 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | David A. Hart | -- | 6278 | 2023-07-20 19:57:12 | | | |
| 2 | Jason Zhu | Meta information modification | 6278 | 2023-07-21 05:42:57 | | |
Video Upload Options
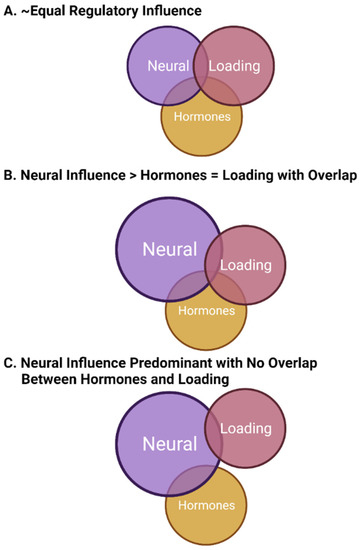
During evolution, the development of bone was critical for many species to thrive and function in the boundary conditions of Earth. Furthermore, bone also became a storehouse for calcium that could be mobilized for reproductive purposes in mammals and other species. The critical nature of bone for both function and reproductive needs during evolution in the context of the boundary conditions of Earth has led to complex regulatory mechanisms that require integration for optimization of this tissue across the lifespan. Three important regulatory variables include mechanical loading, sex hormones, and innervation/neuroregulation. The importance of mechanical loading has been the target of much research as bone appears to subscribe to the “use it or lose it” paradigm. Furthermore, because of the importance of post-menopausal osteoporosis in the risk for fractures and loss of function, this aspect of bone regulation has also focused research on sex differences in bone regulation.
1. The Regulation of Bone by Mechanical Loading
2. Neural Regulation of Bone
2.1. Background
2.2. Loss of Neural Integrity on Bone
3. Possible Role of Neural Input in Fracture Healing and Altered Healing with Brain Trauma
4. Potential Neural Influences on Development of Post-Menopausal Osteoporosis (OP) and Age-Related OP
5. Space Flight, Bedrest, and Neural Regulation
5.1. Influence of Space Flight and Bedrest on Bone Regulation
5.2. Is There a Role for Neural Regulation Dysfunction during Bone Loss during Space Flight and Osteoporosis?
References
- Hart, D.A. Learning from human responses to deconditioning environment: Improved understanding of the “use it or lose it” principle. Front. Sports Act. Living 2021, 3, 685845.
- Hart, D.A.; Zernicke, R.F.; Shrive, N.G. Homo sapiens may incorporate daily acute cycles of “conditioning-deconditioning” to maintain musculoskeletal integrity: Need to integrate with biological clocks and circadian rhythm mediators. Int. J. Mol. Sci. 2022, 23, 9949.
- Hart, D.A.; Zernicke, R.F. Optimal human functioning requires exercise across the lifespan: Mobility in a 1 g environment is intrinsic to the integrity of multiple biological systems. Front. Physiol. 2020, 11, 156.
- Frost, H.M. Bone’s mechanostat: A 2003 update. Anat. Rec. A Discov. Mol. Cell Evol. Biol. 2003, 275A, 1081–1101.
- Ferretti, J.L.; Cointry, G.R.; Capozza, R.F.; Frost, H.M. Bone mass, bone strength, muscle-bone interactions, osteopeias and osteoporosis. Mech. Aging Dev. 2003, 124, 269–279.
- Frost, H.M. A 2003 update of bone physiology and Wolff’s Law for clinicians. Angle Orthod. 2004, 74, 3–15.
- Choi, J.U.A.; Kijas, A.W.; Lauko, J.; Rowan, A.E. The mechanosensory role of osteocytes and implications for bone health and disease states. Front. Cell Dev. Biol. 2022, 9, 770143.
- Cao, W.; Helder, M.N.; Bravenboer, N.; Wu, G.; Jin, J.; Ten Bruggenkate, C.M.; Klein-Nulend, J.; Schulten, E.A.J.M. Is there a governing role of osteocytes in bone tissue regeneration? Curr. Osteopors. Rep. 2020, 18, 541–550.
- Wee, N.K.; Sims, N.A.; Morello, R. The osteocyte transcriptome: Discovering messages buried within bone. Curr. Osteoporos. Rep. 2021, 19, 604–615.
- Zeng, Y.; Riquelme, M.A.; Hua, R.; Zhang, J.; Acosta, F.M.; Gu, S.; Jiang, J.X. Mechanosensitive piezo1 calcium channel activates connexin 43 hemichannels through PI3K signaling pathway in bone. Cell Biosci. 2022, 12, 191.
- Savadipour, A.; Palmer, D.; Ely, E.V.; Collins, K.H.; Garcia-Castrorena, J.M.; Harissa, Z.; Kim, Y.S.; Oestreich, A.; Qu, F.; Rashidi, N.; et al. The role of PIEZO ion channels in the musculoskeletal system. Am. J. Physiol. Cell Physiol. 2023, 324, C728–C740.
- Wang, J.; Sun, Y.X.; Li, J. The role of mechanosensory Piezo1 in bone homeostasis and mechanobiology. Dev. Biol. 2023, 493, 80–88.
- Maycas, M.; Ardura, J.A.; de Castro, L.F.; Bravo, B.; Gortazar, A.R.; Esbit, P. Role of the parathyroid hormone type 1 receptor (PTH1R) as a mechanosensor in osteocyte survival. J. Bone Miner. Res. 2015, 30, 1231–1244.
- Maycas, M.; Esbrit, P.; Gortazar, A.R. Molecular mechanisms in bone mechanotransduction. Histol. Histopathol. 2017, 32, 751–760.
- Tirado-Cabrera, I.; Martin-Guerrero, E.; Heredero-Jimenez, S.; Ardura, J.A.; Gortazar, A.R. PTH1R translocation to primary cilia in mechanically-stimulated osteocytes prevents osteoblast formation via regulation of CXCL5 and IL-6 secretion. J. Cell Physiol. 2022, 237, 3927–3943.
- van Tol, A.F.; Schemenz, V.; Wagermaier, W.; Roschger, A.; Razi, H.; Vitienes, I.; Fratzl, P.; Willie, B.; Weinkamer, R. The mechanoresponse of bone is closely related to osteocyte lacunocanalicular network architecture. Proc. Natl. Acad. Sci. USA 2020, 117, 32251–32259.
- Yang, F.; Yu, W.; Huo, X.; Li, H.; Qi, Q.; Yang, X.; Shi, N.; Wu, X.; Chen, W. Effects of osteocyte shape on fluid flow and fluid shear stress of the loaded bone. Biomed. Res. Int. 2022, 2022, 3935803.
- Kameo, Y.; Ozasa, M.; Adachi, T. Computational framework for analyzing flow-induced strain on osteocyte as modulated by microenvironment. J. Mech. Behav. Biomed. Mater. 2022, 126, 105027.
