Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Julien A. Delbrouck | -- | 1286 | 2023-07-11 00:45:47 | | | |
| 2 | Conner Chen | Meta information modification | 1286 | 2023-07-11 04:24:24 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Delbrouck, J.A.; Desgagné, M.; Comeau, C.; Bouarab, K.; Malouin, F.; Boudreault, P. Steroidal (Glyco)Alkaloids: Classification. Encyclopedia. Available online: https://encyclopedia.pub/entry/46617 (accessed on 21 June 2026).
Delbrouck JA, Desgagné M, Comeau C, Bouarab K, Malouin F, Boudreault P. Steroidal (Glyco)Alkaloids: Classification. Encyclopedia. Available at: https://encyclopedia.pub/entry/46617. Accessed June 21, 2026.
Delbrouck, Julien A., Michael Desgagné, Christian Comeau, Kamal Bouarab, François Malouin, Pierre-Luc Boudreault. "Steroidal (Glyco)Alkaloids: Classification" Encyclopedia, https://encyclopedia.pub/entry/46617 (accessed June 21, 2026).
Delbrouck, J.A., Desgagné, M., Comeau, C., Bouarab, K., Malouin, F., & Boudreault, P. (2023, July 10). Steroidal (Glyco)Alkaloids: Classification. In Encyclopedia. https://encyclopedia.pub/entry/46617
Delbrouck, Julien A., et al. "Steroidal (Glyco)Alkaloids: Classification." Encyclopedia. Web. 10 July, 2023.
Copy Citation
Steroidal (glycol)alkaloids S(G)As are secondary metabolites made of a nitrogen-containing steroidal skeleton linked to a (poly)saccharide, naturally occurring in the members of the Solanaceae and Liliaceae plant families. The genus Solanum is familiar to all of us as a food source (tomato, potato, eggplant), but a few populations have also made it part of their ethnobotany for their medicinal properties. The recent development of the isolation, purification and analysis techniques have shed light on the structural diversity among the SGAs family, thus attracting scientists to investigate their various pharmacological properties.
Solanum
steroid
glycoalkaloid
1. Introduction
Nutrition is a vital act of communion with nature. Plants use the sun’s energy to transform minerals and gases into a cornucopia of organic molecules such as carbohydrates, lipids, terpenes, steroids, etc. Directly or indirectly, they supply all the energy and building blocks needed for the growth and maintenance of the human body. History shows that plants have been used for their medicinal properties in all civilizations and they continue today to be a source of bioactive molecules. The Solanum genus is comparatively a large one, encompassing over 1500 species and it is of particular importance in most of the world, as it includes popular food crops such as tomato (Solanum lycopersicum), potato (Solanum tuberosum) and eggplant (Solanum melongena). Two etymologies have been proposed: sol + anum (“of the Sun”) or solor + nus (“soothing”). Fittingly, it also contains various nightshade species used as folk medicine [1], such as the yellow-fruit nightshade (Solanum virginianum) used in Ayurveda medicine and the black nightshade (Solanum nigrum) used in both Western and Eastern traditional medicine. The genus Solanum is part of the Solanaceae family, which also includes the genera Capsicum (bell and chili peppers), Nicotiana (tobacco) as well as Datura, Mandragora and Atropa among others. It can thus be seen that many plants of this family are cultivated for their secondary metabolites like capsaicin and nicotine or are poisonous such as the deadly nightshade (Atropa belladonna).
At the interface between food, medicine, and poison, Steroidal Alkaloids (SAs) and their glycosidic versions are secondary metabolites that are especially prominent in the Solanum genus. Steroidal GlycoAlkaloids (SGAs) consist of a (poly)saccharide linked to a hydrophobic steroidal skeleton containing a nitrogen atom. They accumulate in different organs of the plants (leaves, roots, fruits, tubers, etc.) [2][3][4] and play a key role in the plant-environment interactions, in particular against bacterial and fungal attacks [5][6]. Their anti-nutrient properties and human toxicity are well-documented [3][7][8][9][10][11], while they more recently have attracted the attention of researchers for their wide range of pharmacological properties (anticancer, antibiotic, anti-inflammatory, etc.) [12][13][14][15]. For instance, Coramsine (SBP002) was an experimental chemotherapeutic drug composed of two steroidal glycoalkaloids (solamargine and solasonine) isolated from the species Solanum linneanum (devil’s apple) [16].
