Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Jiangxia Zheng | -- | 2493 | 2023-05-16 03:36:29 | | | |
| 2 | Jessie Wu | Meta information modification | 2493 | 2023-05-16 05:25:16 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Zhang, X.; Li, J.; Chen, S.; Yang, N.; Zheng, J. Avian Sex Reversal. Encyclopedia. Available online: https://encyclopedia.pub/entry/44338 (accessed on 23 July 2026).
Zhang X, Li J, Chen S, Yang N, Zheng J. Avian Sex Reversal. Encyclopedia. Available at: https://encyclopedia.pub/entry/44338. Accessed July 23, 2026.
Zhang, Xiuan, Jianbo Li, Sirui Chen, Ning Yang, Jiangxia Zheng. "Avian Sex Reversal" Encyclopedia, https://encyclopedia.pub/entry/44338 (accessed July 23, 2026).
Zhang, X., Li, J., Chen, S., Yang, N., & Zheng, J. (2023, May 16). Avian Sex Reversal. In Encyclopedia. https://encyclopedia.pub/entry/44338
Zhang, Xiuan, et al. "Avian Sex Reversal." Encyclopedia. Web. 16 May, 2023.
Copy Citation
Sex determination and differentiation are processes by which a bipotential gonad adopts either a testicular or ovarian cell fate, and secondary sexual characteristics adopt either male or female developmental patterns. In birds, although genetic factors control the sex determination program, sex differentiation is sensitive to hormones, which can induce sex reversal when disturbed. Although these sex-reversed birds can form phenotypes opposite to their genotypes, none can experience complete sex reversal or produce offspring under natural conditions. Promising evidence indicates that the incomplete sex reversal is associated with cell autonomous sex identity (CASI) of avian cells, which is controlled by genetic factors.
chicken
sex determination and differentiation
sex reversal
poultry
1. Introduction
Birds are important agricultural species and long-standing development model animals [1][2]. For a long time, the global poultry industry has sought effective methods to control the sex of hatchlings to achieve different purposes. In the layer industry, for example, only female chickens are required, and day-old male hatchlings are cruelly culled, which seriously increases ethical and legal concerns [3][4][5][6][7]. Deciphering the mechanisms of avian sex determination and differentiation can contribute to achieving sex control in chickens and avoiding such issues. Furthermore, owing to the high accessibility of chicken embryos, which can be genetically manipulated in vitro, they are ideal models for gonadal research [8][9][10][11]. Given the conservation of gonadal morphogenesis, studies on chicken embryos have shed light on the developmental patterns of the human reproductive system and the causes of sex disorders [12]. Thus, understanding the cell biology and genetics of gonadal formation during avian embryogenesis is crucial for improving our knowledge in diverse areas, from livestock breeding to human sex development.
In birds, the sex of offspring is determined by genetic factors. Male determinant genes activate and maintain the cascade reaction of testicular development, whereas female sex-related pathways are responsible for the proliferation and differentiation of the ovary [13][14][15]. However, avian sex differentiation is regulated by multiple factors, including genetics, epigenetics, and sex hormones [16][17]. Under natural conditions, for example, disease-induced gene mutation and alteration of estrogen levels can significantly divert the direction of sex differentiation, leading to sex reversal [17][18][19]. Although adult sex-reversed chickens form almost identical phenotypes to their opposite genetic sexes, they cannot experience complete sex reversal and produce offspring, which is caused by the cell autonomous sex identity (CASI) of avian cells [16][17][20][21]. Promising investigations indicates that hereditary factors are responsible for avian CASI. The research on the sex development of gynandromorphic birds suggests that the CASI is governed by the cellular sex chromosome combination, and is vital to the maintenance of genetic sex [21]. Until now, researchers have failed to clarify the underlying basis of this biological feature in birds, which needs further research.
2. Occurrence of Vertebrate Sex Reversal
In birds and some lower vertebrates, sex differentiation is often affected by various factors, leading to sex reversal. In chicken flocks, the population’s social structure can influence the sex of individuals, similar to the case of fish [22][23][24][25][26]. For instance, when there is no rooster in a chicken flock, a hen may experience sex reversal to maintain the reproduction of the population [27]. This is an adaptive change that arises during the long-term evolutionary process. However, no further research has been conducted to prove this phenomenon. In addition, diseases are a major cause of sex reversal. Ectopic activation of aromatase and accumulation of estrogens as a result of the henny-feathering trait, an autosomal dominant mutation, can feminize the feathering patterns of male chicken [18][28]. However, this disease-mediated sex reversal might not influence gonadal morphology because aromatase is mainly mis-expressed in extragonadal tissues [18][28].