- Wang, H.; Zheng, X.; Zhang, Y.; Huang, J.; Zhou, W.; Li, X.; Tian, H.; Wang, B.; Xing, D.; Fu, W.; et al. The endocrine role of bone: Novel functions of bone-derived cytokines. Biochem. Pharmacol. 2021, 183, 114308.
- Tasevski, V.; Sorbetti, J.M.; Chiu, S.S.; Shrive, N.G.; Hart, D.A. Influence of mechanical and biological signals on gene expression in human MG-63 cells: Evidence for a complex interplay between hydrostatic compression and vitamin D3 or TGF-beta1 on MMP-1 and MMP-3 mRNA levels. Biochem. Cell Biol. 2005, 83, 96–107.
- Bellido, T.; Delgado-Calle, J. Ex vivo organ cultures as models to study bone biology. JBMR Plus 2020, 4.
- Smyth, N.A.; Zachwieja, E.C.; Buller, L.T.; Miranda, A.D.; Steinlauf, S.D. Surgical approaches to the calcaneus and the sural nerve: There is no safe zone. Foot Ankle Surg. 2018, 24, 517–520.
- Feigl, G.C.; Schmid, M.; Zahn, P.K.; Gonzalez, C.A.A.; Litz, R.J. The posterior femoral cutaneous nerve contributes significantly to sensory innervation of the lower leg: An anatomical investigation. Br. J. Anaesth. 2020, 124, 308–313.
- Eguchi, Y.; Ohtori, S.; Yamashita, M.; Yamauchi, K.; Suzuki, M.; Orita, S.; Kamoda, H.; Arai, G.; Ishikawa, T.; Miyagi, M.; et al. Clinical applications of diffusion magnetic resonance imaging of the lumbar foraminal nerve root entrapment. Eur. Spine J. 2010, 19, 1874–1882.
- Bruns, T.M.; Wagenaar, J.B.; Bauman, M.J.; Gaunt, R.A.; Weber, D.J. Real-time control of hind limb functional electrical stimulation using feedback from dorsal root ganglia recordings. J. Neural Eng. 2013, 10, 026020.
- Savastano, L.E.; Laurito, S.R.; Fitt, M.R.; Rasmussen, J.A.; Polo, V.G.; Patterson, S.I. Sciatic nerve injury: A simple and subtle model for investigating many aspects of the nervous system damage and recovery. J. Neurosci. Methods. 2014, 227, 166–189.
- Bron, R.; Wood, R.J.; Brock, J.A.; Ivanusic, J.J. Piezo2 expression in corneal afferent neurons. J. Comp. Neurol. 2014, 522, 2967–2979.
- Nigg, B.; Enders, H. Barefoot running-some critical considerations. Footwear Sci. 2013, 5, 1–7.
- Madansingh, S.; Murphree, D.H.; Kaufman, K.R.; Fortune, E. Assessment of gait kinetics in post-menopausal women using tri-axial ankle accelerometers during barefoot walking. Gait Posture 2019, 69, 85–90.
- Brazill, J.M.; Beeve, A.T.; Craft, C.S.; Ivanusic, J.J.; Scheller, E.L. Nerves in bone: Evolving concepts in pain and anabolism. J. Bone Miner. Res. 2019, 34, 1393–1406.
- Dimitri, P.; Rosen, C. The central nervous system and bone metabolism: An evolving story. Calcif. Tissue Int. 2017, 100, 476–485.
- Xu, J.; Zhang, Z.; Zhao, J.; Meyers, C.A.; Lee, S.; Qin, Q.; James, A.W. Interaction between the nervous and skeletal systems. Front. Cell Dev. Biol. 2022, 10, 976736.
- Minoia, A.; Carbonare, L.D.; Schwamborn, J.C.; Bolognin, S.; Valenti, M.T. Bone tissue and the nervous system: What do they have in common? Cells 2022, 12, 51.
- Cooper, R.R. Nerves in cortical bone. Science 1968, 160, 127–128.
- Elefteriou, F. Impact of the autonomic nervous system on the skeleton. Physiol. Rev. 2018, 98, 1083–1112.
- Tomlinson, R.E.; Christiansen, B.A.; Giannone, A.A.; Genetos, D.C. The role of nerves in skeletal development, adaptation, and aging. Front. Endocrinol. 2020, 11, 646.
- Rajpar, I.; Tomlinson, R.E. Function of peripheral nerves in the development and healing of tendon and bone. Semin. Cell Dev. Biol. 2022, 123, 48–56.
- Abeynayake, N.; Arthur, A.; Gronthos, S. Crosstalk between skeletal and neural tissues is critical for skeletal health. Bone 2021, 142, 115645.
- Wan, Q.Q.; Qin, W.P.; Ma, Y.X.; Shen, M.J.; Li, J.; Zhang, Z.B.; Chen, J.H.; Tay, F.R.; Niu, L.N.; Jiao, K. Crosstalk between bone and nerves within bone. Adv. Sci. 2021, 8, 2003390.
- Liu, S.; Chen, T.; Wang, R.; Huang, H.; Fu, S.; Zhao, Y.; Wang, S.; Wan, L. Exploring the effect of the “quaternary regulation” theory of “peripheral nerve-angiogenesis-osteoclast-osteogenesis” on osteoporosis based on neuropeptides. Front. Endocrinol. 2022, 13, 908043.
- Bjurholm, A.; Kreicbergs, A.; Brodin, E.; Schulteberg, M. Substance P- and CGRP-immunoreactive nerves in bone. Peptides 1988, 9, 165–171.
- Hu, B.; Lv, X.; Wei, L.; Wang, Y.; Zheng, G.; Yang, C.; Zang, F.; Wang, J.; Li, J.; Wu, X.; et al. Sensory nerve maintains intervertebral disc extracellular matrix homeostasis via CGRP/CHSY1 axis. Adv. Sci. 2022, 9, e2202620.
- Xu, J.; Wang, J.; Chen, X.; Li, Y.; Mi, J.; Qin, L. The effects of calcitonin gene-related peptide on bone homeostasis and regeneration. Curr. Osteoporos. Rep. 2020, 18, 621–632.
- Jones, K.B.; Mollano, A.V.; Morcuende, J.A.; Cooper, R.R.; Saltzman, C.L. Bone and brain: A review of neural, hormonal, and musculoskeletal connections. Iowa Orthop. J. 2004, 24, 123–132.
- Wang, L.; Hou, S.; Sabsovich, I.; Guo, T.Z.; Wei, T.; Kingery, W.S. Mice lacking substance P have normal bone modeling but diminished bone formation, increased resorption, and accelerated osteopenia with aging. Bone 2021, 144, 115806.
- Li, F.X.; Xu, F.; Lin, X.; Wu, F.; Zhong, J.W.; Wang, Y.; Guo, B.; Zheng, M.H.; Chan, S.K.; Yuan, L.Q. The role of substance P in the regulation of bone and cartilage metabolic activity. Front. Endocrinol. 2020, 11, 77.