2. Steroidal (Glyco)Alkaloids: Classification
Steroidal alkaloids are nitrogenous derivatives of steroids and have been isolated and characterized from many organisms including animals (amphibians, sea sponges, etc.) and plants (Solanaceae, Liliaceae, etc.). They are important secondary metabolites with a wide range of potential therapeutic applications [17]. For instance, the steroidal alkaloid drug marketed Zytiga®, was approved in 2011 by the FDA for the treatment of metastatic castration-resistant prostate cancer [18]. The nitrogen atom is incorporated to the steroidal backbone either as heterocyclic ring, basic side chain or as a substituent at the C-3 position. Monomeric steroidal alkaloids are commonly classified into four groups based on their carbon heterocyclic skeleton: (1) Pregnane (C21); (2) Cyclopregnane (C24); (3) Cholestane (C27); (4) Others (Figure 1).
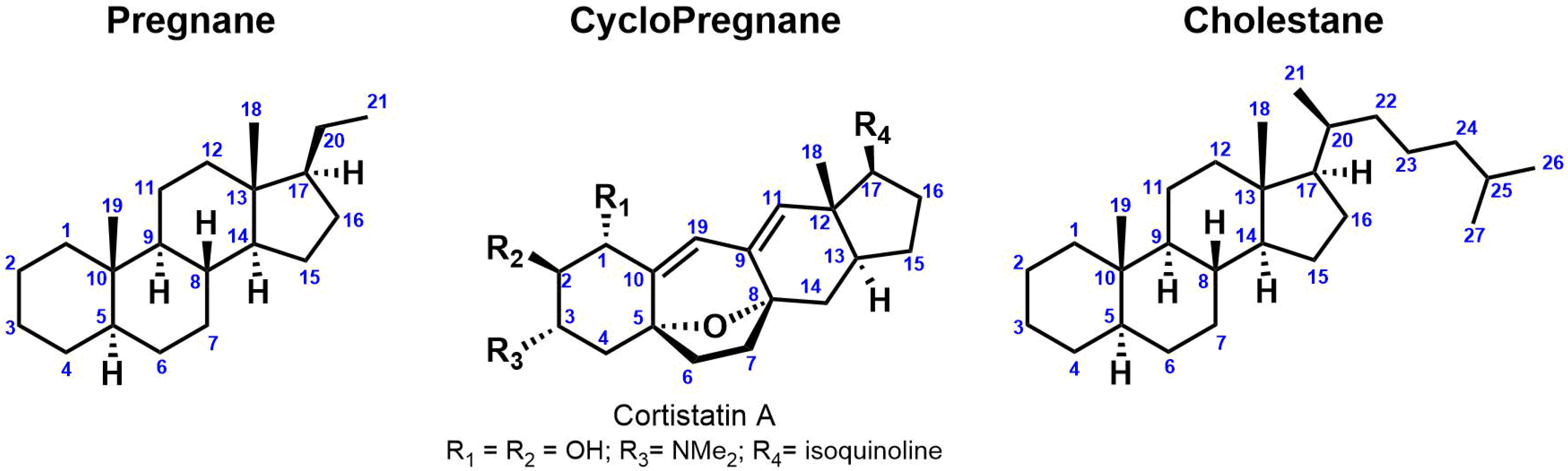
Figure 1. Steroid categories according to their carbon framework.
The occurrence of the Solanum steroidal alkaloid is well-documented and they mainly belong to C27-cholestane family [18][19]. These metabolites are characterized by the common ABCD steroid skeleton, and they are conventionally subdivided into three main types: spirosolane; solanidane (aka. Solanidine); and verazine (aka. 22/23,26-epiminocholestane) types (Figure 2).
