Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Julien Pierrard | -- | 2101 | 2023-05-08 09:38:16 | | | |
| 2 | Conner Chen | + 9 word(s) | 2110 | 2023-05-10 07:56:48 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Pierrard, J.; Van Ooteghem, G.; Van Den Eynde, M. Metastatic Location Matters. Encyclopedia. Available online: https://encyclopedia.pub/entry/43961 (accessed on 05 July 2026).
Pierrard J, Van Ooteghem G, Van Den Eynde M. Metastatic Location Matters. Encyclopedia. Available at: https://encyclopedia.pub/entry/43961. Accessed July 05, 2026.
Pierrard, Julien, Geneviève Van Ooteghem, Marc Van Den Eynde. "Metastatic Location Matters" Encyclopedia, https://encyclopedia.pub/entry/43961 (accessed July 05, 2026).
Pierrard, J., Van Ooteghem, G., & Van Den Eynde, M. (2023, May 08). Metastatic Location Matters. In Encyclopedia. https://encyclopedia.pub/entry/43961
Pierrard, Julien, et al. "Metastatic Location Matters." Encyclopedia. Web. 08 May, 2023.
Copy Citation
The immune microenvironment varies between organs and is influenced by multiple factors, including exposure to external antigens, organ function, and anatomy. Thus, the TIME (tumour immune microenvironment) of metastases differs depending on hosting organs. This variation may account for the different responses to ICIs (immune checkpoint inhibitors) observed in patients with a similar primary tumour in different metastatic locations. Of the most frequent metastatic locations, the presence of secondary lesions in the liver appears to have the most detrimental impact on the prognosis of patients receiving ICIs.
tumour immune microenvironment
radio-immunotherapy combination
liver metastases
1. Liver Metastases
The liver is exposed to many antigens and microbial products that pass through the portal system. These molecules are removed from the blood system by liver cells to prevent an inflammatory reaction [1][2][3]. A wide variety of immune cells exist in the liver sinusoids, including Kuppfer cells, which produce caspase-3 mediated signals to induce T cells’ apoptosis (such as Fas-ligand [FasL] and TNF-receptor apoptosis-inducing ligand). T cell function is further inhibited by immunosuppressive molecules, including PD-L1, IL-10, and transforming growth factor (TGF)-β, which are secreted by liver-specific antigen-presenting cells (APCs, such as Kuppfer cells, hepatic stellate cells, and sinusoidal endothelial cells). In addition, sinusoidal endothelial cells and natural killer (NK) cells stimulate immunosuppressive regulatory T cells (Tregs), which inhibit the action of T cells. However, this state of tolerance favours the establishment and growth of metastases in the liver [1][2][3][4]. Kuppfer cells, neutrophils, and sinusoidal endothelial cells also contribute to the creation of the liver pre-metastatic niche by secreting adhesion molecules that are recognised by cancer cell receptors.
Once established, liver metastases (among frequent metastatic locations, the liver metastases) induce immune modifications that lead to a “systemic immunosuppressive state”. This phenomenon has been inferred from preclinical models that indicate that LM resection decreases systemic immunosuppression [3]. Studies in mice have revealed a reinforcement of FasL-mediated T cell apoptosis by CD11b+ myeloid-derived suppressor cells (MDSCs), which were particularly enriched in the LM TIME [5]. In addition, mice with LM had increased activation of Tregs (CTLA-4, PD-1, and high inducible co-stimulatory molecule [ICOS]), which is known to contribute to immunosuppression (see Figure 1) [6]. A clinical consequence of this immunosuppressive state is poor prognosis for patients with LM, which has been observed in several cancers regardless of systematic treatment [7][8][9][10][11][12][13][14][15][16].
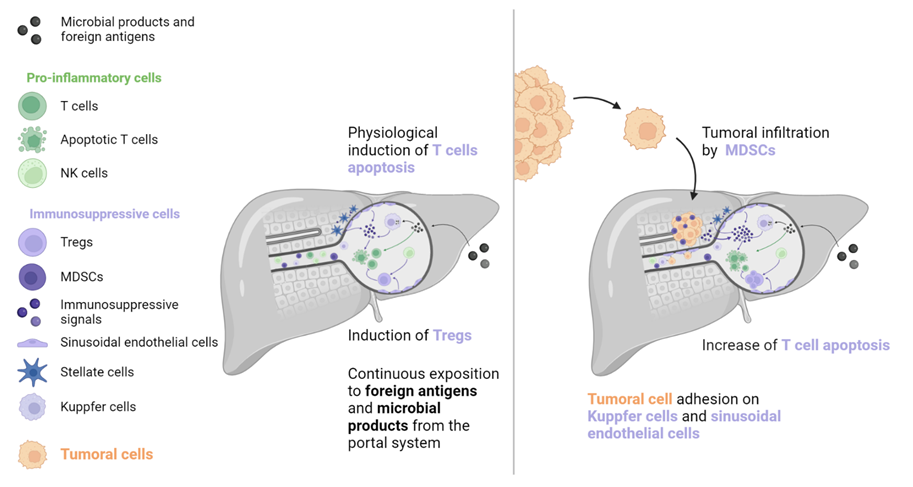
Figure 1. To prevent immune system overactivation, the liver has tolerogenic mechanisms that are primarily mediated by the induction of T cell apoptosis. When a tumour is present, this pathway is strengthened by molecular signals originating from tumoral and TIME cells (such as MDSCs, Kuppfer cells, sinusoidal endothelial cells, and stellate cells). T cell inhibition and depletion may explain the low efficacy of immune checkpoint inhibitors in patients with liver metastases. MDSC: Myeloid-derived suppressive cell, NK: Natural killer, TIME: Tumour immune microenvironment, Treg: Regulator T cell.
