Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Ionela Hotea | -- | 5555 | 2023-07-06 10:30:52 | | | |
| 2 | Camila Xu | Meta information modification | 5555 | 2023-07-06 10:50:41 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Hotea, I.; Sirbu, C.; Plotuna, A.; Tîrziu, E.; Badea, C.; Berbecea, A.; Dragomirescu, M.; Radulov, I. Integrating (Nutri-)Metabolomics into the One Health Tendency. Encyclopedia. Available online: https://encyclopedia.pub/entry/46504 (accessed on 22 July 2026).
Hotea I, Sirbu C, Plotuna A, Tîrziu E, Badea C, Berbecea A, et al. Integrating (Nutri-)Metabolomics into the One Health Tendency. Encyclopedia. Available at: https://encyclopedia.pub/entry/46504. Accessed July 22, 2026.
Hotea, Ionela, Catalin Sirbu, Ana-Maria Plotuna, Emil Tîrziu, Corina Badea, Adina Berbecea, Monica Dragomirescu, Isidora Radulov. "Integrating (Nutri-)Metabolomics into the One Health Tendency" Encyclopedia, https://encyclopedia.pub/entry/46504 (accessed July 22, 2026).
Hotea, I., Sirbu, C., Plotuna, A., Tîrziu, E., Badea, C., Berbecea, A., Dragomirescu, M., & Radulov, I. (2023, July 06). Integrating (Nutri-)Metabolomics into the One Health Tendency. In Encyclopedia. https://encyclopedia.pub/entry/46504
Hotea, Ionela, et al. "Integrating (Nutri-)Metabolomics into the One Health Tendency." Encyclopedia. Web. 06 July, 2023.
Copy Citation
The term ‘metabolomics’ was introduced in the literature in 1998 and refers to the study of small molecules in a biological sample. In 2001, a group of biochemists founded the Society of Metabolomics, and the use of this technology in various fields of research has increased greatly since 2005. Nutrimetabolomics, or nutritional metabolomics, is an integral part of metabolomics with the goal of examining individual functional responses to different diets, analysing specific dietary biomarkers for targeted foods and diets, and investigating the interrelationship between risk factors for certain diseases and different diets both in the human and veterinary science fields.
metabolomics
nutrimetabolomics
One Health
public health
1. Introduction
With recent world developments, the emergence of new diseases endangering global health creates particular challenges for the medical field. To meet these challenges, novel, highly precise technologies can be utilised to facilitate clinical investigations and identify therapeutic and preventive solutions.
Here, the term ‘(nutri-)metabolomics’ is used to refer to both metabolomics and nutrimetabolomics. Metabolomics, one of these novel technologies, is the analysis of the total profile of metabolites within a system (cell, tissue, or organism) in a certain time period and under certain conditions. It is an integral part of the ‘omics’ sciences and makes a direct link with the body’s phenotype, providing biochemical information in addition to genomic and proteomic data [1][2][3]. Metabolomic technology is constantly evolving and becoming widely used in an increasing number of fields, including agriculture, environmental chemistry, biotechnology, and, in particular, medical sciences for clinical diagnosis, toxicology, nutrition, drug progress, and health and disease management [3][4][5].
Nutrimetabolomics, or nutritional metabolomics, is an integral part of metabolomics with the goal of examining individual functional responses to different diets, analysing specific dietary biomarkers for targeted foods and diets, and investigating the interrelationship between risk factors for certain diseases and different diets both in the human and veterinary science fields [3][6][7]. Nutrimetabolomic technology aims to determine individual human and animal reactions to nutrition, as well as to identify and implement personalized nutritional plans for optimal health [3][8].
One Health considers the association of several decision-making authorities at the local, national, and global levels with the aim of establishing interdisciplinary collaborations to ensure optimal living conditions and health for people, animals, and the environment. The most recent diseases in animals and humans (SARS-CoV-2, Ebola, avian flu (H5N1), swine flu (H1N1), etc.) are examples of global issues and the increased vulnerability of humans, animals, and the environment to new disease outbreaks [9][10][11][12].
2. Metabolomics and Nutrimetabolomics
Given that nutrimetabolomics is a branch of metabolomics, here, one term or the other will be used, depending on the context, although the study refers to both technologies—metabolomics and nutrimetabolomics.
The term ‘metabolomics’ was introduced in the literature in 1998 and refers to the study of small molecules in a biological sample. In 2001, a group of biochemists founded the Society of Metabolomics [13], and the use of this technology in various fields of research has increased greatly since 2005 [14][15]. Among the ‘omics’ technologies (genomics, transcriptomics, and proteomics), metabolomics has become increasingly utilized, especially in the last decade. Metabolomics can be described as the overall analysis of small molecules in a biological fluid (blood, urine, culture broth, cell extract, etc.), produced or transformed in the body, as a result of the intervention of a stimulus (nutritional factors, stress, environment, drugs, etc.) [15]. All metabolites that make up the ‘metabolome’ represent the molecular fingerprint of an organism [16]. The profile of the metabolome (metabolic phenotype or ‘metabotype’) reflects the biological state of an organism (Figure 1) [16][17].
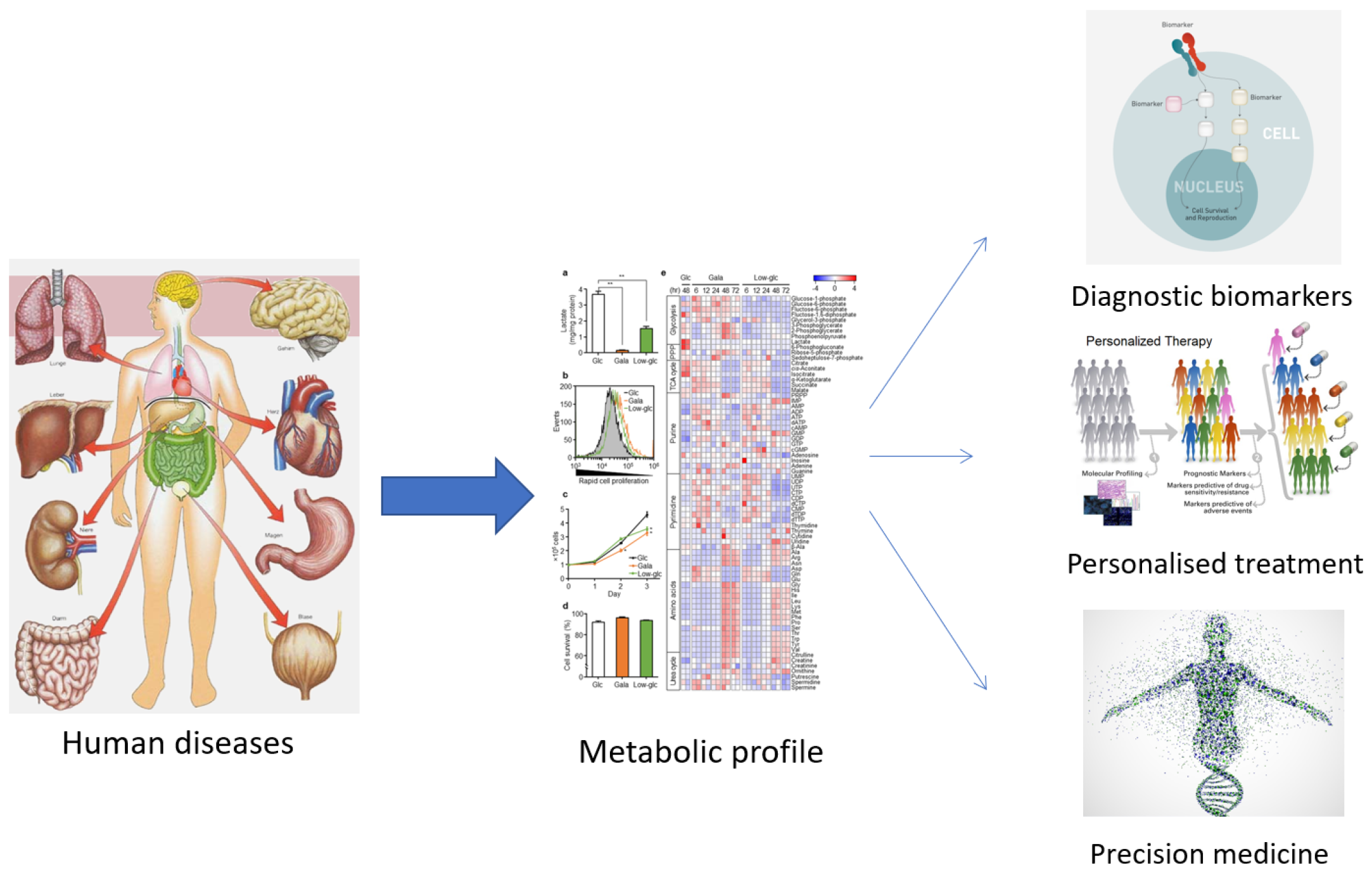
Figure 1. The usefulness of establishing the metabolic profile for medicine.
As can be observed from the numerous studies found on search platforms, metabolomics is a frequently applied technique. Interesting results have been obtained in the pharmaceutical (personal response to drug treatment, efficacy and/or toxicity of drugs), medical (biomarkers for disease diagnosis and prevention), and plant science fields (molecular structure, GMOs, etc.) [15][18].
Although metabolomics has recently been applied in nutrition, it is gaining increasing attention in this field [8]. Nutrition research has entered a new era, in which increased diet diversity and the relationships between nutrition and genotype, lifestyle, and diseases are investigated in great detail, with new methodological approaches, including ‘omics’ technologies to identify responses to various stimuli that have not been determined using traditional approaches [19][20]. Post-genomic technology also provides new methods and opportunities for research in the field of nutrition to explain individual differences in metabolism and assimilation of food and nutrients [15].
Nutritional fields combine traditional nutritional methods with genotyping and phenotyping, molecular epidemiology, and bioinformatics.
Nutrigenomics was the first nutrition domain to emerge investigating the effects of nutrients on genetic expression profiles and assessing how an individual’s genotype may influence nutrient uptake, excretion, or activity [15][21].
The next field that evolved was nutriproteomics, which analyses molecular and cellular changes in protein expression in response to nutrients and studies the interaction of proteins with nutrients in the food consumed [15][22].
Nutrimetabolomics is the dynamic and multivariate research of biological fluid or tissue responses to nutritional stimuli. This technology analyses the direct or indirect effect of diet on metabolism [15].
Nutrimicrobiomics refers to the genetic structure and functional capacity of microbial populations and the possibilities of the intestinal microbiome to process food and nutrients [15].