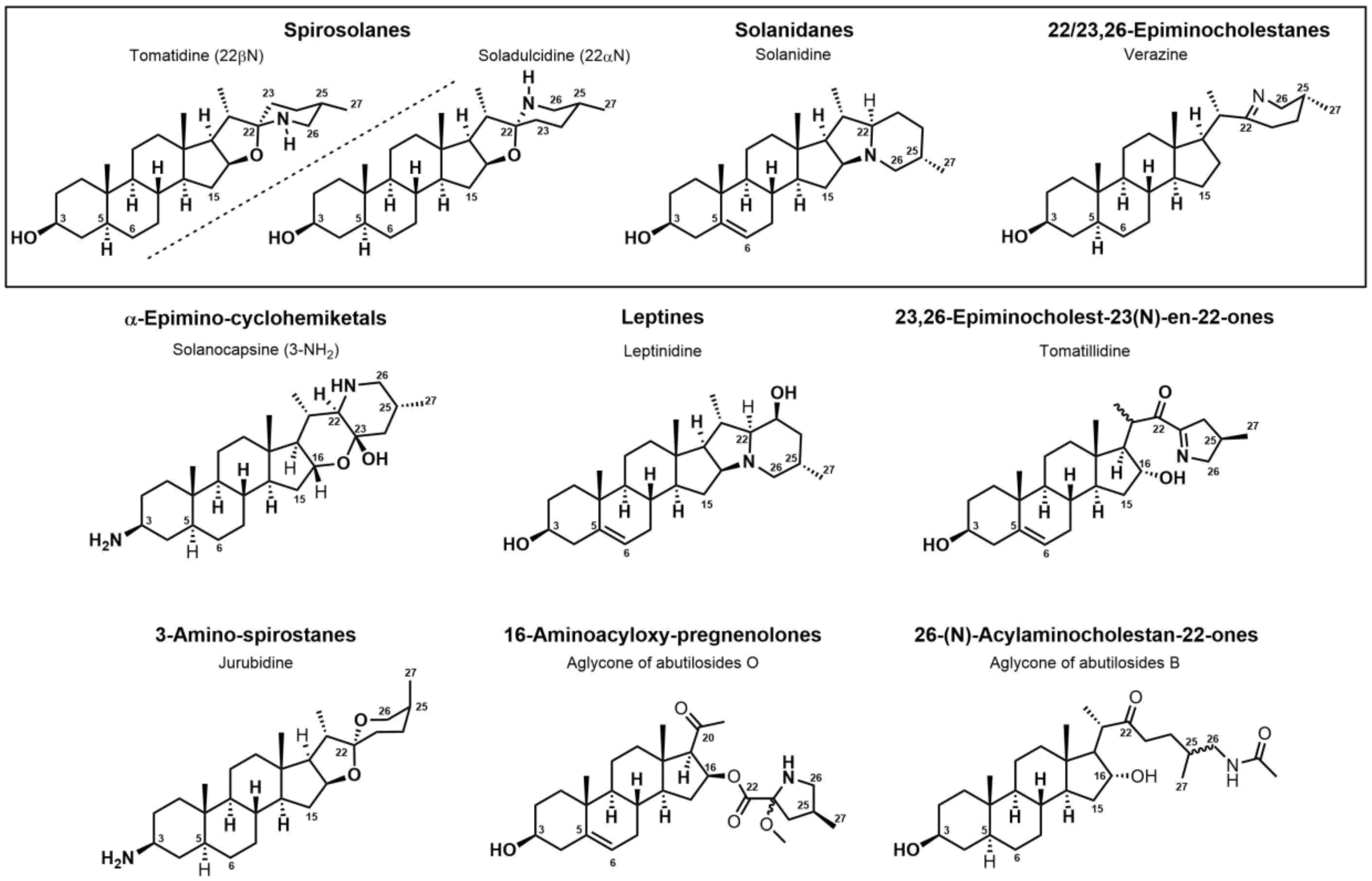
Figure 2. Classification of steroidal alkaloids.