- Chen, Q.C.; Zhang, Y. The role of NPY in the regulation of bone metabolism. Front. Endocrinol. 2022, 13, 833485.
- Irie, K.; Hara-Irie, F.; Ozawa, H.; Yajima, T. Calcitonin gene-related peptide (CGRP)-containing nerve fibers in bone tissue and their involvement in bone remodeling. Microsc. Res. Tech. 2002, 58, 85–90.
- Wu, H.; Lin, X.Q.; Long, Y.; Wang, J. Calcitonin gene-related peptide is potential therapeutic target of osteoporosis. Heliyon 2022, 8, 12288.
- Wee, N.K.Y.; Novak, S.; Ghosh, D.; Root, S.H.; Dickerson, I.M.; Kalajzic, I. Inhibition of CGRP signaling impairs fracture healing in mice. J. Orthop. Res. 2023, 41, 1228–1239.
- Qiao, Y.; Wang, Y.; Zhou, Y.; Jiang, F.; Huang, T.; Chen, L.; Lan, J.; Yang, C.; Guo, Y.; Yan, S.; et al. The role of nervous system in adaptive response of bone to mechanical loading. J. Cell Physiol. 2019, 234, 7771–7780.
- Corr, A.; Smith, J.; Baldock, P. Neuronal control of bone remodeling. Toxicol. Pathol. 2017, 45, 894–903.
- Gajda, M.; Adriaensen, D.; Cichocki, T. Development of the innervation of long bones: Expression of the growth-associated protein. Folia Histochem. Cytobiol. 2000, 38, 103–110.
- Chartier, S.R.; Mitchell, S.A.T.; Majuta, L.A.; Mantyh, P.W. The changing sensory and sympathetic innervation of the young, adult, and aging mouse femur. Neuroscience 2018, 387, 178–190.
- Valdez, G. Effects of disease-afflicted and aging neurons on the musculoskeletal system. Bone 2019, 122, 31–37.
- Salo, P.T.; Tatton, W.G. Age-related loss of knee joint afferents in mice. J. Neurosci. Res. 1993, 35, 664–677.
- Salo, P.T.; Theriault, E. Number, distribution and neuropeptide content of rat knee joint afferents. J. Anat. 1997, 190 Pt 4, 515–522.
- Brighton, C.T.; Taddurin, G.T.; Goll, S.R.; Pollack, S.R. Treatment of denervation/disuse osteoporosis in the rat with a capacitively coupled electrical signal: Effects on bone formation and bone resorption. J. Orthop. Res. 1988, 6, 676–684.
- Brouwers, J.E.M.; Lambers, F.M.; van Riethergen, B.; Ito, K.; Huiskes, R. Comparison of bone loss induced by ovariectomy and neurectomy in rats analyzed by in vivo micro-CT. J. Orthop. Res. 2009, 27, 1521–1527.
- Ma, X.; Lv, J.; Sun, X.; Ma, J.; Xing, G.; Wang, Y.; Sun, L.; Wang, J.; Li, F.; Li, Y.; et al. Naringin ameliorates bone loss induced by sciatic neurectomy and increases semaphoring #A expression in denervated bone. Sci. Rep. 2015, 6, 24562.
- Li, Y.; Jie, L.; Tian, A.Y.; Zhong, S.; Tian, M.Y.; Zhong, Y.; Wang, Y.; Li, H.; Li, J.; Sun, X.; et al. Transforming growth factor beta is regulated by a glucocorticoid-dependent mechanism in denervation mouse bone. Sci. Rep. 2017, 7, 9925.
- Lin, C.C.; Chang, Y.T.; Lin, R.W.; Chang, C.W.; Wang, G.J.; Lai, K.A. Single pulsed electromagnetic field restores bone mass and microarchitecture in denervation/disuse osteopenic mice. Med. Eng. Phys. 2020, 80, 52–59.
- Bayram, P.; Karamese, S.A.; Ozdemir, B.; Salum, C.; Erol, H.S.; Karamese, M. Two flavonoids, baicalein and naringin, are effective as anti-inflammatory and anti-ocidant agents in a rat model of polymicrobial sepsis. Immunopharamacol. Immunotoxicol. 2023, 1–10.
- Brighton, C.T.; Katz, M.J.; Goll, S.R.; Nichols, C.E.; Pollack, S.R. Prevention and treatment of sciatic denervation disuse osteoporosis in the rat tibia with capacitively coupled electrical stimulation. Bone 1985, 6, 87–97.
- Tamaki, H.; Yotani, K.; Ogita, F.; Hayao, K.; Kirimto, H.; Onishi, H.; Kusuga, N.; Yamamoto, N. Low-frequency electrical stimulation of denervated skeletal muscle retards muscle and trabecular bone loss in aged rats. Int. J. Mol. Sci. 2019, 16, 822–830.
- Kawao, N.; Moritake, A.; Tatsumi, K.; Kaji, H. Roles of irisin in the linkage from muscle to bone during mechanical unloading in mice. Calcif. Tissue Int. 2018, 103, 24–34.
- Deng, J.; Cohen, D.J.; Redden, J.; McClure, M.J.; Boyan, B.D.; Schwartz, Z. Differential effects of nerectomy and botox-induced muscle paralysis on bone phenotype and titanium implant osseointegration. Bone 2021, 153, 116145.
- Qin, W.; Bauman, W.A.; Cardozo, C.P. Evolving concepts in neurogenic osteoporosis. Curr. Osteoporos. Rep. 2010, 8, 212–218.
- Morse, L.R.; Biering-Soerensen, F.; Carbone, L.D.; Cervinka, T.; Cirnigliaro, C.M.; Johnston, T.E.; Liu, N.; Troy, K.L.; Weaver, F.M.; Shuhart, C.; et al. Bone mineral density testing in spinal cord injury: 2019 ISCD official position. J. Clin. Densitom. 2019, 22, 554–566.
- Anderson, D.; Park, A.J. Prophylactic treatment of osteoporosis after SCI: Promising research, but not yet indicated. Spinal Cord Ser. Cases 2019, 5, 25.
- Invernizzi, M.; de Sire, A.; Reno, F.; Cisari, C.; Runza, L.; Baricich, A.; Carda, S.; Fusco, N. Spinal cord injury as a model of bone-muscle interactions: Therapeutic implications from in vitro and in vivo studies. Front. Endocrinol. 2020, 11, 204.
- Carda, S.; Cisari, C.; Invernizzi, M.; Bevilacqua, M. Osteoporosis after stroke: A review of the causes and potential treatments. Cerebrovas. Dis. 2009, 28, 191–200.
- Yang, F.Z.; Jehu, D.A.M.; Ouyang, H.; Lam, F.M.H.; Pang, M.Y.C. The impact of stroke on bone properties and muscl-bone relationship: A systematic review and meta-analysis. Osteoporo. Int. 2020, 31, 211–224.
