Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Italia Di Liegro | -- | 1819 | 2023-07-20 11:19:14 | | | |
| 2 | Alfred Zheng | Meta information modification | 1819 | 2023-07-21 03:51:00 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Di Liegro, C.M.; Schiera, G.; Schirò, G.; Di Liegro, I. General Properties of Genes Encoding Histone Variants. Encyclopedia. Available online: https://encyclopedia.pub/entry/47031 (accessed on 28 July 2026).
Di Liegro CM, Schiera G, Schirò G, Di Liegro I. General Properties of Genes Encoding Histone Variants. Encyclopedia. Available at: https://encyclopedia.pub/entry/47031. Accessed July 28, 2026.
Di Liegro, Carlo Maria, Gabriella Schiera, Giuseppe Schirò, Italia Di Liegro. "General Properties of Genes Encoding Histone Variants" Encyclopedia, https://encyclopedia.pub/entry/47031 (accessed July 28, 2026).
Di Liegro, C.M., Schiera, G., Schirò, G., & Di Liegro, I. (2023, July 20). General Properties of Genes Encoding Histone Variants. In Encyclopedia. https://encyclopedia.pub/entry/47031
Di Liegro, Carlo Maria, et al. "General Properties of Genes Encoding Histone Variants." Encyclopedia. Web. 20 July, 2023.
Copy Citation
All the cells of an organism contain the same genome. However, each cell expresses only a minor fraction of its potential and, in particular, the genes encoding the proteins necessary for basal metabolism and the proteins responsible for its specific phenotype. The ability to use only the right and necessary genes involved in specific functions depends on the structural organization of the nuclear chromatin, which in turn depends on the epigenetic history of each cell, which is stored in the form of a collection of DNA and protein modifications. Among these modifications, DNA methylation and many kinds of post-translational modifications of histones play a key role in organizing the complex indexing of usable genes.
histone variants
H3.3 histone
genes encoding histone variants
1. Introduction
Since Gurdon’s experiments [1], based on the transplantation into enucleated oocytes of nuclei purified from somatic cells, it became clear that during differentiation, cells do not lose DNA and thus maintain a nucleus with intact potential to generate an entire organism. Despite the presence of the entire genome, however, each specialized cell expresses only a very small percentage of its genome. The reason for specific gene selection lies in the tridimensional organization of chromatin, which is a complex of DNA and proteins. Among these latter molecules, the most represented are histones, i.e., basic proteins that are highly conserved in evolution, which interact with DNA, allowing condensation of the nuclear genome in the very small volume of eukaryotic nuclei. The first level of chromatin organization is the nucleosome, in which about 147 base pairs (bp) of DNA are wrapped around a protein octamer formed by two molecules of each of the histones H2A, H2B, H3, and H4 (called “core” histones”). A fifth histone, the “linker” H1 histone then seals together the points at which DNA enters and exits the nucleosome. Interaction among H1 molecules allows the formation of more condensed DNA fibers [2][3][4][5][6][7][8].
Most importantly, the chromatin structure differs at the level of different genes that, depending on their structural organization, can be transcriptionally repressed or active, and this is why different cells are able to express different and specific genes, as well as a family of common genes involved in basic metabolism. Thus, specific properties and the behavior of somatic cells do not depend on changes in the genotype but on the specific arrangement of chromatin, according to the concept of “epigenetics” proposed by Waddington many decades ago [9], which is now widely accepted [4][10].
The chromatin structure is highly dynamic and changes in different cells during development and differentiation; it also changes in terminally differentiated cells in response to specific inducing factors, such as, for example, thyroid or steroid hormones [11]. At least three connected mechanisms are known to induce structural rearrangements of chromatin (Figure 1): (i) post-translational modification (PTM) of histone proteins [10][12] together with DNA methylation [10][13][14]; (ii) the activity of ATP-dependent complexes that are able to induce modifications in the structure/position of nucleosomes [15], and, finally, (iii) the synthesis and incorporation of histone variants into chromatin [7][16][17][18].