The Spirosolane type is characterized by a 1-oxa-6-azapiro [4][5] decane ring system (E and F rings) and can be divided into two stereoisomeric subgroups: 22αN- and 22βN-spirosolanes (Figure 2). The C27 methyl group always take place in equatorial position within the Spirosolane family. The Solanidane family possess a fused indolizidine pattern with the 22R,25S configuration (Figure 2). Solanidine and its 5a,6-dihydro analog, demissidine, are the most common SAs of this structural entity. The Verazine group has a 22/23,26-epiminocholestane skeleton and differs by the non-conjugated N-containing F-ring and the absence of ring E (Figure 2). For analogs with a saturated piperidine F-ring, 22αN and 22βN-diastereoisomers also exist. Noteworthy, the C16 position can be saturated or substituted (e.g., -OH).
Over the last few decades, purification and extraction techniques of natural compounds have considerably progressed along with the abilities of researchers to elucidate complex chemical structures. Therefore, further phytochemical investigations resulted in the discovery of new metabolites with unique structural features thus leading to the creation of six additional (sub)types: α-Epiminocyclohemiketal; Leptine; 23,26-Epiminocholest-23(N)-en-22one; 3-Aminospirostane; 26-(N)-Acylaminocholestan-22-one; and 16-Aminocycloxy-pregnenolone (Figure 2) [18][19].
The most prevalent metabolite of the α-Epiminocyclohemiketal family is solanocapsine, which contains two nitrogen atoms and is characterized by its α-epiminocyclohemiketal functionality. Its worth mentioning that 3-hydroxy analogues were discovered later. Leptines are solanidane equivalents with a 23-hydroxy/acetyl group. For instance, leptinidine is the 23β-hydroxy analog of solanidine (Figure 2). The 23,26-Epiminocholest-23(N)-en-22-one type is defined by the C22 exocyclic carbonyl function and the unsaturated bond between the C23 and the nitrogen of the five-membered ring. The 3-Aminospirostane group (aka jurubidine type) is constituted of 3-amino analogues of spirostane steroids that contrast from previous metabolites by the oxygen replacing the basic nitrogen N22. Another family characterized by the lack of N22 basic nitrogen is the 26-(N)-Acylaminocholestan-22-one. These metabolites exhibit an almost identical side chain to the cholestane framework, likely because the N-acetyl group prevents the cyclization. The 16-Aminoacyloxy-pregnenolone type is unique with its C21 pregnane skeleton instead of the usual C27 cholestane framework. It is a recent category with only two SGAs isolated to date [20][21].
It is to be noted that steroidal alkaloids often exist in nature as glycosides named Steroidal GlycoAlkaloids (SGAs) [22]. The biosynthesis of Solanum SGAs has been partially elucidated and it relies on the cytosolic mevalonate pathway with cholesterol as a key precursor [23][24]. It can be separate into two main operations: the aglycone construction and glycosylation. A significant number of glycoalkaloid metabolism (GAME) genes are involved in the initial regulation of the structural modification of the cholesterol backbone by hydroxylation, transamination, oxidation, etc., followed by glycosylation reactions [25].
The polar entity in SGAs includes one to several monosaccharides in various combinations (branched, linked or linear). The most common units are D-galactose, D-glucose, D-xylose, L-arabinose and L-rhamnose which interconnected offer a variety of disaccharides, trisaccharides and tetrasaccharides [19]. The predominant glycosidic chains of the solanum SGAs are solatriose, chacotriose, lycotetraose and commertetraose (Figure 3). The glycosidic linkage mostly occurs at the C3 position of the steroidal backbone (monodesmosidic) by contrast to bisdesmodic metabolites that own an additional sugar at another location of the steroidal skeleton (e.g., Esculeoside A) [26]. The α/β anomeric configuration of the glycosidic bond at the C-3 hydroxy position of the aglycone is driven by the D/L-configuration of the monosaccharide. The oligosaccharide chain plays a pivotal role in directing biological applications as observed for the SGA Tomatine and its non-glycosidic analog Tomatidine that display different therapeutical properties.