Food has long been known to influence the health and well-being of individuals, and nutrients have also been used as medicines to treat and prevent disease [16]. Research in this area has shown that diseases are often linked to poor or unbalanced nutrition [15][16][23][24]. At present, nutrition research has reached the level where metabolomics (nutrimetabolomics) is used to phenotype the nutritional status of individuals and facilitate the discovery of new biomarkers of specific nutrients or metabolic dysfunctions [15][25]. Moreover, metabolomic methods can be used to investigate the component metabolites of food in biological fluids and/or tissues, to study their bioavailability, the body’s response to a particular diet, and to analyse certain foods or nutraceuticals [16][26].
Food and nutrition science focus not only on the interrelationship between diseases and the consumption of macro- and micronutrients, but also study bioactive molecules that may be present in the diet in very small quantities [27]. These molecules can interact with various metabolic pathways and directly or indirectly modulate health, after being transformed by the intestinal microbiota [15]. Nutrimetabolomics offers the most accessible way to investigate the impact of nutritional interference on health, because the metabolic configurations of easily obtained biological fluids, such as blood, plasma, and urine, contain valuable information on both genetic heritage and environmental influences, including the contributions of dietary nutrients and their microbial transformations [15][28].
3. (Nutri-)Metabolomics in Human Medicine
Metabolomics developed rapidly in human medicine. It is primarily used to identify various biomarkers for the diagnosis, prevention, and treatment of various diseases, but also to investigate the role of the intestinal microbiota in the assimilation of dietary nutrients and to establish personalized nutrition plans or detailed food descriptions [15].
To identify the interconnections between lifestyle, diet, and health, it is necessary to establish specific biomarkers that provide information about nutritional profiles. In order to identify them, several studies have been launched to establish and develop nutritional markers [29][30] and to demonstrate their implications in the occurrence and development of metabolic diseases (obesity, diabetes), chronic diseases (HBP, CVD) or other diseases (cancer) [15][31][32][33][34][35][36][37][38].
In this regard, over time, the role of amino acids, fatty acids, or glucose metabolites in the onset and development of obesity or diabetes has been analysed [39][40][41][42]. It is known that the incidences of obesity/overweight and blood pressure (BP) risk values (prehypertensive/hypertensive) are globally widespread. A critical BP is a major risk factor for coronary heart disease and stroke, while obesity can, over time, lead to the onset of insulin resistance and type 2 diabetes or associated metabolic and cardiovascular diseases [15][43][44][45]. Various studies have been conducted on the influence of healthy diets on reducing the risk of developing cardiovascular diseases, type 2 diabetes, or various cancer types [15][46][47][48].
It is well known that nutrition is essential for maintaining life quality. Without nutrients, an organism cannot survive. Balanced nutrition can contribute to good health. Metabolomics is a key tool in modern nutritional research for analysing the number of calories, the ratio of food to fat, protein, and carbohydrate content, or the general intake of nutrients (Figure 2). Metabolomics is also used to analyse non-nutritive molecules that are apparently not vital to the body, but which, by their presence or absence, could affect the health and well-being of the body [16]. Thus, natural nutritive and non-nutritive molecules can be used as biomarkers for specific food consumption. These dietary or nutritional biomarkers can then be used to estimate the intake and quality of nutrients in the food consumed [15][24].
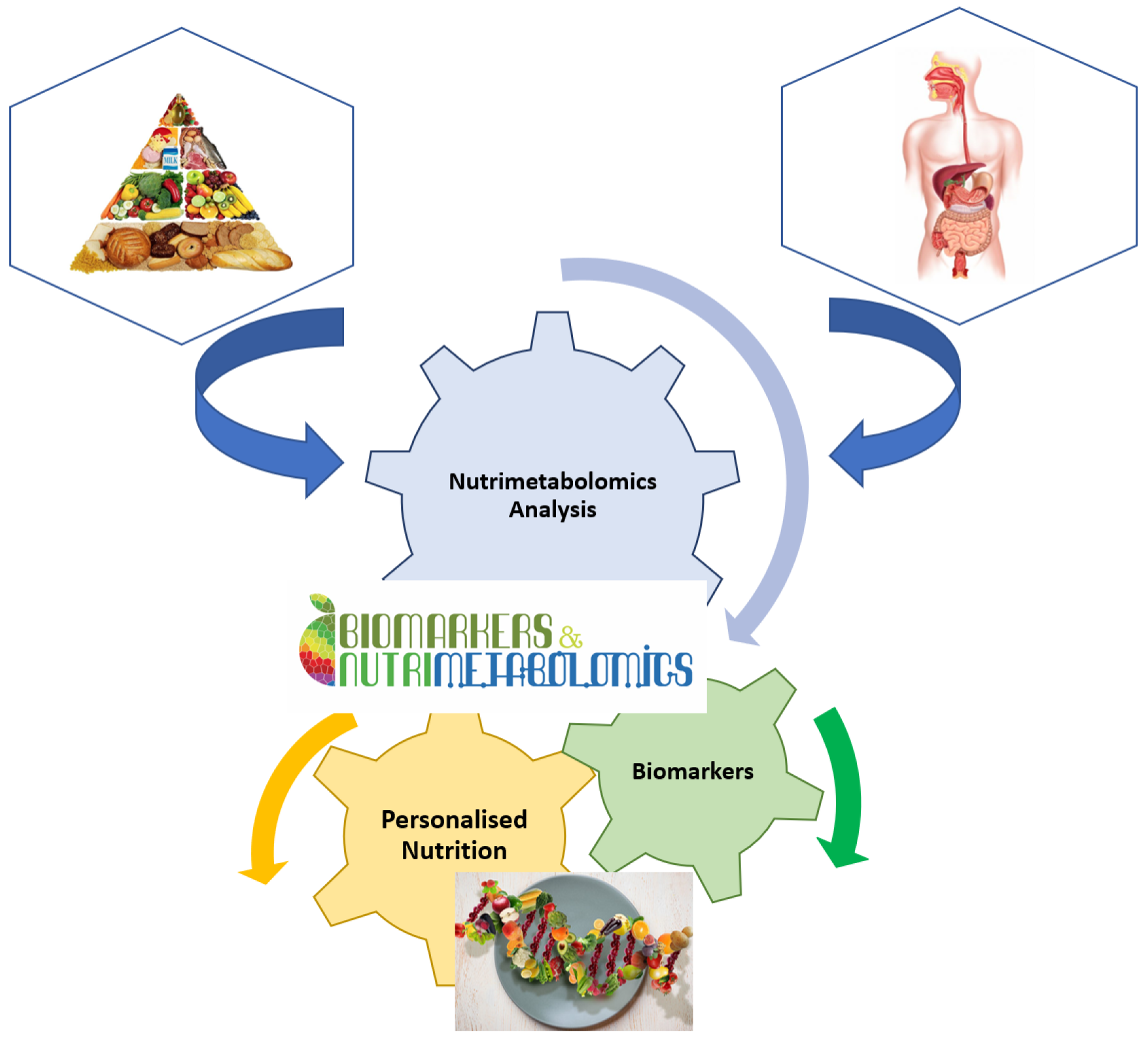
Figure 2. Implication of nutrimetabolomics in nutrition scrience.
Biomarkers are usually specific to each nutrient, food compound, or complex food [15]. For example, recent studies have identified plasma trimethylamine oxide as a marker of fish consumption [49][50] or methylhistidines in urine as a marker of animal protein intake [51]. Another example would be the blood level of long-chain omega-3 fatty acids, identified as an indicator for seafood consumption. Alkylresorcinols can be identified as markers of whole grain consumption, whereas total plasma carotenoids can indicate fruit and vegetable intake [52][53][54][55]. Furthermore, proline betaine has been established as an indicator of citrus fruit consumption [56][57][58][59][60]. Such studies provide evidence that various compounds can be identified as food biomarkers, and their presence or concentration in biological fluids can be a tool to identify the diet consumed [15]. This type of analysis is new in that, by identifying nutrients (i.e., identifying foods consumed by establishing the metabolic profile) the opportunity is provided to evaluate different diets, to establish certain dietary patterns, and to objectively classify individuals, depending on the types of diets they consume [15][61].
Studies are underway regarding the role of intestinal microorganisms in nutritional status and human health. Metabolic changes between intestinal and host microbial populations, i.e., co-metabolism, can influence the metabolic profile of the host (metabotype), an aspect highlighted by studies in this field [62][63][64][65][66]. It has been shown that the intestinal microbiota can predispose or contribute to certain human diseases. Thus, establishing the quantitative profile of intestinal microorganisms may predict certain diseases by identifying metabolites or metabolic processes that will be affected at some point [67]. The identified metabolites may function as metabolomic biomarkers for assessing disease evolution and the body’s response to specific treatments. Considering the significant interindividual differences in the population of intestinal microorganisms and host-microbe interactions, individual variations in susceptibility to certain diseases can be identified, as well as the responses of each organism to different pharmaceutical or nutritional interventions. This underscores the need to develop nutritional modulation strategies through the creation and use of personalized diets, as well as individualized therapeutic regimens to increase healthcare efficiency [67].
The future goal of nutrition research is to assess the metabolic response of each individual to diet. Personalized nutrition is the process by which individuals will modify their lifestyle and diet according to the information they have about their current health or future disease predispositions [15][68]. The theory of personalized nutrition started from the need to apply specific nutritional recommendations for population groups with similar characteristics [15]. The division of subjects into groups first requires the determination of the metabotype of each individual and then, depending on their characteristic metabolic type and physiological state, a nutrition style that suits them is defined. Following this process, personalized nutritional plans can be designed to modify and control the diet for the benefit of health. Therefore, metabolomics is very important in establishing the nutritional phenotype of each individual and identifying specific biomarkers to characterize various nutrients (food metabolome) or different metabolic dysfunctions [15][69].
Recently, an increasing number of publications have emerged highlighting the connections between food science and other fields, such as agriculture, biology, medicine, genetics, or veterinary science [70][71][72][73][74]. Metabolomics is a technology that can facilitate interdisciplinary connections by associating health with food quality and nutritional value [15]. Foodomics is the determination of food components. Macro- (proteins, fats, or carbohydrates) and micronutrients (vitamins, minerals, or other molecular components) determine the nutritional value of edible compounds necessary to maintain human health [16][75][76][77]. Most of the food metabolome is composed of phytochemicals. The best-known phytochemicals, highlighted by metabolomics studies, are fruit polyphenols, tomato lycopene or soy isoflavones [16][70][75][78][79]. Metabolomics can also identify chemicals in the environment, such as insecticides, herbicides, fungicides, antimicrobials, toxins, and other contaminants, that may be present in food and that could be harmful to the health of individuals who consume it [16][80][81][82][83]. Metabolomic studies have also been performed to establish the effects of genetically modified plants on the quality and nutritional value of foods and their by-products [84][85]. The origins of foods can also be determined based on their metabolic profile [86][87]. Furthermore, metabolomic studies can be used to analyse the effects of production technologies applied to transform raw materials into finished food on their nutritional, biochemical, or sensory value [77][88][89]. In today’s society, these studies are becoming increasingly important, as consumers are increasingly aware and concerned about the quality and origin of the food they consume, as well as the effects of food processing on the nutritional value of food.