Particularly regarding ICIs (immune checkpoint inhibitors), LM-mediated immunosuppression has a significant impact on oncological outcomes [5][7][16][17][18][19][20][21][22][23]. A retrospective study by Tumeh and colleagues included two cohorts of 223 and 113 metastatic melanoma patients, respectively, and one cohort of 165 metastatic NSCLC patients treated with pembrolizumab monotherapy [7]. Compared to lung and brain metastases, LM were found to be an independent poor prognostic factor significantly associated with a shorter progression-free survival (PFS) in all three cohorts. The lower density of CD8+ T cells observed in LM TIME compared to that observed in non-liver metastases may account for this difference. Moreover, non-liver metastases from patients with synchronous LM had a lower CD8+ cells density than those from patients with no LM [7]. In a study by Yu and colleagues, LM were also associated with a reduced objective response rate (ORR) and a reduced overall survival (OS) in melanoma (n = 182) and NSCLC (n = 279) patients treated with ICIs, compared to those of patients with brain or lung metastases. Interestingly, this association was not observed for targeted therapy in melanoma patients or chemotherapy in NSCLC patients [5]. A retrospective report by Osorio and colleagues evaluated metastatic NSCLC (n = 214) and dMMR carcinoma (n = 78) patients treated with PD-1 inhibitors and found that the liver was the most frequent site of progression in both cohorts (NSCLC: 24.0%; dMMR carcinoma: 29.7%). Although the response to PD-1 blockade was uniform among synchronous metastases, progression was found to be heterogeneous. The authors hypothesised that the immunotolerance of the TIME in LM explained why the liver was the first progression site following PD-1 blockade therapy [23].
The negative impact of LM has been observed even in cancers with a poor intrinsic response to ICIs. Wang and colleagues conducted a study of 95 MSS CRC patients treated with PD-(L)1 immunotherapy. Patients without LM had a superior ORR (19.5%) and PFS (4.0 months) compared to those with LM, for whom no objective response or increased PFS (1.5 months) were demonstrated [8][23]. Similarly, a reduced response rate was observed in patients with LM who received anti-PD-1 and anti-CTLA-4 combination therapy. In a retrospective cohort of 140 metastatic melanoma patients who received first-line treatment with dual ICIs, the best outcomes (ORR and PFS) were found in patients with lung metastases, whereas comparatively poor results (ORR, PFS, and OS) were observed in patients with LM [24].
2. Non-Liver Metastases
The lungs, like the liver, are a common metastatic site because they are also exposed to a variety of foreign molecules (e.g., tobacco, traffic pollution, inhaled microorganisms). Thus, tolerance mechanisms are necessary to prevent inappropriate immune reactions. In the lungs, such mechanisms include epithelial cells, airway macrophages, mucociliary action, and commensal microbioma [25]. These mechanisms have the potential to favour a pre-metastatic niche formation, often due to overexpression of the cytokine CCL2, which is derived from the primary tumour and leads resident macrophages and endothelial cells to produce endogenous Toll-like receptor 4 ligands such as S100A8 and SAA3. These effectors facilitate the recruitment of MDSCs and increase vascular permeability, supporting tumoral cell implantation [26]. In addition, γδT cells account for up to 20% of T cells in the lung ecosystem and may promote tumour growth by inducing neoangiogenesis and evasion by restricting the action of APCs, recruiting MDSCs, and secreting immunosuppressive molecules such as IL-4, IL-10, and TGF-β (see Figure 2) [27].

Figure 2. Tolerogenic mechanisms in the lungs involve less T cell apoptosis than those in the liver. In the presence of a tumour, a particular lymphocyte population called γδT cells can have ambiguous effects on tumour development. These cells exert a direct cytotoxic action and an indirect antitumoral action via the stimulation of T cells. However, they can also facilitate tumour development through the secretion of immunosuppressive signals, recruitment of immunosuppressive cells, promotion of neoangiogenesis, and limitation of antigen presentation. DC: Dendritic cell, MDSC: Myeloid-derived suppressive cell, Treg: Regulator T cell.