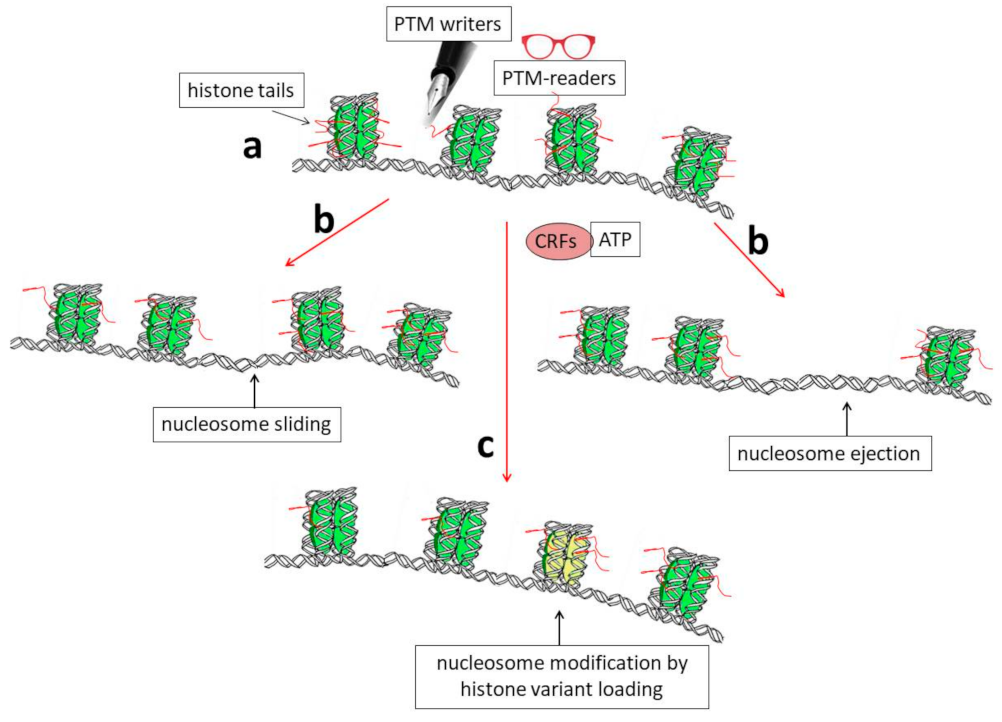
Figure 1. Schematic representation showing the biochemical mechanisms that allow dynamic modifications of chromatin organization. (a) Histone tails protrude from nucleosomes (red segments) and can be the targets of enzymes (PTM writers) that are able to introduce into them post-translational modifications (PTMs). These modifications can, in turn, directly affect DNA–histone interactions and/or allow chromatin structural modifications through the binding of other specific factors (PTM readers). (b) A modification of the nucleosome position by sliding or nucleosome ejection can be catalyzed by ATP-dependent chromatin-remodeling factors (CRFs). (c) Finally, the incorporation of histone variants into chromatin, such as H3.3, can induce structural and functional modifications of nucleosomes (shown as the yellow nucleosome core particle in the figure).
2. General Properties of Genes Encoding Histone Variants
Although highly conserved in evolution, and thus among different species, histone proteins exist under different isoforms in the same species. These isoforms seem to have different effects on chromatin structure and then on gene expression.
Interestingly, genes encoding the main histone species differ both in structure and expression from those encoding variants synthesized only in specific moments of cell differentiation. In particular, the main histone species (also known as replication-dependent or canonical histones) are synthesized exclusively during the S phase of the cell cycle when DNA is replicated. The corresponding genes are highly repeated and very often clustered; moreover, they do not contain introns, and the corresponding mRNAs are not poly-adenylated [19][20][21]. All these features are related to the necessity of having mRNAs that are immediately available for translation and then for degradation. On the other hand, genes encoding constitutive/variant histones (also known as replication-independent or non-canonical histones) are similar to all the other genes in that they are mostly unique genes that are transcribed, independently of DNA replication, into mRNAs that can contain introns and are polyadenylated [22]. These genes should be regulated during differentiation in order to produce proteins that are able to bind specific regions of chromatin, thus allowing activation/repression of specific genes.