For both human and veterinary medicine, the use of metabolomic assessment to ensure food safety is of common interest. Moreover, studies have analysed and highlighted differences in metabolites between foods of animal origin contaminated with micro-organisms or parasites and uncontaminated foods [90][91][92]. Consumers are increasingly interested in organic food, mainly due to concerns about food quality and safety, as well as the publicity of the idea that organic products are healthier and safer than conventional foods. The relationship between different types of agriculture (organic vs. conventional) and the various metabolic components that have been identified through the use of diets of various origins in animal feed has attracted particular interest [15][93][94]. Thus, foodomics is important for ensuring public health on two levels: in the short term, it can be involved in clinical interventions to treat various metabolic dysfunctions, such as diabetes and obesity; and in the long run, it can be used in preventive public health strategies to prevent the occurrence of certain diseases [93].
4. (Nutri-)Metabolomics in Veterinary Medicine
The use of metabolomics in veterinary medicine is more limited compared with its use in the human medical field, in which the applications of this advanced technology have been intensively explored. Although the application of metabolomics in veterinary medicine is slightly behind that in human medicine, there has recently been a growing interest in this method, especially because it is non-invasive and only requires biological fluids, particularly for the investigation of animal health or disease (Figure 3) [95][96].
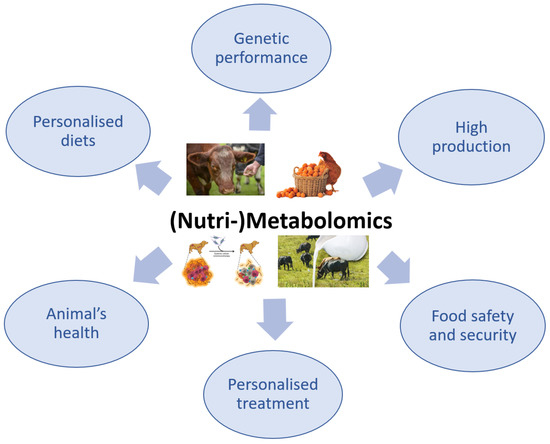
Figure 3. The importance of (Nutri-)Metabolomics in veterinary science.
Human diseases are frequently studied by inducing the disease in laboratory animals, especially rodents [97][98]. It should be noted that the use of animals in the study of diseases is performed in compliance with the ethical principle of avoiding harm. In animal model studies, various pathological conditions are induced to understand the molecular phenomena associated with various diseases and their complications [98]. The veterinary field may offer the possibility to study some spontaneously occurring diseases in animals, even in parallel with the appearance of the disease in humans, both in terms of the disease phenotype and pathogenesis [97]. Metabolomics has been used to study a variety of common diseases, such as type 2 diabetes [99][100], different types of cancer [101][102], and congenital metabolic errors [97][103][104]. This technique has also been applied to the study of the effects of different medicinal regimens [105] or dietary interventions [6][106][107][108][109], toxins [110][111][112], and stress [98][101] on health.
Although the majority of metabolomic studies in veterinary medicine have focused on pets and primarily investigated the pathogenesis and diagnosis of various diseases, studies on farm animals have also been conducted [73]. This is due to the fact that pets and farm animals are increasingly being studied as a reference model for human diseases. In this context, increasing emphasis has been placed on precise comparative analyses of human and animal diseases to accurately characterize the disease pathobiology. The closest animal species to humans, from an anatomo-physiological and pathophysiological point of view, are dogs and pigs [113]. These species are frequently studied using metabolomic tools to obtain a more detailed perspective on the pathobiology of diseases in humans. These types of studies have proven to be very effective, due to the use of standardized environmental conditions in terms of animal feeding and housing [114][115][116].
However, some researchers argue that canine models would be most appropriate for metabolomic studies due to the similarities in physiology and pathophysiology between dogs and humans. Thus, canine models have high clinical significance for human medical research [117][118]. Canine subjects can be used in metabolomic research to identify specific biomarkers associated with many common diseases, including cancer, heart disease [119][120][121][122], liver disease [123], and parasitic or infectious diseases [124][125]. Furthermore, studies of common neuropsychiatric disorders have been performed using canine models [117][126]. Given the similarities in anxiety manifestation and symptoms in humans and dogs, canine subjects could be considered a suitable model for the study of human anxiety, thus contributing to the knowledge of disease mechanisms at the molecular level [117]. Dogs could also be helpful in the study of human ADHD, as it has been observed that dogs can spontaneously exhibit specific ADHD behaviours [126].
Studies of strictly veterinary medical interest using metabolomics have also been con-ducted—in particular, studies aiming to identify methods to increase the quality of life of animals and improve their health [95][96]. Furthermore, metabolomic studies in pets have evaluated differences in the metabolite profiles of dogs and cats [127] and identified specific metabolic fingerprints for various dog breeds [128][129][130] as well as specific toxicological biomarkers in dogs [131][132][133]. Moreover, the metabolomic profile of cerebrospinal fluid of healthy and epileptic dogs has been investigated [96][134][135].
Studies in nutrition physiology using metabolomics highlighted differences in the production of greenhouse gas precursors by establishing personalized diets for ruminants [136]. In the field of farm animal research, metabolomics can also be applied to investigate the effects of nutritional programming on genetics, i.e., monitoring the epigenetic effects of diet during early development [90][91]. For maximum efficiency of the genetic selection response related to the traits pursued, such as disease resistance, performance or product quality, the researchers identified precise biomarkers that can be used to predict the manifestation of phenotypic characteristics [137]. Thus, for metabolomic research in the field of genetics, using livestock species provides several benefits, compared with using human or laboratory animal populations, such as multi-generation genealogies, long-term selection lines with large phenotypic differences, routine population phenotyping, targeted mating, opportunities to standardize environmental conditions, and a genome organization very similar to that of humans—more similar than that of most laboratory animal species [73][116][138].
In livestock farming, the identification of the metabolomic profile can be a starting point for formulating future diets or specific treatments to increase animal production, animal welfare, and the quality of animal products for human consumption [139].
Nutrition is a complex process, conditioned by several factors. Nutritional metabolomics is a significant technology for studying the relationship between an organism and its diet, as well as the interactions between the organism and its genetics, lifestyle, or even gastrointestinal flora [8][127]. The development of pet nutrition science has led to an increase in the lifespan of dogs and cats and a significant improvement in their quality of life. Elucidating the interactions between the nutrient molecules consumed through diet and the pathophysiological mechanisms that can be encountered in the various systems of an organism provides the possibility to identify new intervention methods and clinical management of the patient [140]. In humans, the individual is considered the main element in the emergence of diversity in nutritional metabolomic research, due to interindividual differences in genetic background, age, sex, gastrointestinal flora, and lifestyle. In the case of pets, nutrition studies are less diverse, enabling the conduct of longer-term research in controlled environments with low variation and consistency in sample collection [127].
The body’s biofluids, such as blood, plasma, and urine, contain many metabolites in the form of small molecules, such as amino acids, lipid fractions, or sugars, some of which may function as specific biomarkers and a way to monitor pathological conditions or responses of the body to nutritional interventions [107][141]. Metabolomics can be a useful tool in understanding individual responses to different diets and developing personalized nutritional formulas to improve health [108][115][142][143]. In most studies, personalized nutrition refers to human medicine; however, in an increasing number of recent studies in the field of veterinary medicine, the theory of personalized medicine appears [95].
A future goal in the field of animal nutrition will be to develop metabolomic studies to understand the effects of applying personalized diets using systems biology approaches. This will rely on the interconnection of a large number of data resulting from biological processes involving nutrients and non-nutritive compounds present in various ingredients used in animal feed, to manage the production and/or health status of animals as needed [96].
5. (Nutri-)Metabolomics in the Context of One Medicine
The One Health concept is a global strategy that aims to expand interdisciplinary collaborations and communications in all aspects of health care for humans, animals and the environment [144].
The interconnection between human and veterinary medicine was outlined in the nineteenth century by the German pathologist, Rudolf Virchow (1821–1902). He said: ‘between animal and human medicine there are no dividing lines—nor should there be. The object is different but the experience obtained constitutes the basis of all medicine’ [145][146].
The One Health strategy was initiated in 2007 in collaboration with the American Medical Association and the American Veterinary Medical Association to defend, im-prove, and promote the health and well-being of all species by encouraging and supporting the teamwork of human physicians, veterinarians, and health researchers [147]. The concept of One Health is not new, but its importance has been increasingly recognized in recent years. A result of 21st century concerns, the One Health, One Medicine, One World theory is part of a broader set of research programs and policies, including biosecurity, food security, translational medicine, and global health, with the goal of removing interdisciplinary barriers. This is the scenario in which the future of the One Health initiative will be developed [148].
One Health aims to increase the quality of life of all species, human and animal, given that about two-thirds (60.3%) of emerging infectious diseases result from zoonoses, most of which originate in the wild (71.8%), according to the studies of Frank et al. in 2008 [149]. Karesh et al., in 2012, showed that over 60% of human infectious diseases are caused by pathogens common to animals [148][150]. In this regard, metabolomics can help elucidate specific metabolic pathways and identify in vivo methods of infection, as well as the mechanisms of action of pathogens, which could contribute to discovering novel preventive or therapeutic strategies [125].
Kafsack et al., in 2010, reported the main existing zoonoses, which included brucellosis, bovine tuberculosis, rabies, leptospirosis, human African trypanosomiasis, and malaria [124]. Other protozoan parasites that are the causative agents of serious infections in both animals and humans, including host-life-threatening diseases, are Toxoplasma gondii (toxoplasmosis), Leishmania spp. (leishmaniasis), Cryptosporidium spp. (cryptosporidiosis), and Giardia spp. (giardiasis). While these diseases are becoming less common in developed countries, they continue to endanger the health and lives of a significant number of the population in other parts of the world. Moreover, these pathogens infect other non-specific host species, such as insects, birds, or even mammals, to ensure their complete life cycle [124][151]. The metabolic adaptation of these pathogens can provide them increased resistance and allow them to exploit nutrient sources in variable niches. Detailed analysis of the metabolic profile of each developmental stage of infectious pathogens and the degree of adaptation and modulation of their metabolism to that of the host organism, as well as the host’s immune response to these infections, can ensure the development of new therapeutic and medicinal strategies for humans and animals [125].
From the perspective of One Health, One Medicine, the study of zoonotic diseases is particularly important, given that it involves the inevitable and complex interaction of linked biological systems: humans, animals, and the environment [124]. The biological system comprises all living organisms and their relations with the environment. Thus, metabolomics research cannot be viewed singularly; all the functional systems of an organism are correlated with each other and with the environment. Thus, a complex analysis of metabolites and a multidisciplinary interpretation are needed to understand the overall functioning of organisms. For example, the analysis of plant metabolomics cannot be separated from that of mammals, because plant matter is the main source of food and nutrients for animals.