3. Mechanism of Avian Sex Reversal
Bulk instances of sex reversal in birds are associated with the high sensitivity to alterations in sex hormone levels [16][29]. This biological characteristic of birds is closely related to their evolutionary processes. From the viewpoint of evolution, mechanisms of mammalian sex determination and differentiation evolved from synapsid reptiles, whereas diapsid reptiles gave rise to crocodiles, lizards, and birds 150 million years ago [30][31]. In eutherian mammals, gonadal sex differentiation appears independent of sex steroid hormones and can proceed without steroidogenesis [32][33]. Environmentally insensitive gonads of highly evolved mammalian embryos occur in the maternal womb, which is a potentially dangerous place rich in various hormones (Figure 1) [34]. Hence, the highly evolved placenta and maternal internal pregnancy pattern may have forced eutherians to abandon estrogens as components of the sex-determining cascade. However, sex differentiation of birds occurs in an extra-maternal environment and has characteristics similar to those of lower vertebrates (Figure 1). Therefore, it is likely that they do not have the mechanism to resist the alteration of exogenous estrogen levels, which is usually unlikely to occur under normal hatching conditions. Therefore, the evolutionary position of birds and the biological characteristics of extra-maternal embryogenesis indicate that avian sex differentiation is prone to being affected by environmental hormones.
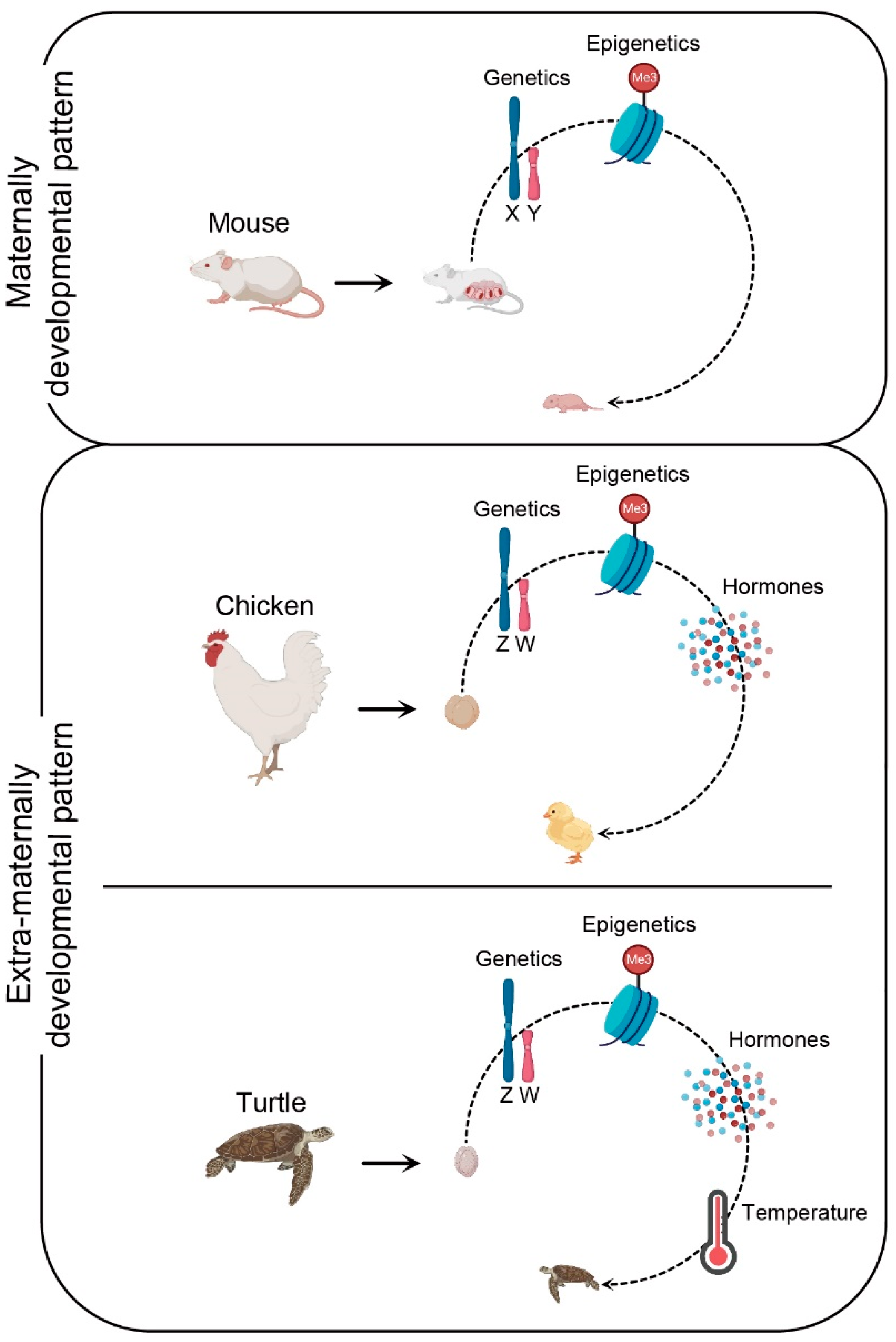
Figure 1. Schematic view of key factors influencing the embryonic sex determination and differentiation in maternal and extra-maternal developmental patterns. In mouse, the process of embryonic sex development occurs in the maternal environment, and is mainly regulated by genetic and epigenetic factors. In chicken, the process of embryonic sex development occurs in the extra-maternal environment, and is governed by genetic and epigenetic factors and sex hormones. Likewise, the process of turtle embryonic sex development occurs in the extra-maternal environment, and is controlled by genetic and epigenetic factors, sex hormones, and temperature.