- Sato, Y. Abnormal bone and calcium metabolism in patients after stroke. Arch. Phys. Med. Rehabil. 2000, 81, 117–121.
- Pluskiewicz, W. Skeletal consequences in patients after stroke. Endokrynol. Pol. 2011, 62, 48–50.
- Hsieh, C.Y.; Sung, S.F.; Huang, H.K. Drug treatment strategies for osteoporosis in stroke patients. Expert Opin. Pharmacother. 2020, 21, 811–821.
- Borschmann, K. Exercise protects bone after stroke, or does it? A narrative review of the evidence. Stroke Res. Treat. 2012, 2012, 103697.
- Borshmann, K.; Pang, M.Y.C.; Bernhardt, J.; Juliano-Burns, S. Strepping towards prevention of bone loss after stroke: A systematic review of the skeletal effects of physical activity after stroke. Int. J. Stroke 2012, 7, 330–335.
- Sallehuddin, H.; Ong, T.; Said, S.M.; Tarmizi, N.A.A.; Loh, S.P.; Lim, W.C.; Nedarajah, R.; Lim, H.T.; Zambri, N.H.M.; Ho, Y.Y.; et al. Non-pharmacological interventions for bone health after stroke: A systematic review. PLoS ONE 2022, 17, e0263935.
- Lichy, A.M.; Groah, S. Asymmetric lower-limb bone loss after spinal cord injury: Case report. J. Rehabil. Res. Dev. 2012, 49, 221–226.
- Bauman, W.A.; Cardozo, C.P. Osteoporosis in individuals with spinal cord injury. PMR 2015, 7, 188–201.
- Shams, R.; Drasites, K.P.; Zaman, V.; Matzelle, D.; Shields, D.C.; Sole, C.J.; Haque, A.; Banik, N.L. The pathophysiology of osteoporosis after spinal cord injury. Int. J. Mol. Sci. 2021, 22, 3057.
- Abdelrahman, S.; Ireland, A.; Winter, E.M.; Purcell, M.; Coupaud, S. Osteoporosis after spinal cord injury: Aetiology, effects and therapeutic approaches. J. Musculoskelet. Neuronal Interact. 2021, 21, 26–50.
- Antoniou, G.; Benetos, I.S.; Vlamis, J.; Pneumaticos, S.G. Bone mineral density post a spinal cord injury: A review of the current literature guidelines. Cureus 2022, 14, e23434.
- Bauman, W.A. Pharmacological approaches for bone health in persons with spinal cord injury. Curr. Opin. Pharmacol. 2021, 60, 346–359.
- Edwards, W.B.; Schnitzer, T.J.; Troy, K.L. Bone mineral and stiffness loss at the distal femur and proximal tibia in acute spinal cord injury. Osteoporos. Int. 2014, 25, 1005–1015.
- Lobos, S.; Cooke, A.; Simonett, G.; Ho, C.; Boyd, S.K.; Edwards, W.B. Trabecular bone score at the distal femur and proximal tibia in individuals with spinal cord injury. J. Clin. Densitom. 2019, 22, 249–256.
- Haider, I.T.; Simonian, N.; Saini, A.S.; Leung, F.M.; Edwards, W.B.; Schnitzer, T.J. Open-label clinical trial of alendronate after teriparatide therapy in people with spinal cord injury and low bone mineral density. Spinal Cord 2019, 57, 832–842.
- Weaver, F.M.; Le, B.; Ray, C.; Miskevics, S.; Gonzalez, B.; Carbone, L.D. Predicting osteoporosis medication receipt in veterans with a spinal cord injury: A retrospective cohort study. J. Spinal Cord Med. 2019, 42, 760–767.
- Sutor, T.W.; Kura, J.; Mattingly, A.J.; Otzel, D.M.; Yarrow, J.F. The effects of exercise and activity-based physical therapy on bone after spinal cord injury. Int. J. Mol. Sci. 2022, 23, 608.
- Castello, F.; Louis, B.; Cheng, J.F.; Armento, M.; Santos, A.M. The use of functional electrical stimulation cycles in children and adolescents with spinal cord dysfunction: A pilot study. J. Pediatr. Rehabil. Med. 2012, 5, 261–273.
- Hart, D.A. Evidence for a potential “knee-eye-brain axis” involved in mobility and navigation control: Knee injury and obesity may disrupt axis integrity. J. Biomed. Sci. Eng. 2018, 11, 37–44.
- Jiang, S.D.; Jiang, L.S.; Dai, L.Y. Mechanisms of osteoporosis in spinal cord injury. Clin. Endocrinol. 2006, 65, 555–565.
- Lombardi, G.; Mondaini, N.; Macchiarella, A.; Del Popolo, G. Female sexual dysfunction and hormonal status in spinal cord injured (SCI) patients. J. Androl. 2007, 28, 722–726.
- Charls, A.C.; Rawat, N.; Zachariah, K. Menstrual cycle changes after spinal cord injury. Spinal Cord 2022, 60, 712–715.
- Gass, M.L.; Kagan, R.; Kohles, J.D.; Martens, M.S. Bone turnover marker profile in relation to the menstrual cycle of premenopausal healthy women. Menopause 2008, 15, 667.
- Sabour, H.; Javidan, A.N.; Latifi, S.; Larijani, B.; Shidfar, F.; Vafa, M.R.; Heshmat, R.; Razavi, H.E. Bone markers in patients with chronic traumatic spinal cord injury. Spine J. 2014, 14, 1132–1138.
- Edwards, W.B.; Schnitzer, T.J. Bone imaging and fracture risk after spinal cord injury. Curr. Osteoporos. Rep. 2015, 13, 310–317.
- Schulte, L.M.; Scully, R.D.; Kappa, J.E. Management of lower extremity long-bone fractures in spinal cord injury patients. J. Am. Acad. Orthop. Surg. 2017, 25, e204–e213.
- Haider, I.T.; Lobos, S.M.; Simonian, N.; Schnitzer, T.J.; Edwards, W.B. Bone fragility after spinal cord injury: Reductions in stiffness and bone mineral at the distal femur and proximal tibia as a function of time. Osteoporos. Int. 2018, 29, 2703–2715.
- Zleik, N.; Weaver, F.; Harmon, R.I.; Le, B.; Radhakrishnan, R.; Jirau-Rosaly, W.D.; Craven, B.C.; Raiford, M.; Hill, J.N.; Etingen, B.; et al. Prevention and management of ostepporosis and osteoporotic fractures in persons with a spinal cord injury or disorder: A systematic scoping review. J. Spinal Cord Med. 2019, 42, 735–759.
- Wang, L.; Yao, X.; Tang, X.; Ding, H.; Zhang, H.; Yuan, J. The effects of spinal cord injury on bone healing in patients with femoral fractures. J. Spinal Cord Med. 2014, 37, 414–419.
- Grassner, L.; Klein, B.; Maier, D.; Buhren, V.; Vogel, M. Lower extremity fractures in patients with spinal cord injury characteristics, outcome and risk factors for non-unions. J. Spinal Cord Metab. 2018, 41, 676–683.