The One Health approach must also include non-communicable diseases, which, according to the World Health Organization, affect the lives of over 36 million people each year [152]. Among the non-communicable diseases common to humans and animals, investigated by numerous studies, cardiovascular diseases, coronary heart disease, hypertension, obesity, diabetes, various types of cancer, epilepsy, etc. can be mentioned [124][153][154][155]. Because both humans and pets (dogs and cats) can be affected by the same diseases, research on diseases in one species may be useful in the early intervention for other affected species [124]. Determining the metabolic profile (metabolomics/metabonomics), compared to other ‘omics’ profiles, has the advantage of mirroring the entire ecosystem of the body, which provides an overview of the functionality of the whole organism. These data are still difficult to analyse, especially at the level of an individual, due to the diversity of consumed food and thus the wide range of nutrients that enter the body. Due to this inevitable complexity, metabolomics research has sought to focus on the use of animal models whose genetic profile and environmental conditions can be more easily monitored compared with those of the human population [15].
Since the 20th century, when canaries were first used as carbon monoxide detectors in coal mines to prevent poisoning in miners, animals have been commonly used as sentinels for various pathologies, to maintain public health [156]. The use and monitoring of sentinel animals and the collection of all data on the occurrence of diseases in animal populations help to identify sources of disease, analyse the effectiveness of drugs or preventive intervention schemes, highlight the epidemiology of pathogens, or design an early intervention plan, thereby ensuring public health [157][158].
Epidemiological research in the veterinary field has several advantages over epidemiological research conducted in humans, such as a shorter period of disease development, an easier process to obtain necropsy and histopathological data, and lower costs. Unlike other types of laboratory studies (e.g., cell cultures), sentinel animal studies allow a more accurate study of conditions manifested in humans [159][160].
The One Health trend aims to support and encourage comparative studies on diseases that can be found in both humans and animals, the most common being obesity, diabetes, autoimmune disorders, and cancer [161][162]. Studies that use metabolomics to study these diseases aim to understand disease pathophysiology and identify biomarkers or therapies to improve public health.
Much of the research that has used sentinel animals to elucidate common pathologies has focused on the study of different types of cancer in pets, especially dogs, which live very close to humans, intimately sharing the same environment and consuming approximately the same categories of food. This is because most cancers found in dogs have been shown to have about the same pathological characteristics, biological basis, clinical manifestations, proportional morbidity, and identified risk factors (including eating habits), as those found in humans [158]. Thus, metabolomic or nutrimetabolomic studies involving detailed analyses of cancer in dogs complement current knowledge in human medicine on the efficacy of immunotherapy, prolonged release of drugs, gene therapy, and the clinical-pathological picture of this ubiquitous disease [163][164]. In the future, these types of ‘omics’ studies will allow the identification of personalized treatments and contribute to the development of personalized medicine in both humans and animals.
Recently, there has been a continuous increase in the average body weight among the human population, resulting in an increase of obesity. As the link between the occurrence of obesity in humans and pets is proving to be increasingly close and complex, much more than what was known in the past, obesity should be seen as a unique health issue and addressed in the context of One Health [154][165]. Through a psychological analysis, obesity in both dogs and their owners could be seen as an involuntary transfer of attitude towards the feeding process or eating habits of owners to their dogs and as an orientation towards the humanization of pets [165][166]. Nutrimetabolomic studies in obese animals can explain the conditions of obesity establishment and its pathophysiology, so as to prevent associated diseases, such as heart disease or diabetes.
As humans and pets share the same environment, the same ecosystem, studies in which animals can be used as sentinels for humans are very appropriate; however, there are instances when humans can also serve as sentinels in some animal health circumstances [167]. Given that metabolomic studies are more advanced in human medicine, the latest medical discoveries can also be applied in veterinary medicine, for the benefit of animal health, helping to identify gaps in the control of animal diseases.
‘Omics’ technologies are fundamental in advancing One Health knowledge. The application of these advanced technologies, together with a multidisciplinary collaboration, are the key tools in improving not only human and animal health, but also food safety and security and, implicitly, ecosystem health [168].
Food safety refers to the preparation, handling and storage of food using methods that ensure the prevention of foodborne illness [169]. Foodborne illnesses are serious problems that can threaten public health. The number of cases of foodborne diseases, including those caused by waterborne pathogens, is increasing [146]. Infectious diseases and food safety are interrelated. Animal disease influences animal production and, thus, the availability of food of animal origin for human consumption. In addition, food insecurity and malnutrition are aggravating factors for opportunistic infections [148]. Thus, plant and animal food safety is an area in which metabolomics can be successfully used, for identifying specific biomarkers of food contamination [170].
The World Food Summit described food security as ‘when all people, at all times, have physical, social, and economic access to sufficient, safe and nutritious food’ [148][171]. In addition to quantity, food quality is also an extremely important aspect of food security. In this context, dogs and cats can serve as sentinels to identify possible food contamination, including chemical contamination, or to determine the effects of consuming genetically modified plants and their derived food and feed products [148][172]. Therefore, in the context of One Health, ‘plant health’ must also be seen as an integrated aspect, especially since plant metabolomics is a frequently studied field [173][174].
Whereas in less developed countries there are people who suffer from hunger due to lack of food, in developed countries there are widespread micronutrient deficiencies, also known as hidden hunger [175]. Recent research in the field of nutrition and the implementation of its results in public health has been shown to have an important effect on reducing the incidence of diseases caused by deficiencies of essential nutrients. However, metabolic disorders are not only caused by deficiencies of essential nutrients but also by micronutrient or non-essential nutrient deficiencies. The increasing frequency of metabolic changes in energy regulation, manifested by diabetes, obesity, or atherosclerosis, has affected a large part of the world’s population, including citizens of the world’s most developed countries [115][176][177]. Nutrimetabolomics can be used for the early identification of the effects of nutrient deficiencies on the human body. These studies can be performed on animals first, as they are cheaper and less time consuming [178].
The usual methods of nutritional analysis that focus on assessing the link between diet and health, in general, do not cover the detailed understanding of the interdependence between a single nutrient and disease occurrence. Using such a perspective, and specific analyses, unique disease biomarkers can be identified and used for disease prevention. To determine the metabolic health of individuals, a more precise approach to nutritional assessment is needed, and this is possible using nutrimetabolomics [115].
Nutrient deficiencies in humans are closely linked to the availability of micronutrients in food (animal or plant origin), and this is dependent on the availability of micronutrients in animal feed and soil (for plants). However, the presence of micronutrients in food sources is not enough; the body’s ability to metabolize some nutrients is also essential because metabolism is closely correlated with health. These interdependencies are part of the concerns of the One Medicine, One Health concept (Figure 4).
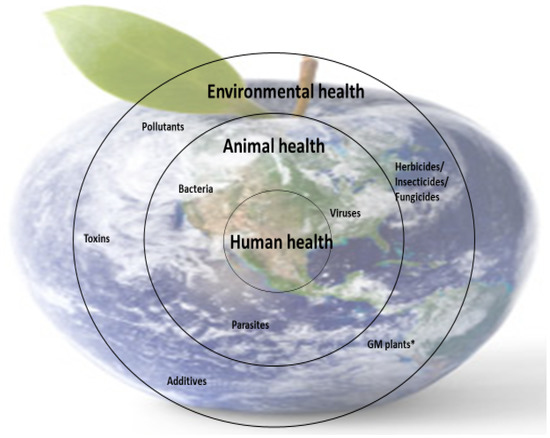
Figure 4. Schematic representation of the importance of One Medicine. * GM plants—genetically modified plants.
The holistic approach of the One Health concept remains one of the main ways to study the direct and underlying causes of food insecurity, malnutrition, and poor health and maximize human, animal, and environmental well-being. The added value of the One Health approach is that the benefits to human health are achieved through simultaneous investigations into human and animal health.
The future of mankind depends on a symbiotic relationship between humans, animals, and the environment. The rapid evolution of the planet towards industrialization, highly advanced technologies, and massive urbanization with a tendency towards human domination has led to an ecosystem imbalance and an increased vulnerability of human health. These trajectories have put humanity in unprecedented situations, with a rapid multiplication of emerging and re-emerging infectious diseases, as shown by the 2019 SARS-CoV-2 pandemic, amplified by the establishment of antimicrobial resistance and associated with an exponential increase in noncommunicable diseases.
In these situations, precise and rapid tools are needed to intervene in the face of threatening challenges to public health. In this context, the integration of metabolomics in most research areas will contribute to identifying customized solutions that will help restore the symbiosis between the human, animal, and plant populations and the environment, under the comprehensive “umbrella’’ of the One Health strategy.
References
- Jones, M.D.; Rainville, P.D.; Isaac, G.; Wilson, I.D.; Smith, N.W.; Plumb, R.S. Ultra high resolution SFC–MS as a high throughput platform for metabolic phenotyping: Application to metabolic profiling of rat and dog bile. J. Chromatogr. B 2014, 966, 200–207.
- Kirwan, J. Metabolomics for the practising vet. Practice 2013, 35, 438–445.
- Carlos, G.; dos Santos, F.P.; Fröehlich, P.E. Canine metabolomics advances. Metabolomics 2020, 16, 16.
- Kuehnbaum, N.L.; Britz-McKibbin, P. New Advances in Separation Science for Metabolomics: Resolving Chemical Diversity in a Post-Genomic Era. Chem. Rev. 2013, 113, 2437–2468.
- Rochfort, S. Metabolomics Reviewed: A New “Omics” Platform Technology for Systems Biology and Implications for Natural Products Research. J. Nat. Prod. 2005, 68, 1813–1820.
- Forster, G.; Heuberger, A.; Broeckling, C.; Bauer, J.; Ryan, E. Consumption of Cooked Navy Bean Powders Modulate the Canine Fecal and Urine Metabolome. Curr. Metab. 2015, 3, 90–101.
- O’Gorman, A.; Brennan, L. Metabolomic applications in nutritional research: A perspective. J. Sci. Food Agric. 2015, 95, 2567–2570.
- Gibney, M.J.; Walsh, M.; Brennan, L.; Roche, H.M.; German, B.; Ben van Ommen, B. Metabolomics in human nutrition: Opportunities and challenges. Am. J. Clin. Nutr. 2005, 82, 497–503.
- Nyatanyi, T.; Wilkes, M.; McDermott, H.; Nzietchueng, S.; Gafarasi, I.; Mudakikwa, A.; Kinani, J.F.; Rukelibuga, J.; Omolo, J.; Mupfasoni, D.; et al. Implementing One Health as an integrated approach to health in Rwanda. BMJ Glob. Health 2017, 2, e000121.