4. Avian Sex Reversal Induced by Transplant Treatment
Sex-reversed birds can be obtained by altering endogenous hormone secretion through gonadal grafting. Transplantation of E13 (HH39) chicken whole testes into E3 female chicken extra-embryonic coelom can induce gonadal sex reversal [35][36]. Under the stimulation of exogenous testes, the left gonad differentiates into a testis instead of an ovary, and germ cells migrate into sex cords. However, they do not experience a meiotic process [36]. Moreover, grafting E2 female (male) sections of the presumptive mesoderm, which gives rise to gonads, to replace the equivalent tissue of male (female) at the same growth stage, induces the formation of mixed-sex chimeras [21]. Therefore, artificial transplantation of exogenous tissues can affect avian sex differentiation, leading to sex reversal.
5. Avian Male-to-Female Sex Reversal Induced by Estrogens Treatment
In most cases, sex-reversed birds are created by adding estrogens or aromatase inhibitors to manipulate endogenous hormone levels (Figure 2) [16][17]. The administration of estrogen to quail eggs during the first half of embryonic life (before the onset of sex differentiation) can feminize genetic males, characterized by the hyperproliferation of the left gonadal cortex and vacuolized inner medulla [16]. Moreover, injecting a 17a-Ethinylestradiol (EE2) emulsion into E3 quail eggs can demasculinize male individuals, which will lose typical masculine sexual behavior and form asymmetric testes with decreased areas of androgen-dependent cloacal glands after maturity [37]. Recently, research has found that treating chicken eggs with this emulsion can feminize male chickens to different degrees [38][39]. The left gonads of these individuals progressively form female-like cortical regions and fluid-filled medullae from low to high degrees of sex reversal, while the characterized testicular tissues are almost lost [39]. However, the effect of estrogen-mediated male-to-female sex reversal is transient. Some reversed individuals revert to normal male phenotypes after hatching, whereas others can persist in female phenotypes for not more than 1 year [30]. In addition, estrogen-regulated sex reversal is not limited to gonads but is also presented in secondary sexual characteristics. Injection of estradiol into leg muscles of adult male chickens can feminize feathering patterns, including reducing saddle feather length, increasing plumulaceous segments, and altering feathering colors (Figure 2) [40]. Similarly, these changes are not permanent and gradually disappear with decreased estrogen levels in the body. Taken together, the addition of estrogens can significantly affect the differentiation of gonadal structures during embryonic periods and influence the formation of secondary sexual characteristics in adult periods; however, these effects are only maintained for a short time.
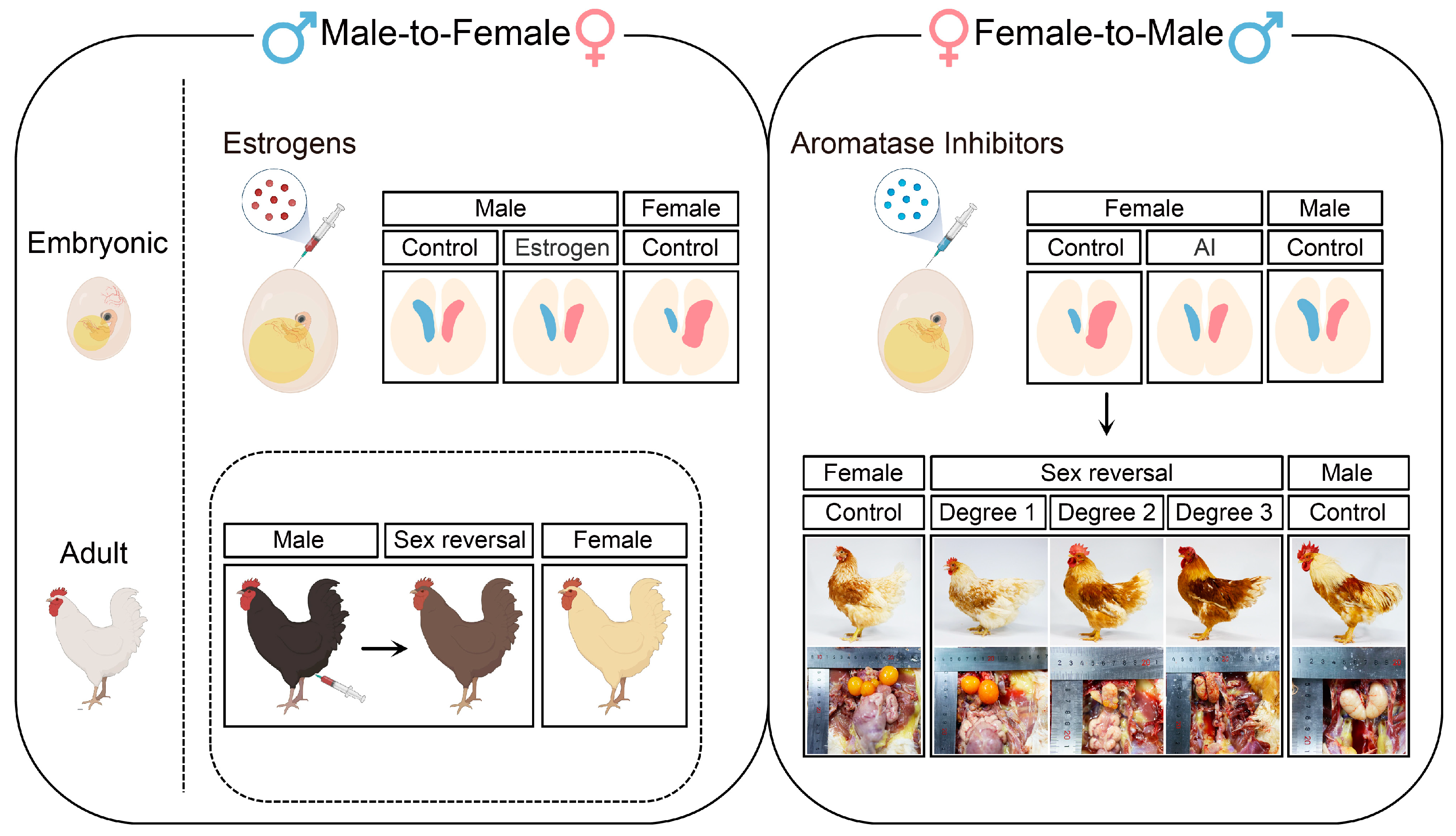
Figure 2. Overview of gonadal phenotypes and secondary sexual characteristics in embryonic and adult sex-reversed chickens created by treatment with either estrogens or aromatase inhibitors. The left panel: Injection of estrogens into chicken eggs can lead to male-to-female sex reversal, characterized by the proliferation of the left gonad and degeneration of the right gonad. During adult period, injection of estrogens into leg muscles of male chicken can induce the feminization of feathering patterns. The right panel: Injection of aromatase inhibitors into chicken eggs can lead to female-to-male sex reversal, characterized by the symmetrical proliferation and differentiation of bilateral gonads. During the adult period, the secondary sexual characteristics and gonadal appearance show different degrees of sex reversal in aromatase inhibitors-treated female chicken.