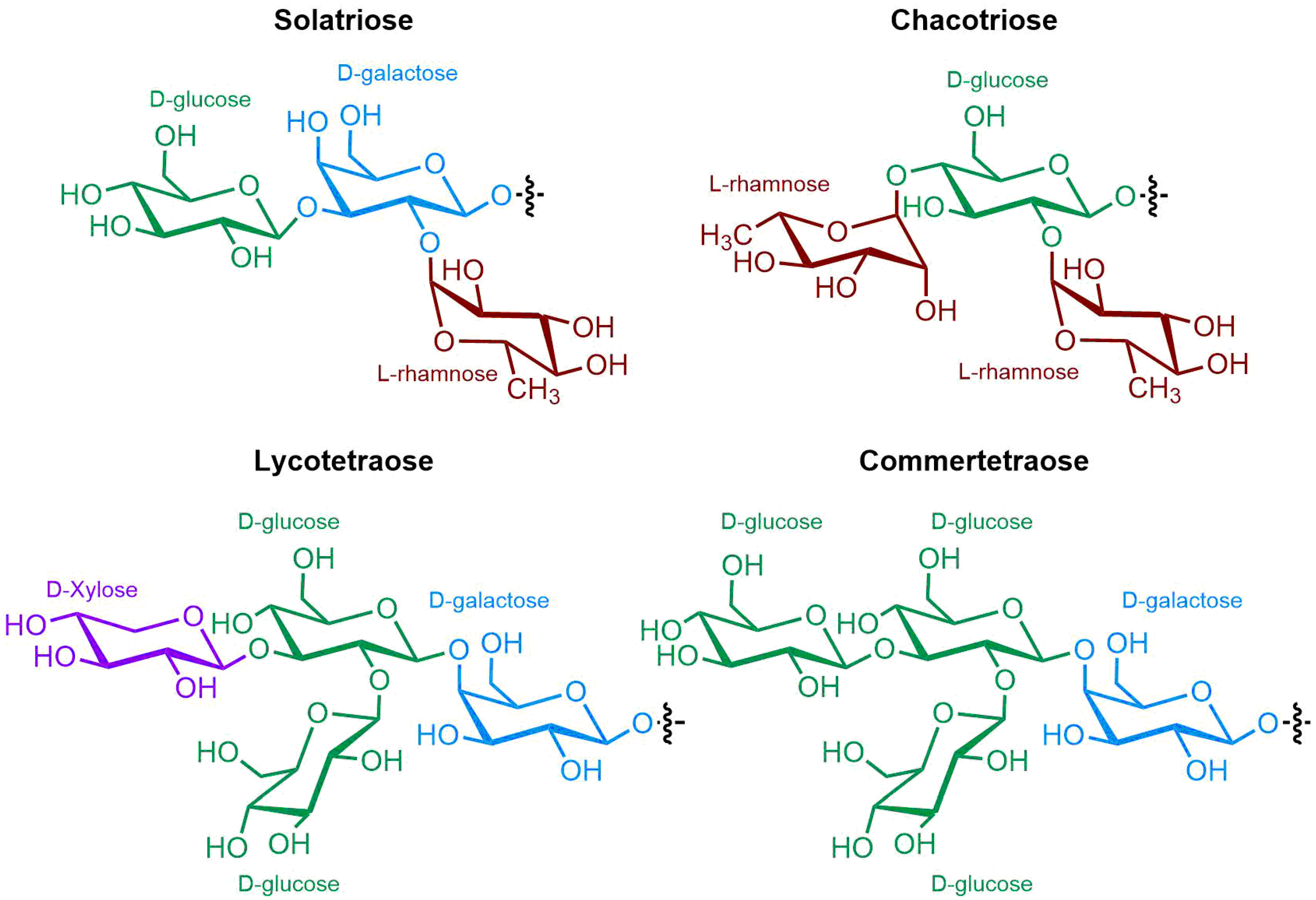
Figure 3. Chemical structures of the main oligosaccharide units of Solanum SGAs.