- American Veterinary Medical Association. One Health: A New Professional Imperative, One Health Initiative Task Force: Final Report; American Veterinary Medical Association: Schaumburg, IL, USA, 2008; Available online: http://www.avma.org/onehealth/default.asp (accessed on 25 October 2022).
- Gibbs, E.P.J. Emerging zoonotic epidemics in the interconnected global community. Vet. Rec. 2005, 157, 673–679.
- Mwacalimba, K.K.; Green, J. ‘One health’ and development priorities in resource-constrained countries: Policy lessons from avian and pandemic influenza preparedness in Zambia. Health Policy Plan. 2014, 30, 215–222.
- Baker, M. Metabolomics: From small molecules to big ideas. Nat. Methods 2011, 8, 117–121.
- Gika, H.G.; Theodoridis, G.A.; Plumb, R.S.; Wilson, I.D. Current practice of liquid chromatography–mass spectrometry in metabolomics and metabonomics. J. Pharm. Biomed. Anal. 2014, 87, 12–25.
- Sebedio, J.L.; Brennan, L. Metabolomics as a Tool in Nutrition Research; Elsevier Ltd.: London, UK, 2015; 269p.
- Astarita, G.; Langridge, J. An Emerging Role for Metabolomics in Nutrition Science. Lifestyle Genom. 2013, 6, 181–200.
- Patti, G.J.; Yanes, O.; Siuzdak, G. Metabolomics: The apogee of the omics trilogy. Nat. Rev. Mol. Cell Biol. 2012, 13, 263–269.
- Roux, A.; Lison, D.; Junot, C.; Heilier, J.-F. Applications of liquid chromatography coupled to mass spectrometry-based metabolomics in clinical chemistry and toxicology: A review. Clin. Biochem. 2011, 44, 119–135.
- Afman, L.; Müller, M. Nutrigenomics: From Molecular Nutrition to Prevention of Disease. J. Am. Diet. Assoc. 2006, 106, 569–576.
- Trujillo, E.; Davis, C.; Milner, J. Nutrigenomics, Proteomics, Metabolomics, and the Practice of Dietetics. J. Am. Diet. Assoc. 2006, 106, 403–413.
- Merched, A.J.; Chan, L. Nutrigenetics and Nutrigenomics of Atherosclerosis. Curr. Atheroscler. Rep. 2013, 15, 328.
- Ganesh, V.; Hettiarachchy, N.S. Nutriproteomics: A promising tool to link diet and diseases in nutritional research. Biochim. Biophys. Acta BBA Proteins Proteom. 2012, 1824, 1107–1117.
- Micha, R.; Michas, G.; Lajous, M.; Mozaffarian, D. Processing of meats and cardiovascular risk: Time to focus on preservatives. BMC Med. 2013, 11, 136.
- Simopoulos, A.P. Dietary Omega-3 Fatty Acid Deficiency and High Fructose Intake in the Development of Metabolic Syndrome, Brain Metabolic Abnormalities, and Non-Alcoholic Fatty Liver Disease. Nutrients 2013, 5, 2901–2923.
- LeMieux, M.; Al-Jawadi, A.; Wang, S.; Moustaid-Moussa, N. Metabolic Profiling in Nutrition and Metabolic Disorders. Adv. Nutr. Int. Rev. J. 2013, 4, 548–550.
- O’Gorman, A.; Brennan, L. The role of metabolomics in determination of new dietary biomarkers. Proc. Nutr. Soc. 2017, 76, 295–302.
- Cade, J.E. Measuring diet in the 21st century: Use of new technologies. Proc. Nutr. Soc. 2017, 76, 276–282.
- Stella, C.; Beckwith-Hall, B.; Cloarec, O.; Holmes, E.; Lindon, J.C.; Powell, J.; van der Ouderaa, F.; Bingham, S.; Cross, A.A.J.; Nicholson, J.K. Susceptibility of Human Metabolic Phenotypes to Dietary Modulation. J. Proteome Res. 2006, 5, 2780–2788.
- Vasil’Ev, A.V.; Sharanova, N.; Kulakova, S.N. Nutrimetabolomics—The new stage of biochemistry of nutrition. The role of nutrilipidomic analysis. Probl. Nutr. 2014, 83, 4–11.
- Odriozola, L.; Corrales, F.J. Discovery of nutritional biomarkers: Future directions based on omics technologies. Int. J. Food Sci. Nutr. 2015, 66, S31–S40.
- Regan, J.A.; Shah, S.H. Obesity Genomics and Metabolomics: A Nexus of Cardiometabolic Risk. Curr. Cardiol. Rep. 2020, 22, 174.
- Wu, Y.; Perng, W.; Peterson, K.E. Precision Nutrition and Childhood Obesity: A Scoping Review. Metabolites 2020, 10, 235.
- Arneth, B.; Arneth, R.; Shams, M. Metabolomics of Type 1 and Type 2 Diabetes. Int. J. Mol. Sci. 2019, 20, 2467.
- Wang, N.; Zhu, F.; Chen, L.; Chen, K. Proteomics, metabolomics and metagenomics for type 2 diabetes and its complications. Life Sci. 2018, 212, 194–202.
- Li, X.S.; Wang, Z.; Cajka, T.; Buffa, J.A.; Nemet, I.; Hurd, A.G.; Gu, X.; Skye, S.M.; Roberts, A.B.; Wu, Y.; et al. Untargeted metabolomics identifies trimethyllysine, a TMAO-producing nutrient precursor, as a predictor of incident cardiovascular disease risk. J. Clin. Investig. 2018, 3, e99096.
- Onuh, J.O.; Aliani, M. Metabolomics profiling in hypertension and blood pressure regulation: A review. Clin. Hypertens. 2020, 26, 23.
- Kumar, A.; Misra, B.B. Challenges and Opportunities in Cancer Metabolomics. Proteomics 2019, 19, e1900042.
- Yang, Q.-J.; Zhao, J.-R.; Hao, J.; Li, B.; Huo, Y.; Han, Y.-L.; Wan, L.-L.; Li, J.; Huang, J.; Lu, J.; et al. Serum and urine metabolomics study reveals a distinct diagnostic model for cancer cachexia. J. Cachexia Sarcopenia Muscle 2017, 9, 71–85.
- Perez-Cornago, A.; Brennan, L.; Ibero-Baraibar, I.; Hermsdorff, H.H.M.; O’gorman, A.; Zulet, M.A.; Martínez, J.A. Metabolomics identifies changes in fatty acid and amino acid profiles in serum of overweight older adults following a weight loss intervention. J. Physiol. Biochem. 2014, 70, 593–602.
- Menni, C.; Fauman, E.; Erte, I.; Perry, J.R.; Kastenmüller, G.; Shin, S.-Y.; Petersen, A.-K.; Hyde, C.; Psatha, M.; Ward, K.J.; et al. Biomarkers for Type 2 Diabetes and Impaired Fasting Glucose Using a Nontargeted Metabolomics Approach. Diabetes 2013, 62, 4270–4276.
- Barber, M.N.; Risis, S.; Yang, C.; Meikle, P.J.; Staples, M.; Febbraio, M.A.; Bruce, C.R. Plasma Lysophosphatidylcholine Levels Are Reduced in Obesity and Type 2 Diabetes. PLoS ONE 2012, 7, e41456.
- Adams, S.H. Emerging Perspectives on Essential Amino Acid Metabolism in Obesity and the Insulin-Resistant State. Adv. Nutr. Int. Rev. J. 2011, 2, 445–456.
- Basu, S.; Millett, C. Social Epidemiology of Hypertension in Middle-Income Countries. Hypertension 2013, 62, 18–26.
- Roberts, L.D.; Gerszten, R.E. Toward New Biomarkers of Cardiometabolic Diseases. Cell Metab. 2013, 18, 43–50.
- Magnusson, M.; Lewis, G.D.; Ericson, U.; Orho-Melander, M.; Hedblad, B.; Engström, G.; Östling, G.; Clish, C.; Wang, T.J.; Gerszten, R.E.; et al. A diabetes-predictive amino acid score and future cardiovascular disease. Eur. Heart J. 2012, 34, 1982–1989.
- O’Connell, T.M. The Complex Role of Branched Chain Amino Acids in Diabetes and Cancer. Metabolites 2013, 3, 931–945.
- Asemi, Z.; Samimi, M.; Tabassi, Z.; Esmaillzadeh, A. The effect of DASH diet on pregnancy outcomes in gestational diabetes: A randomized controlled clinical trial. Eur. J. Clin. Nutr. 2014, 68, 490–495.
- Schwingshackl, L.; Hoffmann, G. Adherence to Mediterranean diet and risk of cancer: A systematic review and meta-analysis of observational studies. Int. J. Cancer 2014, 135, 1884–1897.
- Andersen, M.-B.S.; Rinnan, Å.; Manach, C.; Poulsen, S.K.; Pujos-Guillot, E.; Larsen, T.M.; Astrup, A.; Dragsted, L.O. Untargeted Metabolomics as a Screening Tool for Estimating Compliance to a Dietary Pattern. J. Proteome Res. 2014, 13, 1405–1418.
- Wang, Z.; Tang, W.H.W.; Buffa, J.A.; Fu, X.; Britt, E.B.; Koeth, R.A.; Levison, B.S.; Fan, Y.; Wu, Y.; Hazen, S.L. Prognostic value of choline and betaine depends on intestinal microbiota-generated metabolite trimethylamine-N-oxide. Eur. Heart J. 2014, 35, 904–910.
- Dragsted, L.O. Biomarkers of meat intake and the application of nutrigenomics. Meat Sci. 2010, 84, 301–307.
- Baldrick, F.R.; Woodside, J.V.; Elborn, J.S.; Young, I.S.; McKinley, M.C. Biomarkers of Fruit and Vegetable Intake in Human Intervention Studies: A Systematic Review. Crit. Rev. Food Sci. Nutr. 2011, 51, 795–815.
- Marklund, M.; Landberg, R.; Andersson, A.; Åman, P.; Kamal-Eldin, A. Alkylresorcinol metabolites in urine correlate with the intake of whole grains and cereal fibre in free-living Swedish adults. Br. J. Nutr. 2012, 109, 129–136.
- Silva, V.; Barazzoni, R.; Singer, P. Biomarkers of Fish Oil Omega-3 Polyunsaturated Fatty Acids Intake in Humans. Nutr. Clin. Pract. 2013, 29, 63–72.
- Woodside, J.V.; Draper, J.; Lloyd, A.; McKinley, M. Use of biomarkers to assess fruit and vegetable intake. Proc. Nutr. Soc. 2017, 76, 308–315.