6. Avian Female-to-Male Sex Reversal Induced by Aromatase Inhibitors Treatment
Injecting aromatase inhibitors, which reduce the production of gonadal estrogens, into avian eggs can induce female-to-male sex reversal (Figure 2) [17][41][42][43]. A previous study found that treating E3 chicken eggs with fadrozole, an aromatase inhibitor, can masculinize bilateral female embryonic gonads, which fail to form stratified cortexes, but develop functional medullae enclosing germ cells at E9.5 [44]. Notably, aromatase inhibitors-treated chickens exhibit varying degrees of sex reversal (Figure 2) [45]. The gonadal cortex is reduced in highly sex-reversed individuals, and germ cells are relocated into developed medullary structures [45]. During the development of sex-reversed female embryos, DMRT1, SOX9, and AMH expression is significantly upregulated in gonadal medullae, whereas FOXL2 and CPY19A1 are downregulated in gonadal cortexes with fewer residues in juxtacortical medullae [44][46][47]. In addition, the primary AMH Receptor, AMH receptor type-II (AMHR2), is upregulated in fadrozole-treated females and co-locates with DMRT1 in Sertoli cells [48]. Moreover, gonadal morphology and inner structures of sex-reversed chickens continue to change after hatching. In 1-day-old sex-reversed chickens (D1), both follicles and tubular structures are visible in the gonads [49]. These tubular structures differentiate into abnormal seminiferous tubules at D11 and form atypical tubules with areas of loose connective tissue at D21 [49]. The left and right gonads grow into small ovotestes in D42 chickens, containing greatly enlarged atypical seminiferous tubules and fewer normal appearing seminiferous tubules [49]. This consecutive alteration is due to the permanent effect of aromatase inhibitors on chicken sex differentiation, which is likely achieved by cell reprogramming during the embryonic period. Research has emphasized that fadrozole-mediated female-to-male sex reversal includes a crucial event, namely, pre-granulosa cells trans-differentiate into undifferentiated PAX2+ supporting cells before forming Sertoli cells, which is lost in estrogens-mediated male-to-female sex reversal [50]. However, current evidences are still unable to clarify the potential mechanism of this phenomenon, which needs to be studied in detail.
The permanent effect of aromatase inhibitors on avian sex differentiation can significantly influence the development of reproductive tissues and the appearance of birds. Adult sex-reversed chickens form almost symmetrical gonads and small testes with regressed oviducts on the left side [17][51][52]. In addition, injecting aromatase inhibitors before sex differentiation during the embryonic period or after sexual maturity in the adult period can induce testosterone production while inhibiting estradiol synthesis in the blood [52][53]. The sex-revered females progressively form masculinized hackles, saddle feathers, combs, and wattles. In contrast, their body weights, especially muscle mass and fat pad weight, remain in line with normal females, suggesting that growth performance, unlike secondary sexual characteristics, may be controlled directly by genetic factors and independent of hormones [40][49][52][53][54]. Notably, under the influence of an aromatase inhibitor, Wolffian ducts in sex-reversed chickens develop into vas deferens, and gonads can produce sperms carrying the W chromosome [55]. Further investigation has shown that W-carrying sperms have oocyte-activating potency and can induce the formation of male and female pronuclei [56][57]. In summary, aromatase inhibitors can significantly disrupt estrogen synthesis and permanently influence the process of sex differentiation, causing the formation of sperm-carrying small testes and masculinized secondary sexual characteristics in adult sex-reversed birds.