Many years ago, the group showed that two histone variants, i.e., the linker histone H1.0 and the core histone H3.3, are specifically expressed in the rat brain during brain maturation [23]. Interestingly, a combination of run-on experiments on isolated nuclei and transcription inhibition using actinomycin D demonstrated that the two genes have an “open” structure and that H1.0 and H3.3 histone synthesis in the central nervous system (CNS) is largely regulated at the post-transcriptional level [24]. Indeed, people identified a group of proteins that are able to bind their mRNAs and cloned a couple of them (CSD-C2/PIPPin and LPI/PEP-19) [25][26][27][28][29]. Notably, in a very recent and interesting paper focused on the effects of hunger on neuronal histone modifications and the life span of the Drosophila fruit fly, the effects of a diet containing low amounts of branched-chain amino acids (BCAAs) were analyzed. In particular, the authors found that total histone H3 abundance decreased in flies fed a low BCAA diet, while H3 mRNA increased [30]. This observation suggested that post-transcriptional events might be of importance for general H3 metabolism. Moreover, the paper also reported that canonical H3 is evicted from chromatin and replaced with H3.3 [30].
It is also worth noting that two genes encoding the H3.3 histone are present in mammals: H3.3A and H3.3B (also called H3F3A and H3F3B), which are located on different chromosomes. While the distribution of exons and introns, as well as promoters and other regulatory regions, are different in the two genes, the corresponding proteins are identical [31] and also highly conserved in evolution. Thus, it is highly probable that the existence of the two genes is not important for having two proteins with different activities but, instead, because it offers the possibility to regulate the genes (and the related mRNAs) independently and/or with different mechanisms [32]. In particular, it was suggested that the two genes may have cell type-specific expression [33], although their overall activity in different tissues was reported to be quite similar [34].
Actually, the H3.3 protein is not so different with respect to the canonical H3.1 and H3.2 isoforms: indeed, it differs by only five and four amino acids, respectively, from them [35][36][37]. However, these amino acids are, for example, sufficient to allow H3.3 to interact with specific histone chaperones, such as the Death domain-associated protein (DAXX), the alpha-thalassemia/mental retardation X-linked protein (ATRX) complex, and the histone regulator A (Hira)/calcineurin-binding protein 1 (Cabin 1)/ubinuclein1 (Ubn1) complex, involved in its loading on chromatin [7][38][39][40][41]. The H3.3 interaction with these chaperones is determinant for its deposition on specific regions of the genome. It was shown, for example, that a mutation of the ATRX complex leads, as a consequence, to variation in the deposition of H3.3 and chromatin accessibility in association with an alteration in gene expression [42].
The H3.3 variant is indeed very often bound to active chromatin and regulates transcription. In 2002, Ahmad and Henikoff reported that in Drosophila cells, H3.3 is the only H3 species deposited in chromatin in a replication-independent way, and they suggested that this event might be responsible for the activation of genes previously silenced because of histone PTMs [43]. As a confirmation of its gene-activating function, the H3.3 histone was also found at the level of active enhancers [44][45][46].
It was also reported that some chromatin remodeling complexes, such as the SWItch/Sucrose Non-Fermentable (SWI/SNF) complex and, in particular, its subunit “T-rich interactive domain-containing protein 1A” (ARAD1A) are required for maintaining the H3.3 histone at the level of regulatory sequences, among which are the so-called super-enhancers [47]. On the other hand, by interacting with the ATRX/DAXX chaperones, H3.3 might also be loaded on pericentric heterochromatin and telomeres [38][48][49]. This event, together with lysine 56 (H3.3K56) acetylation, seems necessary for chromosome segregation in mammals. Indeed, it was shown that cell lines carrying the mutation K56R increase cell death and modify cell morphology [50].