- Andersen, M.-B.S.; Kristensen, M.; Manach, C.; Pujos-Guillot, E.; Poulsen, S.K.; Larsen, T.M.; Astrup, A.; Dragsted, L. Discovery and validation of urinary exposure markers for different plant foods by untargeted metabolomics. Anal. Bioanal. Chem. 2014, 406, 1829–1844.
- Heinzmann, S.S.; Brown, I.J.; Chan, Q.; Bictash, M.; Dumas, M.-E.; Kochhar, S.; Stamler, J.; Holmes, E.; Elliott, P.; Nicholson, J.K. Metabolic profiling strategy for discovery of nutritional biomarkers: Proline betaine as a marker of citrus consumption. Am. J. Clin. Nutr. 2010, 92, 436–443.
- Lloyd, A.J.; Beckmann, M.; Favé, G.; Mathers, J.C.; Draper, J. Proline betaine and its biotransformation products in fasting urine samples are potential biomarkers of habitual citrus fruit consumption. Br. J. Nutr. 2011, 106, 812–824.
- May, D.H.; Navarro, S.L.; Ruczinski, I.; Hogan, J.; Ogata, Y.; Schwarz, Y.; Levy, L.; Holzman, T.; McIntosh, M.W.; Lampe, J.W. Metabolomic profiling of urine: Response to a randomised, controlled feeding study of select fruits and vegetables, and application to an observational study. Br. J. Nutr. 2013, 110, 1760–1770.
- Pujos-Guillot, E.; Hubert, J.; Martin, J.-F.; Lyan, B.; Quintana, M.; Claude, S.; Chabanas, B.; Rothwell, J.A.; Bennetau-Pelissero, C.; Scalbert, A.; et al. Mass Spectrometry-based Metabolomics for the Discovery of Biomarkers of Fruit and Vegetable Intake: Citrus Fruit as a Case Study. J. Proteome Res. 2013, 12, 1645–1659.
- O’Sullivan, A.; Gibney, M.J.; Brennan, L. Dietary intake patterns are reflected in metabolomic profiles: Potential role in dietary assessment studies. Am. J. Clin. Nutr. 2011, 93, 314–321.
- Nicholson, J.K.; Holmes, E.; Wilson, I.D. Gut microorganisms, mammalian metabolism and personalized health care. Nat. Rev. Genet. 2005, 3, 431–438.
- Xie, G.; Zhang, S.; Zheng, X.; Jia, W. Metabolomics approaches for characterizing metabolic interactions between host and its commensal microbes. Electrophoresis 2013, 34, 2787–2798.
- Lamichhane, S.; Sen, P.; Dickens, A.M.; Orešič, M.; Bertram, H.C. Gut metabolome meets microbiome: A methodological perspective to understand the relationship between host and microbe. Methods 2018, 149, 3–12.
- Geier, B.; Sogin, E.M.; Michellod, D.; Janda, M.; Kompauer, M.; Spengler, B.; Dubilier, N.; Liebeke, M. Spatial metabolomics of in situ host–microbe interactions at the micrometre scale. Nat. Microbiol. 2020, 5, 498–510.
- Feng, Y.-L.; Cao, G.; Chen, D.-Q.; Vaziri, N.D.; Chen, L.; Zhang, J.; Wang, M.; Guo, Y.; Zhao, Y.-Y. Microbiome–metabolomics reveals gut microbiota associated with glycine-conjugated metabolites and polyamine metabolism in chronic kidney disease. Cell. Mol. Life Sci. 2019, 76, 4961–4978.
- Han, J.; Antunes, L.C.M.; Finlay, B.B.; Borchers, C.H. Metabolomics: Towards understanding host–microbe interactions. Future Microbiol. 2010, 5, 153–161.
- Rezzi, S.; Ramadan, Z.; Martin, F.-P.J.; Fay, L.B.; van Bladeren, P.; Lindon, J.C.; Nicholson, J.K.; Kochhar, S. Human Metabolic Phenotypes Link Directly to Specific Dietary Preferences in Healthy Individuals. J. Proteome Res. 2007, 6, 4469–4477.
- Herder, C.; Kowall, B.; Tabak, A.G.; Rathmann, W. The potential of novel biomarkers to improve risk prediction of type 2 diabetes. Diabetologia 2013, 57, 16–29.
- Feng, Z.; Ding, C.; Li, W.; Wang, D.; Cui, D. Applications of metabolomics in the research of soybean plant under abiotic stress. Food Chem. 2019, 310, 125914.
- Zhu, G.; Wang, S.; Huang, Z.; Zhang, S.; Liao, Q.; Zhang, C.; Lin, T.; Qin, M.; Peng, M.; Yang, C.; et al. Rewiring of the Fruit Metabolome in Tomato Breeding. Cell 2018, 172, 249–261.e12.
- Tian, J.; Wang, Y.Z.; Yan, S.X.; Sun, S.; Jia, J.J.; Hu, X.X. Metabolomics technology and its applications in agricultural animal and plant research. Hereditas 2020, 42, 452–465.
- Goldansaz, S.A.; Guo, A.C.; Sajed, T.; Steele, M.A.; Plastow, G.S.; Wishart, D.S. Livestock metabolomics and the livestock metabolome: A systematic review. PLoS ONE 2017, 12, e0177675.
- Emara, S.; Amer, S.; Ali, A.; Abouleila, Y.; Oga, A.; Masujima, T. Single-Cell Metabolomics. Adv. Exp. Med. Biol. 2017, 965, 323–343.
- Manach, C.; Hubert, J.; Llorach, R.; Scalbert, A. The complex links between dietary phytochemicals and human health deciphered by metabolomics. Mol. Nutr. Food Res. 2009, 53, 1303–1315.
- Palmnäs, M.; Brunius, C.; Shi, L.; Rostgaard-Hansen, A.; Torres, N.E.; González-Domínguez, R.; Zamora-Ros, R.; Ye, Y.L.; Halkjær, J.; Tjønneland, A.; et al. Perspective: Metabotyping—A Potential Personalized Nutrition Strategy for Precision Prevention of Cardiometabolic Disease. Adv. Nutr. Int. Rev. J. 2020, 11, 524–532.
- Braconi, D.; Bernardini, G.; Millucci, L.; Santucci, A. Foodomics for human health: Current status and perspectives. Expert Rev. Proteom. 2017, 15, 153–164.
- Cuadrado-Silva, C.T.; Pozo-Bayón, M.; Osorio, C. Targeted Metabolomic Analysis of Polyphenols with Antioxidant Activity in Sour Guava (Psidium friedrichsthalianum Nied.) Fruit. Molecules 2016, 22, 11.
- Calumpang, C.L.F.; Saigo, T.; Watanabe, M.; Tohge, T. Cross-Species Comparison of Fruit-Metabolomics to Elucidate Metabolic Regulation of Fruit Polyphenolics Among Solanaceous Crops. Metabolites 2020, 10, 209.
- Dixon, R.A.; Gang, D.R.; Charlton, A.J.; Fiehn, O.; Kuiper, H.A.; Reynolds, T.L.; Tjeerdema, R.S.; Jeffery, E.; German, J.B.; Ridley, W.P.; et al. Applications of Metabolomics in Agriculture. J. Agric. Food Chem. 2006, 54, 8984–8994.
- Simpson, M.J.; McKelvie, J.R. Environmental metabolomics: New insights into earthworm ecotoxicity and contaminant bioavailability in soil. Anal. Bioanal. Chem. 2009, 394, 137–149.
- Bundy, J.; Davey, M.P.; Viant, M.R. Environmental metabolomics: A critical review and future perspectives. Metabolomics 2008, 5, 3–21.
- Sumner, L.W.; Mendes, P.; Dixon, R.A. Plant metabolomics: Large-scale phytochemistry in the functional genomics era. Phytochemistry 2003, 62, 817–836.
- Levandi, T.; Leon, C.; Kaljurand, M.; Garcia-Cañas, V.; Cifuentes, A. Capillary Electrophoresis Time-of-Flight Mass Spectrometry for Comparative Metabolomics of Transgenic versus Conventional Maize. Anal. Chem. 2008, 80, 6329–6335.
- García-Villalba, R.; León, C.; Dinelli, G.; Segura-Carretero, A.; Fernández-Gutiérrez, A.; Garcia-Cañas, V.; Cifuentes, A. Comparative metabolomic study of transgenic versus conventional soybean using capillary electrophoresis–time-of-flight mass spectrometry. J. Chromatogr. A 2008, 1195, 164–173.
- Jung, Y.; Lee, J.; Kwon, J.; Lee, K.-S.; Ryu, D.H.; Hwang, G.-S. Discrimination of the Geographical Origin of Beef by 1H NMR-Based Metabolomics. J. Agric. Food Chem. 2010, 58, 10458–10466.
- Cajka, T.; Riddellova, K.; Tomaniova, M.; Hajslova, J. Ambient mass spectrometry employing a DART ion source for metabolomic fingerprinting/profiling: A powerful tool for beer origin recognition. Metabolomics 2010, 7, 500–508.
- Capanoglu, E.; Beekwilder, J.; Boyacioglu, D.; Hall, R.; de Vos, R. Changes in Antioxidant and Metabolite Profiles during Production of Tomato Paste. J. Agric. Food Chem. 2008, 56, 964–973.
- Boudonck, K.J.; Mitchell, M.; Wulff, J.; Ryals, J.A. Characterization of the biochemical variability of bovine milk using metabolomics. Metabolomics 2009, 5, 375–386.
- Castellano, P.; Ibarreche, M.P.; Massani, M.B.; Fontana, C.; Vignolo, G.M. Strategies for Pathogen Biocontrol Using Lactic Acid Bacteria and Their Metabolites: A Focus on Meat Ecosystems and Industrial Environments. Microorganisms 2017, 5, 38.
- Ercolini, D.; Russo, F.; Nasi, A.; Ferranti, P.; Villani, F. Mesophilic and Psychrotrophic Bacteria from Meat and Their Spoilage Potential In Vitro and in Beef. Appl. Environ. Microbiol. 2009, 75, 1990–2001.
- Xu, Y.; Cheung, W.; Winder, C.L.; Goodacre, R. VOC-based metabolic profiling for food spoilage detection with the application to detecting Salmonella typhimurium-contaminated pork. Anal. Bioanal. Chem. 2010, 397, 2439–2449.
- Herrero, M.; Simo, C.; Garcia-Cañas, V.; Ibanez, E.; Cifuentes, A. Foodomics: MS-based strategies in modern food science and nutrition. Mass Spectrom. Rev. 2011, 31, 49–69.
- van Ruth, S.; Alewijn, M.; Rogers, K.; Newton-Smith, E.; Tena, N.; Bollen, M.; Koot, A. Authentication of organic and conventional eggs by carotenoid profiling. Food Chem. 2010, 126, 1299–1305.
- Ceciliani, F.; Roccabianca, P.; Giudice, C.; Lecchi, C. Application of post-genomic techniques in dog cancer research. Mol. Biosyst. 2016, 12, 2665–2679.