- Wang, L.; Liu, L.; Pan, Z.; Zeng, Y. Serum leptin, bone mineral density and the healing of long bone fractures in men with spinal cord injury. Bosn J. Basic Med. Sci. 2015, 15, 69–74.
- Aro, H.; Eerola, E.; Aho, A.J.; Penttinen, R. Healing of experimental fractures in the denervated limbs of rats. Clin. Orthop. Relat. Res. 1981, 155, 211–217.
- Nordsletten, L.; Madsen, J.E.; Almaas, R.; Rootwelt, T.; Halse, J.; Konttinen, Y.T.; Hukkanen, M.; Santavirta, S. The neuronal regulation of fracture healing. Effects of sciatic nerve resection in rat tibia. Acta Orthop. Scand. 1994, 65, 299–304.
- Ding, W.-G.; Jiang, S.-D.; Zhang, Y.-H.; Dai, L.-Y. Bone loss and impaired fracture healing in spinal cord injured mice. Osteoporo. Int. 2011, 22, 507–515.
- Zheng, X.Q.; Huang, J.; Lin, J.L.; Song, C.L. Pathological mechanism of acute bone loss after fracture. J. Adv. Res. 2022, 49, 63–80.
- Benassy, J. Ossifications and fracture-healing in paraplegia and brain injuries. Proc. Annu. Clin. Spinal Cord Inj. Conf. 1966, 15, 55–60.
- Garland, D.E. Clinical observations on fractures and heterotopic ossification in the spinal cord and traumatic brain injured populations. Clin. Orthop. Relat. Res. 1988, 233, 86–101.
- Kushwaha, V.P.; Garland, D.G. Extremity fractures in the patient with a traumatic brain injury. J. Am. Acad. Orthop. Surg. 1998, 6, 298–307.
- Morley, J.; Marsh, S.; Drakoulakis, E.; Pape, H.-C.; Giannoudis, P.Y. Does traumatic brain injury result in accelerated fracture healing? Injury 2005, 36, 363–368.
- Morioka, K.; Marmor, Y.; Sacramento, J.A.; Lin, A.; Shao, T.; Miclau, K.; Clark, D.R.; Beattie, M.S.; Marcucio, R.S.; Miclau, T., 3rd; et al. Differential fracture response to traumatic brain injury suggests dominance of neuroinflammatory response in polytrauma. Sci. Rep. 2019, 9, 12199.
- Mollahosseimni, M.; Ahmadirad, H.; Goujani, R.; Khorramdelazad, H. The association between traumatic brain injury and accelerated fracture healing: A study on the effects of growth factors and cytokines. J. Mol. Neurosci. 2021, 8, 162–168.
- Ravi, P.; Nagesaran, J.; Ramanujam, M.; Suriyakumar, S.; Nambi, E.A. Correlation between traumatic brain injuries and callus formation in long bone fractures. Indian J. Orthop. 2022, 56, 837–846.
- Haffner-Luntzer, M.; Wieber, B.; Morioka, K.; Lackner, I.; Fischer, V.; Bahney, C.; Ignatius, A.; Kalbitz, M.; Marcucio, R.; Miclau, T. Altered early immune response after fracture and traumatic brain injury. Front. Immunol. 2023, 14, 1074207.
- Hofman, M.; Koopmans, G.; Kobbe, P.; Poeze, M.; Andruszkow, H.; Brink, P.R.; Pape, H.-C. Improved fracture healing in patients with concomitant brain injury: Proven or not? Mediat. Inflamm. 2015, 2015, 204842.
- Shim, D.W.; Hong, H.; Cho, K.-C.; Kim, S.H.; Lee, J.W.; Sung, S.-Y. Accelerated tibia fracture healing in traumatic brain injury in accordance with increased hematoma formation. BMC Musculoskelet. Discord. 2022, 23, 1110.
- Wildburger, R.; Zarkovic, N.; Tonkovic, G.; Skoric, T.; Frech, S.; Hartleb, M.; Loncaric, I.; Zakovic, K. Post-traumatic hormonal disturbances: Prolactin as a link between head injury and ehanced osteogenesis. J. Endocrinol. Investig. 1998, 21, 78–86.
- Wildburger, R.; Zarkovic, N.; Egger, G.; Petek, W.; Zarkovic, K.; Hofer, H.P. Basic fibroblast growth factor (BFGF) immunoreactivity as a possible link between head injury and impaired bone fracture healing. Bone Miner. 1994, 27, 183–192.
- Wildburger, R.; Zarkovic, N.; Egger, G.; Petek, W.; Meinitzer, A.; Borovic, S.; Zarkovic, K.; Li, L.; Stipancic, I.; Trbojevic-Cepe, M.; et al. Comparison of the values of basic fibroblast growth factor determined by an immunoassay in the sera of patients with traumatic brain injury and enhanced osteogenesis and the effects of the same sera on the fibroblast growth in vitro. Eur. J. Clin. Chem. Clin. Biochem. 1995, 33, 693–698.
- Song, Y.; Bi, L.; Zhang, Z.; Huang, Z.; Hou, W.; Lu, X.; Sun, P.; Han, Y. Increased levels of calcitonin gene-related peptide in serum accelerate fracture healing following traumatic brain injury. Mol. Med. Rep. 2012, 5, 432–438.
- Zhang, R.; Liang, Y.; Wei, S. The expressions of NGF and VEGF in the fracture tissues are closely associated with accelerated clavicle fracture healing in patients with traumatic brain injury. Ther. Clin. Risk Manag. 2018, 14, 2315–2322.
- Khare, G.N.; Gautam, V.K.; Gupta, L.N.; Gupta, A.K. A new hypothesis for faster healing of fractures in head injured patients. Indian J. Med. Sci. 1995, 49, 281–284.
- Cadosch, D.; Gautschi, O.P.; Thyer, M.; Song, S.; Skirving, A.P.; Filgueira, L.; Zellweger, R. Humoral factors enhance fracture-healing and callus formation in patients with traumatic brain injury. J. Bone Joint Surg. Am. 2009, 91, 282–288.
- Xu, Y.-Q.; Qin, M.-L.; Feng, S.-Y.; Huang, Y.; Jia, Z. Expressions and significance of calcitonin gene-related peptide and nerve growth factor in rabbit model of traumatic brain injury complicated with tibial fracture: Preliminary results. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 5040–5050.
- Wei, Y.; Wang, L.; Clark, J.C.M.; Dass, C.R.; Choong, P.F.M. Elevated leptin expression in a rat model of fracture and traumatic brain injury. J. Pharm. Pharmacol. 2008, 60, 1667–1672.
- Boes, M.; Kain, M.; Kakar, S.; Nicholls, F.; Cullinane, D.; Gerstenfeld, L.; Einhorn, T.A.; Tornetta, P., 3rd. Osteogenic effects of traumatic brain injury on experimental fracture-healing. J. Bone Joint Surg. Am. 2006, 88, 738–743.