7. Avian Sex Reversal and Cell Autonomous Sex Identity
Sex reversal can be induced in birds, but neither does the male bird has the ability to form well-functional ovaries, nor do female birds show normal masculinized courtship behavior and successfully produce fertile sperm after sexual maturity [17][58]. This incomplete sex reversal may be regulated by the CASI of avian somatic cells, which is a cellular inherent sex identity that can help individuals maintain their genetic sex independently of the impact of hormones [21][59]. Current research points out that the CASI is determined by the genetic information, especially that carried by sex chromosomes, and epigenetic factors, such as DNA methylation and histone lysine methylation [21][38]. At early developmental stages when primary gonads have not been induced to differentiation, sexually dimorphic gene expression can be detected in several tissues between male and female birds, indicating that the CASI is widely involved in organogenesis programs [60][61]. Studies of the CASI mainly focus on gynandromorphic birds, which display a significant bilateral asymmetry; one side of the body appears to be male, but the other appears to be female [59]. In gynandromorphic chickens, most cells on the side with a female appearance are of the ZW genotype, and most cells on the other side with a male appearance are of the ZZ genotype [21]. Notably, the gonadal morphologies of these gynandromorphic chickens conform to their cellular compositions, testicular and ovarian appearances are mainly composed of ZZ and ZW genotypic cells, whereas ovotestis is composed of a mixture of both cells [21]. Consistently, male-determining genes, such as DMRT1 and SOX9, are highly expressed in Sertoli cells of the seminiferous cords on the masculinized side, where aromatase is not detectable [62]. However, the gonads of the feminized side show areas of peripheral aromatase expression, together with areas of SOX9-positive and DMRT1-positive seminiferous tubules [62]. Moreover, the concept of CASI has been confirmed in mixed-sex chicken chimeras. Cells from donor females are restricted to the interstitial tissue between the sex cords and are not recruited into functional Sertoli cells in testes. Similarly, donor male cells cannot be recruited to form granulosa cells in ovaries [21]. That is, there is no interaction between GFP+ donor female cells and AMH+ host male cells in the testes, and GFP+ donor male cells and CYP19A1+ host female cells in the ovaries, suggesting that the differentiation of these cells is confirmed by their genetic fates [21]. Notably, epigenetic factors are likely to contribute to maintaining gonadal fate. The chromatin accessibility of DMRT1 in embryonic female-to-male sex-reversed chickens cannot be increased to the normal male level, corresponding to its transcriptional activity [63]. In addition, the pattern of DNA methylation around CYP19A1 in estrogen-mediated sex-reversed chicken gonads fails to be induced to form the same as that in control females, suggesting that epigenetic elements are involved in the establishment of avian gonadal CASI [38][39].
In addition, studies have found that the CASI also affects the growth of avian secondary sexual characteristics [64][65]. In a gynandromorphic finch, although both halves of the brain are exposed to the same hormone environment, the neural song circuit of the male side has a more masculine phenotype than that of the female side [66]. Strikingly, recent research found that DMRT1-knockout male chicken, which had ovaries instead of testes, can form the normal masculinized appearance, featuring large combs and wattles, hackle feathers, and long leg spurs in adult periods [14]. Similarly, in gynandromorphic chickens, the female side forms sex-linked feathering patterns, smaller spurs, and wattles than those on the male side after sexual maturity [62]. These results suggest that avian CASI may be critical in differentiating sex phenotypes.
References
- Tizard, M.L.; Jenkins, K.A.; Cooper, C.A.; Woodcock, M.E.; Challagulla, A.; Doran, T.J. Potential benefits of gene editing for the future of poultry farming. Transgenic Res. 2019, 28, 87–92.
- Zhu, R.; Fogelholm, M.; Jalo, E.; Poppitt, S.D.; Silvestre, M.P.; Møller, G.; Huttunen-Lenz, M.; Stratton, G.; Sundvall, J.; Macdonald, I.A.; et al. Animal-based food choice and associations with long-term weight maintenance and metabolic health after a large and rapid weight loss: The PREVIEW study. Clin. Nutr. 2022, 41, 817–828.
- Doran, T.J.; Morris, K.R.; Wise, T.G.; O’Neil, T.E.; Cooper, C.A.; Jenkins, K.A.; Tizard, M.L.V. Sex selection in layer chickens. Anim. Prod. Sci. 2018, 58, 476.
- Doran, T.J.; Cooper, C.A.; Jenkins, K.A.; Tizard, M.L. Advances in genetic engineering of the avian genome: “Realising the promise”. Transgenic Res. 2016, 25, 307–319.
- Sinclair, M.; Zhang, Y.; Descovich, K.; Phillips, C.J.C. Farm Animal Welfare Science in China-A Bibliometric Review of Chinese Literature. Animals 2020, 10, 540.