Recently, a brain-specific function of the chromodomain-helicase-DNA binding protein 1 (CHD1) which is a member of the SWI/SNF family of chromatin remodeling complexes was reported in Drosophila. CHD1 is indeed involved in the loading of H3.3 in the fly brain, where it seems to contribute to the regulation of genes that control the homeostasis of hunger and satiety signals [51]. On the other hand, as a demonstration of the wide range of tissues and functions in which H3.3 histone is probably involved, it was also found to be essential for the chromatin transitions that accompany Drosophila male germline maturation [52].
Further work is required to understand how H3.3, which, as mentioned above, is not so different from the canonical H3 species, can stimulate transcription. Evidently, its sequence should contain features that are able to attract, directly or indirectly, the transcriptional apparatus to the genes to which it is bound. Interestingly, for example, the H3.3 amino-terminal tail contains a serine residue (S31) that is not present in the other H3 species (which contain, instead, an alanine at that position). Moreover, this serine can be phosphorylated, and it has been suggested that this might represent a feature determinant for preferential transcription [53]. It should be noted that S31 is actually S32 in the original amino acid sequence of the H3.3 protein; however, in the mature protein, it becomes S31 because the initiator methionine is immediately cleaved during translation ([54] and references therein). Notably, it was recently demonstrated that S31 phosphorylation can also modify the accessibility of regulatory factors at telomeres during replication, thus stabilizing heterochromatin probably by influencing the activity of histone lysine demethylase 4B (KDM4B) [55].
One possible link between H3.3 modification and gene expression was suggested by Martire and co-workers [56]. Using mouse embryonal stem cells (mESCs), these authors showed that the cells missing histone H3.3 cannot normally acetylate the enhancers that are activated during differentiation and, more specifically, show a reduction in the acetylation of the H3 histone at lysine 27. The normal ability to regulate acetylation would depend on the stimulation of p300 acetyltransferase by phosphorylation at specific sites of the H3.3 histone variant [56][57].
References
- Gurdon, J.B.; Elsdale, T.R.; Fischberg, M. Sexually mature individuals of Xenopus laevis from the transplantation of single somatic nuclei. Nature 1958, 182, 64–65.
- Kornberg, R.D. Chromatin structure: A repeating unit of histones and DNA. Science 1974, 184, 868–871.
- Luger, K.; Mäder, A.W.; Richmond, R.K.; Sargent, D.F.; Richmond, T.J. Crystal structure of the nucleosome core particle at 2.8 Å resolution. Nature 1997, 389, 251–260.
- Felsenfeld, G. A brief history of epigenetics. Cold Spring Harb. Perspect. Biol. 2014, 6, a018200.
- Allis, C.D.; Jenuwein, T. The molecular hallmarks of epigenetic control. Nat. Rev. Genet. 2016, 17, 487–500.
- Klemm, S.L.; Shipony, Z.; Greenleaf, W.J. Chromatin accessibility and the regulatory epigenome. Nat. Rev. Genet. 2019, 20, 207–220.
- Sokolova, V.; Sarkar, S.; Tan, D. Histone variants and chromatin structure, update of advances. Comput. Struct. Biotechnol. J. 2022, 21, 299–311.
- Hergeth, S.P.; Schneider, R. The H1 linker histones: Multifunctional proteins beyond the nucleosomal core particle. EMBO Rep. 2015, 16, 1439–1453.
- Waddington, C.H. The epigenotype. 1942. Int. J. Epidemiol. 2012, 41, 10–13.
- Kim, U.; Lee, D.-S. Epigenetic Regulations in Mammalian Cells: Roles and Profiling Techniques. Mol. Cells 2023, 46, 86–98.
- Kato, S.; Yokoyama, A.; Fujiki, R. Nuclear receptor coregulators merge transcriptional coregulation with epigenetic regulation. Trends Biochem. Sci. 2011, 36, 272–281.
- Strahl, B.D.; Allis, C.D. The language of covalent histone modifications. Nature 2000, 403, 41–45.
- Zhu, H.; Wang, G.; Qian, J. Transcription factors as readers and effectors of DNA methylation. Nat. Rev. Genet. 2016, 17, 551–565.