- Arik, M.; Ekinci, Y.; Gurbuz, K.; Batin, S. The effects of focal brain damage on fracture healing: An experimental study. Eklem Hastalik. Cerrahisi. 2019, 30, 267–274.
- Yang, C.; Gao, C.; Liu, N.; Zhu, Y.; Zhu, X.; Su, X.; Zhang, Q.; Wu, Y.; Zhang, C.; Liu, A.; et al. The effect of traumatic brain injury on bone healing from a novel exosome centered perspective in a mice model. J. Orthop. Translat. 2021, 30, 70–81.
- Kesavan, C.; Rundle, C.; Mohan, S. Repeated mild traumatic brain injury impairs fracture healing in male mice. BMC Res. Notes 2022, 15, 25.
- Meyer, M.H.; Etienne, W.; Meyer, R.A., Jr. Altered mRNA expression of genes related to nerve cell activity in the fracture callus of older rats: A randomized, controlled, microarray study. BMC Musculoskelet. Disord. 2004, 5, 24.
- Wang, L.; Tang, X.; Zhang, H.; Yuan, J.; Ding, H.; Wei, Y. Elevated leptin expression in rat model of traumatic spinal cord injury and femoral fracture. J. Spinal Cord Med. 2011, 34, 501–509.
- Yan, H.; Zhang, H.-W.; Fu, P.; Liu, B.-L.; Jin, W.-Z.; Duan, S.-B.; Xue, J.; Liu, K.; Sun, Z.-M.; Zeng, X.-W. Leptin’s effect on accelerated fracture healing after traumatic brain injury. Neurol. Res. 2013, 35, 537–544.
- Garbe, A.; Graef, F.; Appelt, J.; Schmidt-Bleek, K.; Jahn, D.; Lunnemann, T.; Tsitsilonis, S.; Seemann, R. Leptin mediated pathways stabilize posttraumatic insulin and osteocalcin patterns after long bone fracture and concomitant traumatic brain injury and thus influence fracture healing in a combined murine trauma model. Int. J. Mol. Sci. 2020, 21, 9144.
- Graef, F.; Seemann, R.; Garbe, A.; Schmidt-Bleek, K.; Schaser, K.D.; Keller, J.; Duda, G.; Tsitisilonis, S. Impaired fracture healing with high non-union rates remain irreversible after traumatic brain injury in leptin-deficient mice. J. Musculoskelet. Neuronal Interact. 2017, 17, 78–85.
- Seemann, R.; Graef, F.; Garbe, A.; Keller, J.; Huang, F.; Duda, G.; Schmidt-Bleek, K.; Schaser, K.-D.; Tsitsilonis, S. Leptin-deficiency eradicates the positive effect of traumatic brain injury on bone healing: Histological analyses in a combined trauma mouse model. J. Musculoskelet. Neuronal Interact. 2018, 18, 32–41.
- Khallaf, F.G.; Kehinde, E.O.; Hussein, S. Bone healing and hormonal bioassay in patients with long-bone fractures and concomitant head injury. Med. Princ. Pract. 2016, 25, 336–342.
- Imayama, I.; Prasad, B. Role of leptin in obstructive sleep apnea. Ann. Am. Thorac. Soc. 2017, 14, 1607–1621.
- Moore, T.J. Functional outcome following surgical excision of heterotopic ossification in patients with traumatic brain injury. J. Orthop. Trauma. 1993, 7, 11–14.
- Zhao, X.-G.; Zhao, G.-F.; Ma, Y.-F.; Jiang, G.-Y. Research progress in mechanism of traumatic brain injury affecting speed of fracture healing. Clin. J. Traumatol. 2007, 10, 376–380.
- Huang, H.; Cheng, W.-X.; Hu, Y.-P.; Chen, J.-H.; Zheng, Z.-T.; Zhang, P. Relationship between heterotopic ossification and traumatic brain injury: Why severe traumatic brain injury increases risk of heterotopic ossification. J. Orthop. Translat. 2017, 12, 16–25.
- Anthonissen, J.; Steffen, C.T.; Hofmann, A.; Victor, J. The pathogenesis of heterotopic ossification after traumatic brain injury: A review of current literature. Acta Orthop. Belg. 2020, 86, 369–377.
- O’Brien, E.J.O.; Frank, C.B.; Shrive, N.G.; Halgrimsson, B.; Hart, D.A. Heterotopic mineralization (ossification or calcification) in tendinopathy or following surgical tendon trauma. Int. J. Exp. Pathol. 2012, 93, 319–331.
- O’Brien, E.J.O.; Shrive, N.G.; Rosvold, J.M.; Thornton, G.M.; Frank, C.B.; Hart, D.a. Tendon mineralization is accelerated bilaterally and creep of contralateral tendons is increased after unilateral needle injury of murine Achilles tendon. J. Orthop. Res. 2013, 31, 1520–1528.
- Salles, J.P. Bone metabolism during pregnancy. Ann. Endocrinol. 2016, 77, 163–168.
- Liu, X.S.; Wang, L.; de Bakker, C.M.J.; Lai, X. Mechanical regulation of the maternal skeleton during reproduction and lactation. Curr. Osteoporos. Rep. 2019, 17, 375–386.
- Kovacs, C.S. The skeleton is a storehouse of mineral that is plundered during lactation and (fully?) replenished afterwards. J. Bone Miner. Res. 2017, 32, 676–680.
- Maliha, G.; Morgan, J.; Vranhas, M. Transient osteoporosis of pregnancy. Injury 2012, 43, 1237–1241.
- Cohen, A.; Kamanda-Kosseh, M.; Dempster, D.W.; Zhou, H.; Muller, R.; Goff, E.; Colon, I.; Bucovsky, M.; Stubby, J.; Nickolas, T.L.; et al. Women with pregnancy and lactation-associated osteoporosis (PLO) have low bone remodeling rates at the tissue level. J. Bone Miner. Res. 2019, 34, 1552–1561.
- Hart, D.A. Sex differences in biological systems and the conundrum of menopause: Potential commonalities in post-menopausal disease mechanisms. Int. J. Mol. Sci. 2022, 23, 4119.
- Zhang, W.; Liu, Y.; Xu, J.; Fan, C.; Zhang, B.; Feng, P.; Wang, Y.; Kong, Q. The role of sympathetic nerves in osteoporosis: A narrative review. Biomedicines 2022, 11, 33.
- Schumacher, M.; Sitruk-Ware, R.; De Nicola, A.F. Progesterone and progestins: Neuroprotection and myelin repair. Curr. Opin. Pharmacol. 2008, 8, 740–746.
- Nugent, B.M.; Tobet, S.A.; Lara, H.E.; Lucion, A.B.; Wilson, M.E.; Recabarren, S.E.; Paredes, A.H. Hormonal programing across the lifespan. Horm. Metab. Res. 2012, 44, 577–586.