- de Godoy, M.R.; Hervera, M.; Swanson, K.S.; Fahey, G.C. Innovations in Canine and Feline Nutrition: Technologies for Food and Nutrition Assessment. Annu. Rev. Anim. Biosci. 2016, 4, 311–333.
- Honneffer, J.B.; Steiner, J.M.; Lidbury, J.A.; Suchodolski, J.S. Variation of the microbiota and metabolome along the canine gastrointestinal tract. Metabolomics 2017, 13, 26.
- Zhang, S.; Gowda, G.N.; Asiago, V.; Shanaiah, N.; Barbas, C.; Raftery, D. Correlative and quantitative 1H NMR-based metabolomics reveals specific metabolic pathway disturbances in diabetic rats. Anal. Biochem. 2008, 383, 76–84.
- O’kell, A.L.; Garrett, T.J.; Wasserfall, C.; Atkinson, M.A. Untargeted metabolomic analysis in naturally occurring canine diabetes mellitus identifies similarities to human Type 1 Diabetes. Sci. Rep. 2017, 7, 9467.
- O’kell, A.L.; Garrett, T.J.; Wasserfall, C.; Atkinson, M.A. Untargeted metabolomic analysis in non-fasted diabetic dogs by UHPLC–HRMS. Metabolomics 2019, 15, 15.
- Zhang, A.; Sun, H.; Wang, P.; Han, Y.; Wang, X. Recent and potential developments of biofluid analyses in metabolomics. J. Proteom. 2012, 75, 1079–1088.
- Tamai, R.; Furuya, M.; Hatoya, S.; Akiyoshi, H.; Yamamoto, R.; Komori, Y.; Yokoi, S.-I.; Tani, K.; Hirano, Y.; Komori, M.; et al. Profiling of Serum Metabolites in Canine Lymphoma Using Gas Chromatography Mass Spectrometry. J. Vet. Med. Sci. 2014, 76, 1513–1518.
- Minamoto, Y.; Otoni, C.C.; Steelman, S.M.; Büyükleblebici, O.; Steiner, J.M.; Jergens, A.E.; Suchodolski, J.S. Alteration of the fecal microbiota and serum metabolite profiles in dogs with idiopathic inflammatory bowel disease. Gut Microbes 2015, 6, 33–47.
- Abdullah, M.; Kornegay, J.N.; Honcoop, A.; Parry, T.L.; Balog-Alvarez, C.J.; O’neal, S.K.; Bain, J.R.; Muehlbauer, M.J.; Newgard, C.B.; Patterson, C.; et al. Non-Targeted Metabolomics Analysis of Golden Retriever Muscular Dystrophy-Affected Muscles Reveals Alterations in Arginine and Proline Metabolism, and Elevations in Glutamic and Oleic Acid In Vivo. Metabolites 2017, 7, 38.
- Martini, F.M.; De Bellesini, A.B.; Miolo, A.; Del Coco, L.; Fanizzi, F.P.; Crovace, A. Combining a joint health supplement with tibial plateau leveling osteotomy in dogs with cranial cruciate ligament rupture. An exploratory controlled trial. Int. J. Vet. Sci. Med. 2017, 5, 105–112.
- Wang, Y.; Lawler, D.; Larson, B.; Ramadan, Z.; Kochhar, S.; Holmes, E.; Nicholson, J.K. Metabonomic Investigations of Aging and Caloric Restriction in a Life-Long Dog Study. J. Proteome Res. 2007, 6, 1846–1854.
- Hall, J.A.; Brockman, J.A.; Jewell, D.E. Dietary fish oil alters the lysophospholipid metabolomic profile and decreases urinary 11-dehydro thromboxane B2 concentration in healthy Beagles. Vet. Microbiol. 2011, 144, 355–365.
- Allaway, D.; Kamlage, B.; Gilham, M.S.; Hewson-Hughes, A.K.; Wiemer, J.C.; Colyer, A. Efects of dietary glucose supplementation on the fasted plasma metabolome in cats and dogs. Metabolomics 2013, 9, 1096–1108.
- Schmidt, M.; Unterer, S.; Suchodolski, J.S.; Honneffer, J.B.; Guard, B.C.; Lidbury, J.A.; Steiner, J.M.; Fritz, J.; Kölle, P. The fecal microbiome and metabolome differs between dogs fed Bones and Raw Food (BARF) diets and dogs fed commercial diets. PLoS ONE 2018, 13, e0201279.
- Yan, G.; Zhao, Y.; Deng, P.; Lv, L.; Wang, Y.; Bu, Q.; Liu, B.; Hu, C.; Zhuo, Y.; Yang, X.; et al. Investigation of toxicological effects of Shuanghuanglian injection in Beagle dogs by metabonomic and traditional approaches. Exp. Biol. Med. 2010, 235, 1356–1364.
- Zhang, H.; Patrone, L.; Kozlosky, J.; Tomlinson, L.; Cosma, G.; Horvath, J. Pooled Sample Strategy in Conjunction with High-Resolution Liquid Chromatography−Mass Spectrometry-Based Background Subtraction to Identify Toxicological Markers in Dogs Treated with Ibipinabant. Anal. Chem. 2010, 82, 3834–3839.
- Osaki, T.; Azuma, K.; Kurozumi, S.; Takamori, Y.; Tsuka, T.; Imagawa, T.; Okamoto, Y.; Minami, S. Metabolomic Analyses of Blood Plasma after Oral Administration of D-Glucosamine Hydrochloride to Dogs. Mar. Drugs 2012, 10, 1873–1882.
- Kararli, T.T. Comparison of the gastrointestinal anatomy, physiology, and biochemistry of humans and commonly used laboratory animals. Biopharm. Drug Dispos. 1995, 16, 351–380.
- Kang, C.-I.; Kim, S.-H.; Park, W.B.; Lee, K.-D.; Kim, H.-B.; Kim, E.-C.; Oh, M.-D.; Choe, K.-W. Bloodstream Infections Caused by Antibiotic-Resistant Gram-Negative Bacilli: Risk Factors for Mortality and Impact of Inappropriate Initial Antimicrobial Therapy on Outcome. Antimicrob. Agents Chemother. 2005, 49, 760–766.
- German, J.B.; Roberts, M.-A.; Watkins, S.M. Personal Metabolomics as a Next Generation Nutritional Assessment. J. Nutr. 2003, 133, 4260–4266.
- Suhre, K. Genetics Meets Metabolomics—From Experiment to Systems Biology; Springer: Berlin/Heidelberg, Germany, 2012; 318p.
- Puurunen, J.; Tiira, K.; Lehtonen, M.; Hanhineva, K.; Lohi, H. Non-targeted metabolite profiling reveals changes in oxidative stress, tryptophan and lipid metabolisms in fearful dogs. Behav. Brain Funct. 2016, 12, 7.
- González-Ramírez, M.T. Compatibility between Humans and Their Dogs: Benefits for Both. Animals 2019, 9, 674.
- De Souza, A.I.; Cardin, S.; Wait, R.; Chung, Y.-L.; Vijayakumar, M.; Maguy, A.; Camm, A.J.; Nattel, S. Proteomic and metabolomic analysis of atrial profibrillatory remodelling in congestive heart failure. J. Mol. Cell. Cardiol. 2010, 49, 851–863.
- Shibayama, J.; Taylor, T.G.; Venable, P.W.; Rhodes, N.L.; Gil, R.B.; Warren, M.; Wende, A.R.; Abel, E.D.; Cox, J.; Spitzer, K.W.; et al. Metabolic Determinants of Electrical Failure in Ex-Vivo Canine Model of Cardiac Arrest: Evidence for the Protective Role of Inorganic Pyrophosphate. PLoS ONE 2013, 8, e57821.
- Shibayama, J.; Yuzyuk, T.N.; Cox, J.; Makaju, A.; Miller, M.; Lichter, J.; Li, H.; Leavy, J.D.; Franklin, S.; Zaitsev, A.V. Metabolic Remodeling in Moderate Synchronous versus Dyssynchronous Pacing-Induced Heart Failure: Integrated Metabolomics and Proteomics Study. PLoS ONE 2015, 10, e0118974.
- Li, Q.; Freeman, L.M.; Rush, J.E.; Huggins, G.S.; Kennedy, A.D.; Labuda, J.A.; Laflamme, D.P.; Hannah, S.S. Veterinary Medicine and Multi-Omics Research for Future Nutrition Targets: Metabolomics and Transcriptomics of the Common Degenerative Mitral Valve Disease in Dogs. OMICS J. Integr. Biol. 2015, 19, 461–470.
- Whitfeld, P.D.; Noble, P.J.M.; Major, H.; Beynon, R.J.; Burrow, R.; Freeman, A.I. Metabolomics as a diagnostic tool for hepatology: Validation in a naturally occurring canine model. Metabolomics 2005, 1, 215–225.
- Kafsack, B.F.; Llinás, M. Eating at the Table of Another: Metabolomics of Host-Parasite Interactions. Cell Host Microbe 2010, 7, 90–99.
- Kloehn, J.; Blume, M.; Cobbold, S.; Saunders, E.; Dagley, M.; McConville, M. Using metabolomics to dissect host–parasite interactions. Curr. Opin. Microbiol. 2016, 32, 59–65.
- Puurunen, J.; Sulkama, S.; Tiira, K.; Araujo, C.; Lehtonen, M.; Hanhineva, K.; Lohi, H. A non-targeted metabolite profiling pilot study suggests that tryptophan and lipid metabolisms are linked with ADHD-like behaviours in dogs. Behav. Brain Funct. 2016, 12, 27.
- Colyer, A.; Gilham, M.S.; Kamlage, B.; Rein, D.; Allaway, D. Identification of intra- and inter-individual metabolite variation in plasma metabolite profiles of cats and dogs. Br. J. Nutr. 2011, 106, S146–S149.
- Viant, M.R.; Ludwig, C.; Rhodes, S.; Günther, U.L.; Allaway, D. Validation of a urine metabolome fingerprint in dog for phenotypic classification. Metabolomics 2007, 3, 453–463.
- Lloyd, A.J.; Beckmann, M.; Tailliart, K.; Brown, W.; Draper, J.; Allaway, D. Characterisation of the main drivers of intra- and inter- breed variability in the plasma metabolome of dogs. Metabolomics 2016, 12, 72.
- Lloyd, A.J.; Beckmann, M.; Wilson, T.; Tailliart, K.; Allaway, D.; Draper, J. Ultra high performance liquid chromatography–high resolution mass spectrometry plasma lipidomics can distinguish between canine breeds despite uncontrolled environmental variability and non-standardized diets. Metabolomics 2017, 13, 15.