- Luo, C.; Hajkova, P.; Ecker, J.R. Dynamic DNA methylation: In the right place at the right time. Science 2018, 361, 1336–1340.
- Jiang, D.; Li, T.; Guo, C.; Tang, T.S.; Liu, H. Small molecule modulators of chromatin remodeling: From neurodevelopment to neurodegeneration. Cell Biosci. 2023, 13, 10.
- Shindo, Y.; Brown, M.G.; Amodeo, A.A. Versatile roles for histones in early development. Curr. Opin. Cell Biol. 2022, 75, 102069.
- Peng, J.; Zhang, W.J.; Zhang, Q.; Su, Y.H.; Tang, L.P. The dynamics of chromatin states mediated by epigenetic modifications during somatic cell reprogramming. Front. Cell. Dev. Biol. 2023, 11, 1097780.
- Simon, L.; Probst, A.V. Maintenance and dynamic reprogramming of chromatin organization during development. Plant J. 2023.
- Marzluff, W.F.; Gongidi, P.; Woods, K.R.; Jin, J.; Maltais, L.J. The human and mouse replication-dependent histone genes. Genomics 2002, 80, 487–498.
- Marzluff, W.F.; Wagner, E.J.; Duronio, R.J. Metabolism and regulation of canonical histone mRNAs: Life without a poly(A) tail. Nat. Rev. Genet. 2008, 9, 843–854.
- Amatori, S.; Tavolaro, S.; Gambardella, S.; Fanelli, M. The dark side of histones: Genomic organization and role of oncohistones in cancer. Clin. Epigenetics 2021, 13, 71.
- Talbert, P.B.; Henikoff, S. Histone variants at a glance. J. Cell Sci. 2021, 134, jcs244749.
- Castiglia, D.; Cestelli, A.; Scaturro, M.; Nastasi, T.; Di Liegro, I. H1.0 and H3.3B mRNA levels in developing rat brain. Neurochem. Res. 1994, 19, 1531–1537.
- Scaturro, M.; Cestelli, A.; Castiglia, D.; Nastasi, T.; Di Liegro, I. Posttranscriptional Regulation of H1.0 and H3.3B histone genes in differentiating rat cortical neurons. Neurochem. Res. 1995, 20, 969–976.
- Castiglia, D.; Scaturro, M.; Nastasi, T.; Cestelli, A.; Di Liegro, I. PIPPin, a putative RNA-binding protein specifically expressed in the rat brain. Biochem. Biophys. Res. Commun. 1996, 218, 390–394.
- Scaturro, M.; Nastasi, T.; Raimondi, L.; Bellafiore, M.; Cestelli, A.; Di Liegro, I. H1(0) RNA-binding proteins specifically expressed in the rat brain. J. Biol. Chem. 1998, 273, 22788–22791.
- Nastasi, T.; Scaturro, M.; Bellafiore, M.; Raimondi, L.; Beccari, S.; Cestelli, A.; Di Liegro, I. PIPPin is a brain-specific protein that contains a cold-shock domain and binds specifically to H1 degrees and H3.3 mRNAs. J. Biol Chem. 1999, 274, 24087–24093.
- Sala, A.; Scaturro, M.; Proia, P.; Schiera, G.; Balistreri, E.; Aflalo-Rattenbach, R.; Créau, N.; Di Liegro, I. Cloning of a rat-specific long PCP4/PEP19 isoform. Int. J. Mol. Med. 2007, 19, 501–509.
- Saladino, P.; Di Liegro, C.M.; Proia, P.; Sala, A.; Schiera, G.; Lo Cicero, A.; Di Liegro, I. RNA-binding activity of the rat calmodulin-binding PEP-19 protein and of the long PEP-19 isoform. Int. J. Mol. Med. 2012, 29, 141–145.
- Weaver, K.J.; Holt, R.A.; Henry, E.; Pletcher, S.D. Effects of hunger on neuronal histone modifications slow aging in Drosophila. Science 2023, 380, 625–632.