- Baker, S.E.; Limberg, J.K.; Ranadive, S.M.; Joyner, M.J. Neurovascular control of blood pressure is influenced by aging, sex, and sex hormones. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 311, R1271–R1275.
- Clarkoudian, N.; Hart, E.C.; Barnes, J.N.; Joyner, M.J. Autonomic control of body temperature and blood pressure: Influences of female sex hormones. Clin. Auton. Res. 2017, 27, 149–155.
- Ghoumari, A.M.; Ghanem, C.A.; Asbelaoui, N.; Schaumaker, M.; Hussain, R. Roles of progesterone, testosterone and their nuclear receptors in central nervous system myelination and remyelination. Int. J. Mol. Sci. 2020, 21, 3163.
- Littlejohn, E.L.; Fedorchak, S.; Boychuk, C.R. Sex-steroid-dependent plasticity of brain-stem autonomic circuits. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2020, 319, R60–R68.
- Rosato, E.; Sciarra, F.; Anastasiadou, E.; Lenzi, A.; Venneri, M.A. Revisiting the physiological role of androgens in women. Expert Rev. Endocrinol. Metab. 2022, 17, 547–561.
- Ansdell, P.; Brownstein, C.G.; Skarabot, J.; Hicks, K.M.; Simones, D.C.M.; Thomas, K.; Howatson, G.; Hunter, S.K.; Goodall, S. Menstrual cycle-associated modulations in neuromuscular function and fatigability of the knee extensors in eumenorrheic women. J. Appl. Physiol. 2019, 126, 701–712.
- Hwang, C.L.; Okazaki, K.; Shibata, S.; Liu, Y.L.; Fu, Q. Menstrual cycle effects on sympathetic neural burst amplitude distribution during orthostasis in young women. Clin. Auton. Res. 2021, 31, 767–773.
- Pellegrino, A.; Ticlus, P.M.; Vandenboom, R. Mechanisms of estrogen influence on skeletal muscle: Mass, regeneration, and mitochondrial function. Sports Med. 2022, 52, 2853–2869.
- Oosthuyse, T.; Strauss, J.A.; Hackney, A.C. Understanding the female athlete: Molecular mechanisms underpinning menstrual phase differences in exercise metabolism. Eur. J. Appl. Physiol. 2023, 123, 423–450.
- Bilke, D.D. The free hormone hypothesis: When, why, and how to measure the free hormone levels to assess vitamin D, thyroid, sex hormone, and cortisol status. JBMR Plus 2020, 5, e10418.
- Cheng, C.H.; Chen, L.R.; Chen, K.H. Osteoporosis due to hormonal imbalance: An overview of the effects of estrogen deficiency and glucocorticoid overuse on bone turnover. Int. J. Mol. Sci. 2022, 23, 1376.
- Albright, F.; Smith, P.H.; Richardson, A.M. Postmenopausal osteoporosis. JAMA 1941, 116, 2465–2474.
- Forbes, A.P. Fuller Albright. His concept of postmenopausal osteoporosis and what came of it. Clin. Orthop. Relat. Res. 1991, 269, 128–141.
- Hart, D.A. Are secondary effects of bisphosphonates on the vascular system of bone contributing to increased risk for atypical femoral fractures in osteoporosis? BioEssays 2023, 45, 2200206.
- Seeman, E. During aging, men lose less bone than women because they gain more periosteal bone, not because they resorb less endosteal bone. Calif. Tissue Int. 2001, 69, 205–208.
- Seeman, E. Periosteal bone formation—A neglected determinant of bone strength. N. Engl. J. Med. 2003, 349, 320–323.
- Seeman, E. The periosteum- a surface for all seasons. Osteoporo. Int. 2007, 18, 123–128.
- Seeman, E. Structural basis of growth-related gain and age-related loss of bone strength. Rheumatology 2008, 47 (Suppl. S4), iv2–iv8.
- Courties, A.; Sellam, J.; Berenbaum, F. Role of the autonomic nervous system in osteoarthritis. Best Pract. Res. Clin. Rheumatol. 2017, 31, 661–675.
- Plack, C.J.; Barker, D.; Predergast, G. Perceptual consequences of “hidden” hearing loss. Trends Hear. 2014, 18, 2331216514550621.
- Pollock, N.; Staunton, C.A.; Vasilaki, A.; McArdle, A.; Jackson, M.J. Denervated muscle fibers induce mitochondrial peroxide generation in neighboring innervated fibers: Role in muscle aging. Free Radic. Biol. Med. 2017, 112, 84–92.
- Maryanovich, M.; Zahalka, A.H.; Pierce, H.; Pinho, S.; Nakahara, F.; Asada, N.; Wei, Q.; Wang, X.; Ciero, P.; Xu, J.; et al. Adrenergic nerve degeneration in bone marrow drives aging of the hematopoietic stem cell niche. Nat. Med. 2018, 24, 782–791.
- Vincent, K.; Mohanty, S.; Pinelli, R.; Bonavita, R.; Pricop, P.; Albert, T.J.; Dahia, C.L. Aging of mouse intervertebral disc and association with back pain. Bone 2019, 123, 246–259.
- Fogarty, M.J.; Porras, M.A.G.; Mantilla, C.B.; Sieck, G.C. Diaphragm neuromuscular transmission failure in aged rats. J. Neurophysiol. 2019, 122, 93–104.
- Blaszkiewicz, M.; Willows, J.W.; Dubois, A.L.; Waible, S.; DiBello, K.; Lyons, L.L.; Johnson, C.P.; Paradie, E.; Banks, N.; Motyl, K.; et al. Neuropathy and neural plasticity in the subcutaneous white adipose depot. PLoS ONE 2019, 14, e0221766.
- Kos, O.; Hughson, R.L.; Hart, D.A.; Clement, G.; Frings-Meuthen, P.; Linnarsson, D.; Paloski, W.H.; Rittweger, J.; Wuyts, F.; Zange, J.; et al. Elevated serum soluble CD200 and CD200R as surrogate markers of bone loss under bed rest conditions. Bone 2014, 60, 33–40.
- Hart, D.A. Homo sapiens-A species not designed for space flight: Health risks in low Earth orbit and beyond, including potential risks when traveling beyond the geomagnetic field of Earth. Life 2023, 13, 757.
- Endo, I.; Matsumoto, T. Space flight/bedrest immobilization and bone. Bisphosphonate and the loss of bone mineral due to space flight or prolonged bed rest. Clin. Calcium. 2012, 22, 1863–1870. (In Japanese)
- Baran, R.; Wehland, M.; Schultz, H.; Heer, M.; Infanger, M.; Grimm, D. Microgravity-related changes in bone density and treatment options: A systematic review. Int. J. Mol. Sci. 2022, 23, 8650.
- Rengel, A.; Tran, V.; Toh, L.S. Denosumab as a pharmacological countermeasure against osteopenia in long duration spaceflight. Aerosp. Med. Hum. Perform. 2023, 94, 389–395.