- McColl, K.A.; Clarke, B.; Doran, T.J. Role of genetically engineered animals in future food production. Aust. Vet. J. 2013, 91, 113–117.
- Cooper, C.A.; Doran, T.J.; Challagulla, A.; Tizard, M.L.V.; Jenkins, K.A. Innovative approaches to genome editing in avian species. J. Anim. Sci. Biotechnol. 2018, 9, 15.
- Hirst, C.E.; Serralbo, O.; Ayers, K.L.; Roeszler, K.N.; Smith, C.A. Genetic Manipulation of the Avian Urogenital System Using In Ovo Electroporation. Methods Mol. Biol. 2017, 1650, 177–190.
- Yoshino, T.; Murai, H.; Saito, D. Hedgehog-BMP signalling establishes dorsoventral patterning in lateral plate mesoderm to trigger gonadogenesis in chicken embryos. Nat. Commun. 2016, 7, 12561.
- Sekido, R.; Lovell-Badge, R. Mechanisms of gonadal morphogenesis are not conserved between chick and mouse. Dev. Biol. 2007, 302, 132–142.
- Guioli, S.; Zhao, D.; Nandi, S.; Clinton, M.; Lovell-Badge, R. Oestrogen in the chick embryo can induce chromosomally male ZZ left gonad epithelial cells to form an ovarian cortex that can support oogenesis. Development 2020, 147, dev181693.
- Chue, J.; Smith, C.A. Sex determination and sexual differentiation in the avian model. FEBS J. 2011, 278, 1027–1034.
- Smith, C.A.; Roeszler, K.N.; Ohnesorg, T.; Cummins, D.M.; Farlie, P.G.; Doran, T.J.; Sinclair, A.H. The avian Z-linked gene DMRT1 is required for male sex determination in the chicken. Nature 2009, 461, 267–271.
- Ioannidis, J.; Taylor, G.; Zhao, D.; Liu, L.; Idoko-Akoh, A.; Gong, D.; Lovell-Badge, R.; Guioli, S.; McGrew, M.J.; Clinton, M. Primary sex determination in birds depends on DMRT1 dosage, but gonadal sex does not determine adult secondary sex characteristics. Proc. Natl. Acad. Sci. USA 2021, 118, e2020909118.
- Major, A.T.; Ayers, K.; Chue, J.; Roeszler, K.; Smith, C. FOXL2 antagonises the male developmental pathway in embryonic chicken gonads. J. Endocrinol. 2019, 243, 211–228.
- Scheib, D. Effects and role of estrogens in avian gonadal differentiation. Differentiation 1983, 23, S87–S92.
- Elbrecht, A.; Smith, R.G. Aromatase enzyme activity and sex determination in chickens. Science 1992, 255, 467–470.
- George, F.W.; Wilson, J.D. pathogenesis of the henny feathering trait in the Sebright bantam chicken. Increased conversion of androgen to estrogen in skin. J. Clin. Investig. 1980, 66, 57–65.
- Warren, W.C.; Hillier, L.W.; Tomlinson, C.; Minx, P.; Kremitzki, M.; Graves, T.; Markovic, C.; Bouk, N.; Pruitt, K.D.; Thibaud-Nissen, F.; et al. A New Chicken Genome Assembly Provides Insight into Avian Genome Structure. G3 Genes Genomes Genet. 2017, 7, 109–117.
- Vaillant, S.; Dorizzi, M.; Pieau, C.; Richard-Mercier, N. Sex reversal and aromatase in chicken. J. Exp. Zool. 2001, 290, 727–740.
- Zhao, D.; McBride, D.; Nandi, S.; McQueen, H.A.; McGrew, M.J.; Hocking, P.M.; Lewis, P.D.; Sang, H.M.; Clinton, M. Somatic sex identity is cell autonomous in the chicken. Nature 2010, 464, 237–242.
- Godwin, J. Social determination of sex in reef fishes. Semin. Cell. Dev. Biol. 2009, 20, 264–270.
- Lamm, M.S.; Liu, H.; Gemmell, N.J.; Godwin, J.R. The Need for Speed: Neuroendocrine Regulation of Socially-controlled Sex Change. Integr. Comp. Biol. 2015, 55, 307–322.
- Liu, H.; Todd, E.V.; Lokman, P.M.; Lamm, M.S.; Godwin, J.R.; Gemmell, N.J. Sexual plasticity: A fishy tale. Mol. Reprod. Dev. 2017, 84, 171–194.
- Warner, R. Mating Behavior and Hermaphroditism in Coral Reef Fishes. Am. Sci. 1970, 72, 128–136.
- Warner, R.R.; Swearer, S.E. Social Control of Sex Change in the Bluehead Wrasse, Thalassoma bifasciatum (Pisces: Labridae). Biol. Bull. 1991, 181, 199–204.
- Major, A.T.; Smith, C.A. Sex Reversal in Birds. Sex. Dev. 2016, 10, 288–300.