- Kumar, V.C.; Pai, R. Genes of the month: H3.3 histone genes: H3F3A and H3F3B. J. Clin. Pathol. 2021, 74, 753–758.
- Bryant, L.; Sangree, A.; Clark, K.; Bhoj, E. Histone 3.3-related chromatinopathy: Missense variants throughout H3-3A and H3-3B cause a range of functional consequences across species. Hum. Genet. 2023.
- Bush, K.; Cervantes, V.; Yee, J.Q.; Klein, R.H.; Knoepfler, P.S. A knockout-first model of H3f3a gene targeting leads to developmental lethality. Genesis 2023, 61, e23507.
- Bachu, M.; Tamura, T.; Chen, C.; Narain, A.; Nehru, V.; Sarai, N.; Ghosh, S.B.; Ghosh, A.; Kavarthapu, R.; Dufau, M.L.; et al. A versatile mouse model of epitope-tagged histone H3.3 to study epigenome dynamics. J. Biol. Chem. 2019, 294, 1904–1914.
- Elsaesser, S.J.; Goldberg, A.D.; Allis, C.D. New functions for an old variant: No substitute for histone H3.3. Curr. Opin. Genet. Dev. 2010, 20, 110–117.
- Filipescu, D.; Szenker, E.; Almouzni, G. Developmental roles of histone H3 variants and their chaperones. Trends Genet. 2013, 29, 630–640.
- Delaney, K.; Almouzni, G. Transcription-coupled H3.3 recycling: A link with chromatin states. Semin. Cell Dev. Biol. 2023, 135, 13–23.
- Lewis, P.W.; Elsaesser, S.J.; Noh, K.M.; Stadler, S.C.; Allis, C.D. Daxx is an H3.3-specific histone chaperone and cooperates with ATRX in replication-independent chromatin assembly at telomeres. Proc. Natl Acad. Sci. USA 2010, 107, 14075–14080.
- Goldberg, A.D.; Banaszynski, L.A.; Noh, K.M.; Lewis, P.W.; Elsaesser, S.J.; Stadler, S.; Dewell, S.; Law, M.; Guo, X.; Li, X.; et al. Distinct factors control histone variant H3.3 localization at specific genomic regions. Cell 2010, 140, 678–691.
- Smith, R.; Susor, A.; Ming, H.; Tait, J.; Conti, M.; Jiang, Z.; Lin, C.-J. The H3.3 chaperone Hira complex orchestrates oocyte developmental competence. Development 2022, 149, dev200044.
- Yang, Y.; Zhang, L.; Xiong, C.; Chen, J.; Wang, L.; Wen, Z.; Yu, J.; Chen, P.; Xu, Y.; Jin, J.; et al. HIRA complex presets transcriptional potential through coordinating depositions of the histone variants H3.3 and H2A.Z on the poised genes in mESCs. Nucleic Acids Res. 2022, 50, 191–206.
- Truch, J.; Downes, D.J.; Scott, C.; Gür, E.R.; Telenius, J.M.; Repapi, E.; Schwessinger, R.; Gosden, M.; Brown, J.M.; Taylor, S.; et al. The chromatin remodeller ATRX facilitates diverse nuclear processes, in a stochastic manner, in both heterochromatin and euchromatin. Nat. Commun. 2022, 13, 3485.
- Ahmad, K.; Henikoff, S. The histone Variant H3.3 marks active chromatin by replication-independent nucleosome assembly. Mol. Cell 2002, 9, 1191–1200.
- Chen, P.; Zhao, J.; Wang, Y.; Wang, M.; Long, H.; Liang, D.; Huang, L.; Wen, Z.; Li, W.; Li, X.; et al. H3.3 actively marks enhancers and primes gene transcription via opening higher ordered chromatin. Genes Dev. 2013, 27, 2109–2124.
- Deaton, A.M.; Gomez-Rodriguez, M.; Mieczkowski, J.; Tolstorukov, M.Y.; Kundu, S.; Sadreyev, R.I.; Jansen, L.E.; Kingston, R.E. Enhancer regions show high histone H3.3 turnover that changes during differentiation. Elife 2016, 5, e15316.