- Acconcia, F.; Fiocchetti, M.; Busonero, C.; Fernandez, V.S.; Montalesi, E.; Cipolletti, M.; Pallottini, V.; Marino, M. The extra-nuclear interactome of the estrogen receptors: Implications for physiological functions. Mol. Cell Endocrinol. 2021, 538, 111452.
- Trevino, L.S.; Gorelick, D.A. The interface of nuclear and membrane steroid signaling. Endocrinology 2021, 162, bqab107.
- Thomas, P. Membrane progesterone receptors (mPRs, PAQRs): Review of structural and signaling characteristics. Cells 2022, 11, 1785.
- Thomas, P.; Pang, Y.; Camilletti, M.A.; Castelnovo, L.F. Functions of membrane progesterone receptors (mPRs, PAQRs) in nonreproductive tissues. Endocrinology 2022, 163, bqac147.
- Koszegi, Z.; Cheong, R.Y. Targeting the non-classical estrogen pathway in neurodegenerative diseases and brain injury disorders. Front. Endocrinol. 2022, 13, 999236.
- Goncalves, F.J.; Soares, F.A.; Pouso, M.R.; Longo, M.; Cairrao, E. Non-genomic effect of estradiol on the neurovascular unit and possible involvement in the cerebral vascular accident. Mol. Neurobiol. 2023, 60, 1964–1985.
- Schumacher, M.; Baulieu, E.E. neurosteroids: Synthesis and functions in the central and peripheral nervous systems. Ciba Found. Symp. 1995, 191, 90–106.
- Blacklock, A.D.; Johnson, M.S.; Krizsan-Agbas, D.; Smith, P.G. Estrogen increases sensory nociceptor neuritogenesis in vitro by a direct, nerve growth factor-independent mechanism. Eur. J. Neurosci. 2005, 21, 2320–2328.
- Krizsan-Agbas, D.; Pedchenko, T.; Smith, P.G. Neurotrimmin is an estrogen-regulated determinant of peripheral sympathetic innervation. J. Neurosci. Res. 2008, 86, 3086–3095.
- Vongpatanasin, W. Autonomic regulation of blood pressure in menopause. Semin. Reprod. Med. 2009, 27, 338–345.
- Wang, Q.; Cao, J.; Hu, F.; Lu, R.; Wang, J.; Ding, H.; Gao, R.; Xiao, H. Effects of estradiol on voltage-gated sodium channels in mouse dorsal root ganglion neurons. Brain Res. 2013, 1512, 1–8.
- Sample, S.J.; Racette, M.A.; Hao, Z.; Thomas, C.F.; Behan, M.; Muir, P. Functional adaptation in female rats: The role of estrogen signaling. PLoS ONE 2012, 7, e43215.
- Chen, Y.; Sun, Y.; Xue, X.; Ma, H. Comprehensive analysis of epigenetics mechanisms in osteoporosis. Front. Genet. 2023, 14, 1153585.
- Hart, D.A. Sex differences in musculoskeletal injury and disease risks across the lifespan: Are there unique subsets of females at higher risk than males for these conditions at distinct stages of the life cycle? Front. Physiol. 2023, 14.
- Seeman, E. Age- and menopause-related bone loss compromise cortical and trabecular microstructure. J. Gerontol. A Biol. Sci. Med. Sci. 2013, 68, 1218–1225.
- Ghasem-Zadeh, A.; Burghardt, A.; Wang, X.F.; Iuliano, S.; Bonaretti, S.; Bui, M.; Zebaze, R.; Seeman, E. Quantifying sex, race, and age specific differences in bone microstructure requires measurement of anatomically equivalent regions. Bone 2017, 101, 206–213.
- Wulf, M.J.; Tom, V.J. Consequences of spinal cord injury on the sympathetic nervous system. Front. Cell Neurosci. 2023, 17, 999253.
- Madden, K.S.; Felton, S.Y.; Felton, D.L.; Hardy, C.A.; Livant, S. Sympathetic nervous system modulation of the immune system. II. Induction of lymphocyte proliferation and migration in vivo by chemical sympathectomy. J. Neuroimmunol. 1994, 49, 67–75.
- Madden, K.S.; Moyihan, J.A.; Brenner, G.J.; Felton, S.Y.; Felton, D.L.; Livant, S. Sympathectic nervous system modulation of the immune system. III. Alterations in T and B cell proliferation and differentiation in vitro following chemical sympathectomy. J. Neuroimmunol. 1994, 49, 77–87.
- Grebe, K.M. Editorial: Regulation of the regulator: Sympathetic nervous system control of regulatory T cells. J. Leukoc. Biol. 2009, 86, 1269–1270.
- Wu, K.; Li, R.; Zhang, Y.; Liu, Y.M.; Wang, M.C.; Huang, J.; Zhu, C.; Zhang, J.; Yuan, X.; Liu, Q. The discovery of a new type of innervation in lymphoid organs. Physiol. Rep. 2023, 11, 15604.
- Kruszewska, B.; Felton, S.Y.; Moyihan, J.A. Alterations in cytokine and antibody production following chemical sympathectomy in two strains of mice. J. Immunol. 1995, 155, 4613–4620.
- Kirkland, L.G.; Barbe, C.G.; Hadaya, J.; Benson, P.V.; Wagner, B.M.; Tankovic, S.; Hoover, D.B. Sympathetic innervation of human and porcine spleens: Implications for between species variation in function. Bioelectron. Med. 2022, 8, 20.
- De Virgliis, F.; Oliva, V.M.; Kizil, B.; Scheiermann, C. Control of lymph node activity by direct local innervation. Trends Neurosci. 2022, 45, 704–712.
- Gonzalez-Ariki, S.; Husband, A.J. The role of sympathetic innervation of the gut in regulating mucosal immune responses. Brain Behav. Immun. 1998, 12, 53–63.
- Veny, M.; Grases, D.; Kucharova, K.; Lin, W.W.; Nguyen, J.; Huang, S.; Ware, C.F.; Ranscht, B.; Sedy, J.R. Contactin-1 is required for peripheral innervation and immune homeostatsis within the intestinal mucosa. Front. Immunol. 2020, 11, 1268.
- Madden, K.S.; Rajan, S.; Bellinger, D.L.; Felton, S.Y.; Felton, D.L. Age-associated alterations in sympathetic neural interactions with the immune system. Dev. Comp. Immunol. 1997, 21, 479–485.
- Afan, A.M.; Broome, C.S.; Nicholls, S.E.; Whetton, A.D.; Miyan, J.A. Bone marrow innervation regulates cellular retention in the murine haemopoietic system. Br. J. Haematol. 1997, 98, 569–577.
- Wang, X.; Xu, J.; Kang, Q. Neuromodulation of bone: Role of different peptides and their interactions (review). Mol. Med. Rep. 2021, 23, 32.