- Matsumine, H.; Herbst, M.A.; Ou, S.H.; Wilson, J.D.; McPhaul, M.J. Aromatase mRNA in the extragonadal tissues of chickens with the henny-feathering trait is derived from a distinctive promoter structure that contains a segment of a retroviral long terminal repeat. Functional organization of the Sebright, Leghorn, and Campine aromatase genes. J. Biol. Chem. 1991, 266, 19900–19907.
- Balthazart, J.; Cornil, C.A.; Charlier, T.D.; Taziaux, M.; Ball, G.F. Estradiol, a key endocrine signal in the sexual differentiation and activation of reproductive behavior in quail. J. Exp. Zool. A Ecol. Genet. Physiol. 2009, 311, 323–345.
- Smith, C.A.; Sinclair, A.H. Sex determination: Insights from the chicken. Bioessays 2004, 26, 120–132.
- Pieau, C. Temperature variation and sex determination in reptiles. BioEssays 1996, 18, 19–26.
- Jost, A. Hormonal factors in the sex differentiation of the mammalian foetus. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1970, 259, 119–130.
- Hu, M.C.; Hsu, N.C.; El Hadj, N.B.; Pai, C.I.; Chu, H.P.; Wang, C.K.; Chung, B.C. Steroid deficiency syndromes in mice with targeted disruption of Cyp11a1. Mol. Endocrinol. 2002, 16, 1943–1950.
- Wolf, U. Reorganization of the sex-determining pathway with the evolution of placentation. Hum. Genet. 1999, 105, 288–292.
- Maraud, R.; Rashedi, M.; Stoll, R. Influence of a temporary embryonic testis graft on the regression of Müllerian ducts in female chick embryo. Development 1982, 67, 81–87.
- Maraud, R.; Vergnaud, O.; Rashedi, M. New insights on the mechanism of testis differentiation from the morphogenesis of experimentally induced testes in genetically female chick embryos. Am. J. Anat. 1990, 188, 429–437.
- Halldin, K.; Berg, C.; Brandt, I.; Brunstrom, B. Sexual behavior in Japanese quail as a test end point for endocrine disruption: Effects of in ovo exposure to ethinylestradiol and diethylstilbestrol. Env. Health Perspect. 1999, 107, 861–866.
- Ellis, H.L.; Shioda, K.; Rosenthal, N.F.; Coser, K.R.; Shioda, T. Masculine epigenetic sex marks of the CYP19A1/aromatase promoter in genetically male chicken embryonic gonads are resistant to estrogen-induced phenotypic sex conversion. Biol. Reprod. 2012, 87, 1–12.
- Shioda, K.; Odajima, J.; Kobayashi, M.; Kobayashi, M.; Cordazzo, B.; Isselbacher, K.J.; Shioda, T. Transcriptomic and Epigenetic Preservation of Genetic Sex Identity in Estrogen-feminized Male Chicken Embryonic Gonads. Endocrinology 2021, 162, bqaa208.
- Widelitz, R.B.; Lin, G.W.; Lai, Y.C.; Mayer, J.A.; Tang, P.C.; Cheng, H.C.; Jiang, T.X.; Chen, C.F.; Chuong, C.M. Morpho-regulation in diverse chicken feather formation: Integrating branching modules and sex hormone-dependent morpho-regulatory modules. Dev. Growth Differ. 2019, 61, 124–138.
- Ellem, S.J.; Risbridger, G.P. Aromatase and regulating the estrogen:androgen ratio in the prostate gland. J. Steroid Biochem. Mol. Biol. 2010, 118, 246–251.
- Nelson, L.R.; Bulun, S.E. Estrogen production and action. J. Am. Acad. Derm. 2001, 45, S116–S124.
- Bulun, S.E.; Lin, Z.; Imir, G.; Amin, S.; Demura, M.; Yilmaz, B.; Martin, R.; Utsunomiya, H.; Thung, S.; Gurates, B.; et al. Regulation of aromatase expression in estrogen-responsive breast and uterine disease: From bench to treatment. Pharm. Rev. 2005, 57, 359–383.
- Hirst, C.E.; Major, A.T.; Ayers, K.L.; Brown, R.J.; Mariette, M.; Sackton, T.B.; Smith, C.A. Sex Reversal and Comparative Data Undermine the W Chromosome and Support Z-linked DMRT1 as the Regulator of Gonadal Sex Differentiation in Birds. Endocrinology 2017, 158, 2970–2987.
- Wartenberg, H.; Lenz, E.; Schweikert, H.U. Sexual differentiation and the germ cell in sex reversed gonads after aromatase inhibition in the chicken embryo. Andrologia 1992, 24, 1–6.
- Smith, C.A.; Katz, M.; Sinclair, A.H. DMRT1 is upregulated in the gonads during female-to-male sex reversal in ZW chicken embryos. Biol. Reprod. 2003, 68, 560–570.
- Testaz, S.; Duband, J.L. Central role of the alpha4beta1 integrin in the coordination of avian truncal neural crest cell adhesion, migration, and survival. Dev. Dyn. 2001, 222, 127–140.
- Cutting, A.D.; Ayers, K.; Davidson, N.; Oshlack, A.; Doran, T.; Sinclair, A.H.; Tizard, M.; Smith, C.A. Identification, expression, and regulation of anti-Mullerian hormone type-II receptor in the embryonic chicken gonad. Biol. Reprod. 2014, 90, 106.