- Shi, L.; Wen, H.; Shi, X. The histone variant H3.3 in transcriptional regulation and human disease. J. Mol. Biol. 2017, 429, 1934–1945.
- Reske, J.J.; Wilson, M.R.; Armistead, B.; Harkins, S.; Perez, C.; Hrit, J.; Adams, M.; Rothbart, S.B.; Missmer, S.A.; Fazleabas, A.T.; et al. ARID1A-dependent maintenance of H3.3 is required for repressive CHD4-ZMYND8 chromatin interactions at super-enhancers. BMC Biol. 2022, 20, 209.
- Wong, L.H.; Ren, H.; Williams, E.; McGhie, J.; Ahn, S.; Sim, M.; Tam, A.; Earle, E.; Anderson, M.A.; Mann, J.; et al. Histone H3.3 incorporation provides a unique and functionally essential telomeric chromatin in embryonic stem cells. Genome Res. 2009, 19, 404–414.
- Udugama, M.; Chang, F.T.M.; Chan, F.L.; Tang, M.C.; Pickett, H.A.; McGhie, J.D.R.; Mayne, L.; Collas, P.; Mann, J.R.; Wong, L.H. Histone variant H3.3 provides the heterochromatic H3 lysine 9 tri-methylation mark at telomeres. Nucleic Acids Res. 2015, 43, 10227–10237.
- Fang, L.; Chen, D.; Zhang, J.; Li, H.; Bradford, B.; Jin, C. Potential functions of histone H3.3 lysine 56 acetylation in mammals. Epigenetics 2022, 17, 498–517.
- Schoberleitner, I.; Mertens, B.; Bauer, I.; Lusser, A. Regulation of sensory perception and motor abilities by brain-specific action of chromatin remodelling factor CHD1. Front. Mol. Neurosci. 2022, 15, 840966.
- Sakai, A.; Schwartz, B.E.; Goldstein, S.; Ahmad, K. Transcriptional and developmental functions of the H3.3 histone variant in Drosophila. Curr. Biol. 2009, 19, 1816–1820.
- Armache, A.; Yang, S.; Martínez de Paz, A.; Robbins, L.E.; Durmaz, C.; Cheong, J.Q.; Ravishankar, A.; Daman, A.W.; Ahimovic, D.J.; Klevorn, T.; et al. Histone H3.3 phosphorylation amplifies stimulation-induced transcription. Nature 2020, 583, 852–857.
- Chen, J.; Horton, J.; Sagum, C.; Zhou, J.; Cheng, X.; Bedford, M.T. Histone H3 N-terminal mimicry drives a novel network of methyl-effector interactions. Biochem. J. 2021, 478, 1943–1958.
- Udugama, M.; Vinod, B.; Chan, F.L.; Hii, L.; Garvie, A.; Collas, P.; Kalitsis, P.; Steer, D.; Das, P.P.; Tripathi, P.; et al. Histone H3.3 phosphorylation promotes heterochromatin formation by inhibiting H3K9/K36 histone demethylase. Nucleic Acids Res. 2022, 50, 4500–4514.
- Martire, S.; Gogate, A.A.; Whitmill, A.; Tafessu, A.; Nguyen, J.; Teng, Y.C.; Tastemel, M.; Banaszynski, L.A. Phosphorylation of histone H3.3 at serine 31 promotes p300 activity and enhancer acetylation. Nat. Genet. 2019, 51, 941–946.
- Tafessu, A.; O’Hara, R.; Martire, S.; Dube, A.L.; Saha, P.; Gant, V.U.; Banaszynski, L.A. H3.3 contributes to chromatin accessibility and transcription factor binding at promoter-proximal regulatory elements in embryonic stem cells. Genome Biol. 2023, 24, 25.
More
Information
Subjects:
Biochemistry & Molecular Biology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
649
Revisions:
2 times
(View History)
Update Date:
21 Jul 2023
Table of Contents



Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No