The popularity of SGAs arises from the structural diversity offered by the aglycone structure, the nature of the side chain and the combination of carbohydrates. The aglycone unit often shares the tetracyclic 5α-androstane of the C27 cholestane skeleton while divergence mostly occurs by substitutions; rearrangement of the side chain (C20–C27); and the position of substituents. Interestingly, 5α-saturated and 5-unsaturaded analogs have been detected for almost all categories of Solanum SGAs.
References
- Jain, R.; Sharma, A.; Gupta, S.; Sarethy, I.P.; Gabrani, R. Solanum Nigrum: Current Perspectives on Therapeutic Properties. Altern. Med. Rev. 2011, 16, 78–85.
- Friedman, M.; Levin, C.E. Dehydrotomatine Content in Tomatoes. J. Agric. Food Chem. 1998, 46, 4571–4576.
- Friedman, M. Potato Glycoalkaloids and Metabolites: Roles in the Plant and in the Diet. J. Agric. Food Chem. 2006, 54, 8655–8681.
- Bednarz, H.; Roloff, N.; Niehaus, K. Mass Spectrometry Imaging of the Spatial and Temporal Localization of Alkaloids in Nightshades. J. Agric. Food Chem. 2019, 67, 13470–13477.
- Tingey, W.M. Glycoalkaloids as Pest Resistance Factors. Am. Potato J. 1984, 61, 157–167.
- Dahlin, P.; Muller, M.C.; Ekengren, S.; McKee, L.S.; Bulone, V. The Impact of Steroidal Glycoalkaloids on the Physiology of Phytophthora Infestans, the Causative Agent of Potato Late Blight. Mol. Plant Microbe Interact. 2017, 30, 531–542.
- Friedman, M. Tomato Glycoalkaloids: Role in the Plant and in the Diet. J. Agric. Food Chem. 2002, 50, 5751–5780.
- Fogelman, E.; Oren-Shamir, M.; Hirschberg, J.; Mandolino, G.; Parisi, B.; Ovadia, R.; Tanami, Z.; Faigenboim, A.; Ginzberg, I. Nutritional Value of Potato (Solanum tuberosum) in Hot Climates: Anthocyanins, Carotenoids, and Steroidal Glycoalkaloids. Planta 2019, 249, 1143–1155.
- Chowanski, S.; Winkiel, M.; Szymczak-Cendlak, M.; Marciniak, P.; Manczak, D.; Walkowiak-Nowicka, K.; Spochacz, M.; Bufo, S.A.; Scrano, L.; Adamski, Z. Solanaceae Glycoalkaloids: Alpha-Solanine and Alpha-Chaconine Modify the Cardioinhibitory Activity of Verapamil. Pharm. Biol. 2022, 60, 1317–1330.
- EFSA Panelon Contaminants in the Food Chain (CONTAM); Schrenk, D.; Bignami, M.; Bodin, L.; Chipman, J.K.; del Mazo, J.; Hogstrand, C.; Hoogenboom, L.; Leblanc, J.; Nebbia, C.S.; et al. Risk Assessment of Glycoalkaloids in Feed and Food, in Particular in Potatoes and Potato-derived Products. EFS2 2020, 18, e06222.
- Smith, D. Potato Glycoalkaloids: Some Unanswered Questions. Trends Food Sci. Technol. 1996, 7, 126–131.
- Friedman, M.; Lee, K.-R.; Kim, H.-J.; Lee, I.-S.; Kozukue, N. Anticarcinogenic Effects of Glycoalkaloids from Potatoes against Human Cervical, Liver, Lymphoma, and Stomach Cancer Cells. J. Agric. Food Chem. 2005, 53, 6162–6169.
- Al Sinani, S.S.S.; Eltayeb, E.A. The Steroidal Glycoalkaloids Solamargine and Solasonine in Solanum Plants. S. Afr. J. Bot. 2017, 112, 253–269.
- Sbhatu, D.B.; Abraha, H.B. Preliminary Antimicrobial Profile of Solanum incanum L.: A Common Medicinal Plant. Evid. Based Complement. Alternat. Med. 2020, 2020, 3647065.