- Burke, W.H.; Henry, M.H. Gonadal development and growth of chickens and turkeys hatched from eggs injected with an aromatase inhibitor. Poult. Sci. 1999, 78, 1019–1033.
- Estermann, M.A.; Smith, C.A. Fadrozole-mediated sex reversal in the embryonic chicken gonad involves a PAX2 positive undifferentiated supporting cell state. bioRxiv 2022.
- Yang, X.; Zheng, J.; Qu, L.; Chen, S.; Li, J.; Xu, G.; Yang, N. Methylation status of cMHM and expression of sex-specific genes in adult sex-reversed female chickens. Sex. Dev. 2011, 5, 147–154.
- Yang, X.; Zheng, J.; Na, R.; Li, J.; Xu, G.; Qu, L.; Yang, N. Degree of sex differentiation of genetic female chicken treated with different doses of an aromatase inhibitor. Sex. Dev. 2008, 2, 309–315.
- Sechman, A.; Rzasa, J.; Paczoska-Eliasiewicz, H. Effect of non-steroidal aromatase inhibitor on blood plasma ovarian steroid and thyroid hormones in laying hen (Gallus domesticus). J. Vet. Med. A Physiol. Pathol. Clin. Med. 2003, 50, 333–338.
- Mohammadrezaei, M.; Toghyani, M.; Gheisari, A.; Toghyani, M.; Eghbalsaied, S. Synergistic effect of fadrozole and insulin-like growth factor-I on female-to-male sex reversal and body weight of broiler chicks. PLoS ONE 2014, 9, e103570.
- Abinawanto; Zhang, C.; Saito, N.; Matsuda, Y.; Shimada, K. Identification of sperm-bearing female-specific chromosome in the sex-reversed chicken. J. Exp. Zool. 1998, 280, 65–72.
- Vaillant, S.; Guemene, D.; Dorizzi, M.; Pieau, C.; Richard-Mercier, N.; Brillard, J.P. Degree of sex reversal as related to plasma steroid levels in genetic female chickens (Gallus domesticus) treated with Fadrozole. Mol. Reprod. Dev. 2003, 65, 420–428.
- Takagi, S.; Tsukada, A.; Saito, N.; Shimada, K. Fertilizing ability of chicken sperm bearing the W chromosome. Poult. Sci. 2007, 86, 731–738.
- Brunstrom, B.; Axelsson, J.; Mattsson, A.; Halldin, K. Effects of estrogens on sex differentiation in Japanese quail and chicken. Gen. Comp. Endocrinol. 2009, 163, 97–103.
- Clinton, M.; Zhao, D.; Nandi, S.; McBride, D. Evidence for avian cell autonomous sex identity (CASI) and implications for the sex-determination process? Chromosome Res. 2012, 20, 177–190.
- Scholz, B.; Kultima, K.; Mattsson, A.; Axelsson, J.; Brunstrom, B.; Halldin, K.; Stigson, M.; Dencker, L. Sex-dependent gene expression in early brain development of chicken embryos. BMC Neurosci. 2006, 7, 12.
- Ayers, K.L.; Davidson, N.M.; Demiyah, D.; Roeszler, K.N.; Grutzner, F.; Sinclair, A.H.; Oshlack, A.; Smith, C.A. RNA sequencing reveals sexually dimorphic gene expression before gonadal differentiation in chicken and allows comprehensive annotation of the W-chromosome. Genome Biol. 2013, 14, R26.
- Morris, K.R.; Hirst, C.E.; Major, A.T.; Ezaz, T.; Ford, M.; Bibby, S.; Doran, T.J.; Smith, C.A. Gonadal and Endocrine Analysis of a Gynandromorphic Chicken. Endocrinology 2018, 159, 3492–3502.
- Zhang, X.; Li, J.; Wang, X.; Jie, Y.; Sun, C.; Zheng, J.; Li, J.; Yang, N.; Chen, S. ATAC-seq and RNA-seq analysis unravel the mechanism of sex differentiation and infertility in sex reversal chicken. Epigenetics Chromatin 2023, 16, 2.
- Briganti, F.; Papeschi, A.; Mugnai, T.; Dessì-Fulgheri, F. Effect of testosterone on male traits and behaviour in juvenile pheasants. Ethol. Ecol. Evol. 1999, 11, 171–178.
- Fennell, M.J.; Scanes, C.G. Inhibition of growth in chickens by testosterone, 5 alpha-dihydrotestosterone, and 19-nortestosterone. Poult. Sci. 1992, 71, 357–366.
- Agate, R.J.; Grisham, W.; Wade, J.; Mann, S.; Wingfield, J.; Schanen, C.; Palotie, A.; Arnold, A.P. Neural, not gonadal, origin of brain sex differences in a gynandromorphic finch. Proc. Natl. Acad. Sci. USA 2003, 100, 4873–4878.
More
Information
Subjects:
Genetics & Heredity
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.7K
Revisions:
2 times
(View History)
Update Date:
16 May 2023
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No