cell Death Differ. 2023, 30, 488–499.
- Gou, W.; Shen, D.; Yang, X.; Zhao, S.; Liu, Y.; Sun, H.; Su, R.; Luo, J.; Zheng, H. ING5 suppresses proliferation, apoptosis, migration and invasion, and induces autophagy and differentiation of gastric cancer cells: A good marker for carcinogenesis and subsequent progression. Oncotarget 2015, 6, 19552–19579.
- Ding, X.-Q.; Zhao, S.; Yang, L.; Zhao, X.; Zhao, G.-F.; Zhao, S.-P.; Li, Z.-J.; Zheng, H.-C. The nucleocytoplasmic translocation and up-regulation of ING5 protein in breast cancer: A potential target for gene therapy. Oncotarget 2017, 8, 81953–81966.
- Zhang, Z.; Guo, M.; Li, Y.; Shen, M.; Kong, D.; Shao, J.; Ding, H.; Tan, S.; Chen, A.; Zhang, F.; et al. RNA-binding protein ZFP36/TTP protects against ferroptosis by regulating autophagy signaling pathway in hepatic stellate cells. Autophagy 2020, 16, 1482–1505.
- Xie, C.-M.; Sun, Y. The MTORC1-mediated autophagy is regulated by the FBXW7-SHOC2-RPTOR axis. Autophagy 2019, 15, 1470–1472.
- Xie, C.-M.; Tan, M.; Lin, X.-T.; Wu, D.; Jiang, Y.; Tan, Y.; Li, H.; Ma, Y.; Xiong, X.; Sun, Y. The FBXW7-SHOC2-Raptor Axis Controls the Cross-Talks between the RAS-ERK and mTORC1 Signaling Pathways. Cell Rep. 2019, 26, 3037–3050.e4.
- Qin, S.; Wang, G.; Chen, L.; Geng, H.; Zheng, Y.; Xia, C.; Wu, S.; Yao, J.; Deng, L. Pharmacological vitamin C inhibits mTOR signaling and tumor growth by degrading Rictor and inducing HMOX1 expression. PLOS Genet. 2023, 19, e1010629.
- Lan, H.; Tan, M.; Zhang, Q.; Yang, F.; Wang, S.; Li, H.; Xiong, X.; Sun, Y. LSD1 destabilizes FBXW7 and abrogates FBXW7 functions independent of its demethylase activity. Proc. Natl. Acad. Sci. USA 2019, 116, 12311–12320.
- Feng, X.; Zou, B.; Nan, T.; Zheng, X.; Zheng, L.; Lan, J.; Chen, W.; Yu, J. MiR-25 enhances autophagy and promotes sorafenib resistance of hepatocellular carcinoma via targeting FBXW7. Int. J. Med. Sci. 2022, 19, 257–266.
- Wang, H.; Chen, J.; Zhang, S.; Zheng, X.; Xie, S.; Mao, J.; Cai, Y.; Lu, X.; Hu, L.; Shen, J.; et al. MiR-223 regulates autophagy associated with cisplatin resistance by targeting FBXW7 in human non-small cell lung cancer. Cancer Cell Int. 2020, 20, 258.
- Li, A.S.; Reuter, J.A.; Cenik, C.; Synder, M.P. Abstract 2457: Investigating the functional significance of novel, recurrent noncoding mutations of TBC1D12 in bladder cancer. Cancer Res. 2017, 77, 2457.
- Januchowski, R.; Zawierucha, P.; Ruciński, M.; Nowicki, M.; Zabel, M. Extracellular Matrix Proteins Expression Profiling in Chemoresistant Variants of the A2780 Ovarian Cancer Cell Line. Biomed Res. Int. 2014, 2014, 365867.
- Lou, W.; Ding, B.; Zhong, G.; Yao, J.; Fan, W.; Fu, P. RP11-480I12.5-004 Promotes Growth and Tumorigenesis of Breast Cancer by Relieving miR-29c-3p-Mediated AKT3 and CDK6 Degradation. Mol. Ther.-Nucleic Acids 2020, 21, 916–931.
- Mamoor, S. Differential Expression of Tubulin α 3d in Human Epithelial Ovarian Cancer; OSF: Charlottesville, VA, USA, 2021; pp. 1–10.
- Nami, B.; Wang, Z. Genetics and Expression Profile of the Tubulin Gene Superfamily in Breast Cancer Subtypes and Its Relation to Taxane Resistance. Cancers 2018, 10, 274.
More
Information
Subjects:
Biochemistry & Molecular Biology; Pathology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
582
Revisions:
2 times
(View History)
Update Date:
11 Aug 2023
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No