- Osaki, T.; Kurozumi, S.; Sato, K.; Terashi, T.; Azuma, K.; Murahata, Y.; Tsuka, T.; Ito, N.; Imagawa, T.; Minami, S.; et al. Metabolomic Analysis of Blood Plasma after Oral Administration of N-acetyl-d-Glucosamine in Dogs. Mar. Drugs 2015, 13, 5007–5015.
- Wagoner, M.P.; Yang, Y.; McDuffie, J.E.; Klapczynski, M.; Buck, W.; Cheatham, L.; Eisinger, D.; Sace, F.; Lynch, K.M.; Sonee, M.; et al. Evaluation of Temporal Changes in Urine-based Metabolomic and Kidney Injury Markers to Detect Compound Induced Acute Kidney Tubular Toxicity in Beagle Dogs. Curr. Top. Med. Chem. 2017, 17, 2767–2780.
- Söbbeler, F.J.; Carrera, I.; Pasloske, K.; Ranasinghe, M.G.; Kircher, P.; Kästner, S.B.R. Effects of isoflurane, sevoflurane, propofol and alfaxalone on brain metabolism in dogs assessed by proton magnetic resonance spectroscopy (1H MRS). BMC Vet. Res. 2018, 14, 69.
- Musteata, M.; Nicolescu, A.; Solcan, G.; Deleanu, C. The 1H NMR Profile of Healthy Dog Cerebrospinal Fluid. PLoS ONE 2013, 8, e81192.
- Hasegawa, T.; Sumita, M.; Horitani, Y.; Tamai, R.; Tanaka, K.; Komori, M.; Takenaka, S. Gas Chromatography-Mass Spectrometry-Based Metabolic Profiling of Cerebrospinal Fluid from Epileptic Dogs. J. Vet. Med. Sci. 2014, 76, 517–522.
- Yanibada, B.; Hohenester, U.; Pétéra, M.; Canlet, C.; Durand, S.; Jourdan, F.; Boccard, J.; Martin, C.; Eugène, M.; Morgavi, D.P.; et al. Inhibition of enteric methanogenesis in dairy cows induces changes in plasma metabolome highlighting metabolic shifts and potential markers of emission. Sci. Rep. 2020, 10, 15591.
- Zhou, M.; Jing, J.H.; Mao, R.H.; Guo, J.; Wang, Z.P. Applications of metabonomics in animal genetics and breeding. Hereditas 2019, 41, 111–124.
- Long, J.A. The ‘omics’ revolution: Use of genomic, transcriptomic, proteomic and metabolomic tools to predict male reproductive traits that impact fertility in livestock and poultry. Anim. Reprod. Sci. 2020, 220, 106354.
- Bertram, H.C.; Jakobsen, L.M.A. Nutrimetabolomics: Integrating metabolomics in nutrition to disentangle intake of animal-based foods. Metabolomics 2018, 14, 34.
- Pugliese, A.; Gruppillo, A.; Di Pietro, S.; Pietro, S. Clinical Nutrition in Gerontology: Chronic Renal Disorders of the Dog and Cat. Vet. Res. Commun. 2005, 29, 57–63.
- Moore, R.E.; Kirwan, J.; Doherty, M.K.; Whitfield, P.D. Biomarker Discovery in Animal Health and Disease: The Application of Post-Genomic Technologies. Biomark. Insights 2007, 2, 185–196.
- Whitfield, P.; German, A.; Noble, P.-J.M. Metabolomics: An emerging post-genomic tool for nutrition. Br. J. Nutr. 2004, 92, 549–555.
- Zeisel, S.H. Nutrigenomics and metabolomics will change clinical nutrition and public health practice: Insights from studies on dietary requirements for choline. Am. J. Clin. Nutr. 2007, 86, 542–548.
- One Health Initiative. Available online: http://www.onehealthinitiative.com/ (accessed on 8 December 2020).
- Kahn, L.H.; Kaplan, B.; Steele, J.H. Confronting zoonoses through closer collaboration between medicine and veterinary medicine (as ‘one medicine’). Vet. Ital. 2010, 43, 5–19.
- Cecaro, M. Food Borne Illness and One Medicine Approach. J. Mass Commun. J. 2014, s1, 002.
- Mersha, C.; Tewodros, F. One Health One Medicine One World: Co-joint of Animal and Human Medicine with Perspectives, A review. Vet. World 2012, 5, 238–243.
- Zinsstag, J.; Schelling, E.; Schelling, E.; Waltner-Toews, D.; Tanner, M. One Health: The Theory and Practice of Integrated Health Approaches; CABI: São Paulo, Brazil, 2015; 478p.
- Frank, D. One world, one health, one medicine. Can. Vet. J. 2008, 49, 1063–1065.
- Karesh, W.B.; Dobson, A.; Lloyd-Smith, J.O.; Lubroth, J.; Dixon, M.A.; Bennett, M.; Aldrich, S.; Harrington, T.; Formenty, P.; Loh, E.H.; et al. Ecology of zoonoses: Natural and unnatural histories. Lancet 2012, 380, 1936–1945.
- Dubey, J.P.; Hotea, I.; Olariu, T.R.; Jones, J.L.; Dărăbuş, G. Epidemiological review of toxoplasmosis in humans and animals in Romania. Parasitology 2013, 141, 311–325.
- World Health Organization. Noncommunicable Diseases. Fact Sheet 355. 2014. Available online: http://www.who.int/mediacentre/factsheets/fs355/en (accessed on 16 December 2020).
- Chandler, M.; Cunningham, S.; Lund, E.M.; Khanna, C.; Naramore, R.; Patel, A.; Day, M.J. Obesity and Associated Comorbidities in People and Companion Animals: A One Health Perspective. J. Comp. Pathol. 2017, 156, 296–309.
- Muñoz-Prieto, A.; Nielsen, L.R.; Dąbrowski, R.; Bjørnvad, C.R.; Söder, J.; Lamy, E.; Monkeviciene, I.; Ljubić, B.B.; Vasiu, I.; Savic, S.; et al. European dog owner perceptions of obesity and factors associated with human and canine obesity. Sci. Rep. 2018, 8, 13353.
- Delicano, R.A.; Hammar, U.; Egenvall, A.; Westgarth, C.; Mubanga, M.; Byberg, L.; Fall, T.; Kennedy, B. The shared risk of diabetes between dog and cat owners and their pets: Register based cohort study. BMJ 2020, 371, m4337.
- Thibault, M.; Houlbrèque, F.; Lorrain, A.; Vidal, E. Seabirds: Sentinels beyond the oceans. Science 2019, 366, 813.
- Schmidt, P.L. Companion Animals as Sentinels for Public Health. Veter. Clin. N. Am. Small Anim. Pract. 2009, 39, 241–250.
- Reif, J.S. Animal Sentinels for Environmental and Public Health. Public Health Rep. 2011, 126, 50–57.
- National Research Council (US) Committee on Animals as Monitors of Environmental Hazards. Animals as Sentinels of Environmental Health Hazards; National Academies Press (US): Washington, DC, USA, 2000.
- Scorpio, D.G. Do cats serve as good sentinels for Bartonella species infection risk in people and animals? Vet. Rec. 2017, 180, 322–324.
- Ellis, K.H. One Health Initiative will Unite Veterinary, Human Medicine. Infectious Disease News. February 2008. Available online: https://www.healio.com/news/infectious-disease/20120225/one-health-initiative-will-unite-veterinary-human-medicine (accessed on 10 May 2022).
- Mobasheri, A. COVID-19, Companion Animals, Comparative Medicine, and One Health. Front. Vet. Sci. 2020, 7, 522.
- Thamm, D.; Dow, S. How companion animals contribute to the fight against cancer in humans. Vet. Ital. 2010, 45, 111–120.
- Jacob, M.; Lopata, A.L.; Dasouki, M.; Rahman, A.M.A. Metabolomics toward personalized medicine. Mass Spectrom. Rev. 2019, 38, 221–238.
- Sandøe, P.; Palmer, C.; Corr, S.; Astrup, A.; Bjornvad, C.R. Canine and feline obesity: A One Health perspective. Vet. Rec. 2014, 175, 610–616.
- Chijiiwa, H.; Takagi, S.; Arahori, M.; Hori, Y.; Saito, A.; Kuroshima, H.; Fujita, K. Dogs and cats prioritize human action: Choosing a now-empty instead of a still-baited container. Anim. Cogn. 2020, 24, 65–73.
- Scotch, M.; Odofin, L.; Rabinowitz, P. Linkages between animal and human health sentinel data. BMC Vet. Res. 2009, 5, 15.
- Harding, J.C. Genomics, animal models, and emerging diseases: Relevance to One Health and food security. Genome 2015, 58, 499–502.
- European Commission—Food Safety Overwiew. Available online: https://ec.europa.eu/food/overview_en (accessed on 15 December 2021).
- Brennan, L. Metabolomics: A Powerful Tool to Enrich our Understanding of the Impact of Food on Health. Mol. Nutr. Food Res. 2019, 63, e1870087.
- FAO. Rome Declaration on World Food Security and World Food Summit Plan of Action. 2000. Available online: http://www.fao.org/docrep/003/W3613E/W3613E00.htm (accessed on 13 December 2020).
- Van Haver, E.; Alink, G.; Barlow, S.; Cockburn, A.; Flachowsky, G.; Knudsen, I.; Kuiper, H.; Massin, D.P.; Pascal, G.; Peijnenburg, A.D.; et al. Safety and nutritional assessment of GM plants and derived food and feed: The role of animal feeding trials. Food Chem. Toxicol. 2008, 46, S2–S70.
- Sun, W.; Chen, Z.; Hong, J.; Shi, J. Promoting Human Nutrition and Health through Plant Metabolomics: Current Status and Challenges. Biology 2020, 10, 20.
- Hemler, E.C.; Hu, F.B. Plant-Based Diets for Personal, Population, and Planetary Health. Adv. Nutr. Int. Rev. J. 2019, 10, S275–S283.
- Gupta, S.M.; Arora, S.; Mirza, N.; Pande, A.; Lata, C.; Puranik, S.; Kumar, J.; Kumar, A. Finger Millet: A “Certain” Crop for an “Uncertain” Future and a Solution to Food Insecurity and Hidden Hunger under Stressful Environments. Front. Plant Sci. 2017, 8, 643.
- Alberti, G. Noncommunicable diseases: Tomorrow’s pandemics. Bull. World Health Organ. 2001, 79, 907.
- Beran, D.; Pedersen, H.B.; Robertson, J. Noncommunicable diseases, access to essential medicines and universal health coverage. Glob. Health Action 2019, 12, 1670014.
- Gibbons, H.; O’gorman, A.; Brennan, L. Metabolomics as a tool in nutritional research. Curr. Opin. Infect. Dis. 2015, 26, 30–34.
More
Information
Subjects:
Veterinary Sciences
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
687
Revisions:
2 times
(View History)
Update Date:
06 Jul 2023
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No