- Kenny, O.M.; McCarthy, C.M.; Brunton, N.P.; Hossain, M.B.; Rai, D.K.; Collins, S.G.; Jones, P.W.; Maguire, A.R.; O’Brien, N.M. Anti-Inflammatory Properties of Potato Glycoalkaloids in Stimulated Jurkat and Raw 264.7 Mouse Macrophages. Life Sci. 2013, 92, 775–782.
- Van der Most, R.G.; Himbeck, R.; Aarons, S.; Carter, S.J.; Larma, I.; Robinson, C.; Currie, A.; Lake, R.A. Antitumor Efficacy of the Novel Chemotherapeutic Agent Coramsine Is Potentiated by Cotreatment With CpG-Containing Oligodeoxynucleotides. J. Immunother. 2006, 29, 134–142.
- Dey, P.; Kundu, A.; Chakraborty, H.J.; Kar, B.; Choi, W.S.; Lee, B.M.; Bhakta, T.; Atanasov, A.G.; Kim, H.S. Therapeutic Value of Steroidal Alkaloids in Cancer: Current Trends and Future Perspectives. Int. J. Cancer 2019, 145, 1731–1744.
- Xiang, M.-L.; Hu, B.-Y.; Qi, Z.-H.; Wang, X.-N.; Xie, T.-Z.; Wang, Z.-J.; Ma, D.-Y.; Zeng, Q.; Luo, X.-D. Chemistry and Bioactivities of Natural Steroidal Alkaloids. Nat. Prod. Bioprospect. 2022, 12, 23.
- Eich, E. Solanaceae and Convolvulaceae: Secondary Metabolites; Springer: Berlin/Heidelberg, Germany, 2008.
- Ono, M.; Takara, Y.; Egami, M.; Uranaka, K.; Yoshimitsu, H.; Matsushita, S.; Fujiwara, Y.; Ikeda, T.; Nohara, T. Steroidal Alkaloid Glycosides, Esculeosides C and D, from the Ripe Fruit of Cherry Tomato. Chem. Pharm. Bull. 2006, 54, 237–239.
- Yoshimitsu, H.; Nishida, M.; Nohara, T. Steroidal Glycosides from the Fruits of Solanum abutiloides. Phytochemistry 2003, 64, 1361–1366.
- Heftmann, E. Biogenesis of Steroids in Solanaceae. Phytochemistry 1983, 22, 1843–1860.
- Akiyama, R.; Watanabe, B.; Nakayasu, M.; Lee, H.J.; Kato, J.; Umemoto, N.; Muranaka, T.; Saito, K.; Sugimoto, Y.; Mizutani, M. The Biosynthetic Pathway of Potato Solanidanes Diverged from That of Spirosolanes Due to Evolution of a Dioxygenase. Nat. Commun. 2021, 12, 1300.
- Sonawane, P.D.; Jozwiak, A.; Panda, S.; Aharoni, A. ‘Hijacking’ Core Metabolism: A New Panache for the Evolution of Steroidal Glycoalkaloids Structural Diversity. Curr. Opin. Plant Biol. 2020, 55, 118–128.
- Cardenas, P.D.; Sonawane, P.D.; Pollier, J.; Vanden Bossche, R.; Dewangan, V.; Weithorn, E.; Tal, L.; Meir, S.; Rogachev, I.; Malitsky, S.; et al. GAME9 Regulates the Biosynthesis of Steroidal Alkaloids and Upstream Isoprenoids in the Plant Mevalonate Pathway. Nat. Commun. 2016, 7, 10654.
- Nohara, T.; Ono, M.; Ikeda, T.; Fujiwara, Y.; El-Aasr, M. The Tomato Saponin, Esculeoside A. J. Nat. Prod. 2010, 73, 1734–1741.
More
Information
Subjects:
Pharmacology & Pharmacy
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.3K
Revisions:
2 times
(View History)
Update Date:
11 Jul 2023
Table of Contents




Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No