Bone is another common metastatic site in various solid malignancies, particularly prostate cancer, NSCLC, renal cell carcinoma, and breast cancer [28]. The bone environment is considered immune tolerant due to the abundance of Tregs and MDSCs and the lack of NK and cytotoxic T cells [29][30][31]. Primary tumour cells can generate systemic molecular signals that interact with osteoblasts and osteoclasts to promote angiogenesis, cancer cell adhesion, and bone resorption. The combination of these actions generates a pre-metastatic bone niche (see Figure 3) [30][32].
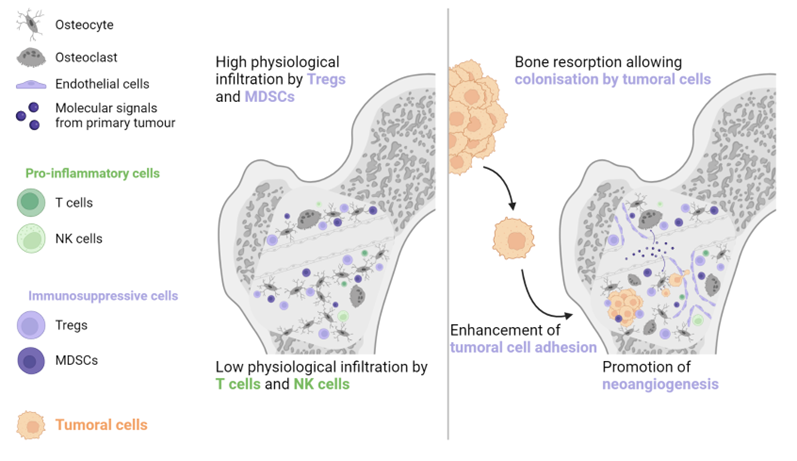
Figure 3. The physiological abundance of Tregs and MDSCs, combined with the paucity of antitumoral immune cells, is the main reason why bones are frequently invaded by metastatic lesions. The primary tumour generates molecular signals, which contribute to the formation of a pre-metastatic niche by increasing bone resorption, stimulating neoangiogenesis, and enhancing tumour cell adhesion. MDSC: Myeloid-derived suppressive cell, NK: Natural killer, Treg: Regulator T cell.
For a long time, the brain was considered an immune-depleted organ due to the presence of immunosuppressive molecules and low T cell infiltration in patients with primary brain tumours [33][34][35]. However, lymphatic vessels have been recently found in regular brain sinuses, transporting immune cells and molecules to the cervical lymph nodes, the first lymphatic relay [34]. A large population of immune cells has also been found in the dura and meninges around the brain, coming through channels directly from the skull bone marrow [36]. Additionally, the microglia, a specialised population of resident macrophages in the brain, is considered the key responder to all types of brain aggressions. It phagocytoses cellular wastes, migrates to the site of disease, secretes pro-inflammatory factors such as IL-1β and tumour-necrosis factor (TNF)-α, and has a direct cytotoxic effect [35][37]. However, prolonged microglial activation induces chronic inflammation and destruction of normal brain cells (astrocytes and neurons), which creates a local environment that favours metastatic invasion (see Figure 4) [35][37][38]. Although patients with brain metastases are usually excluded from pivotal ICIs trials, retrospective data from NSCLC, renal cell carcinoma, and melanoma patients suggest that brain metastases are not a prognostic factor for poor ORR or OS [39][40][41][42].
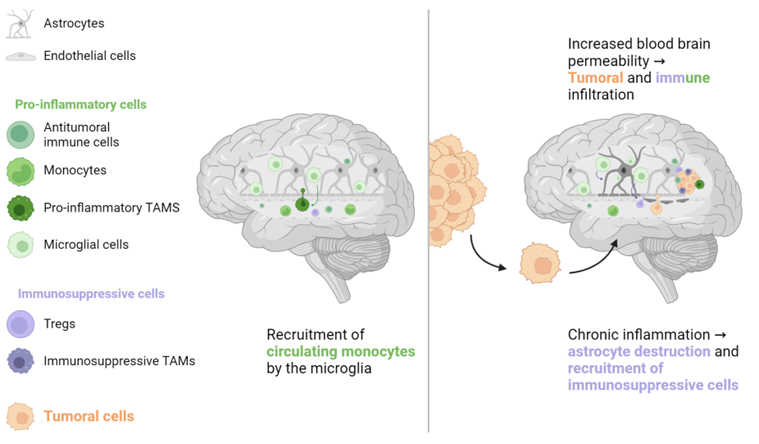
Figure 4. The brain has its own population of macrophages, the microglia. These cells, the main responders to brain injury or aggression, stimulate the recruitment of immune cells, mainly monocytes, from systemic circulation. In the case of chronic inflammation, microglial cells can directly damage normal brain cells such as astrocytes, which are crucial for preserving the blood-brain barrier. Increased blood-brain barrier permeability can promote tumour cell migration and recruitment of immune cells, some of which have an immunosuppressive phenotype that favours tumour cell implantation. TAM: Tumour-associated macrophage, Treg: Regulator T cell.
Studies that have examined ICIs in the metastatic setting do not report lung, bone, or brain metastases as an independent poor prognostic factor for oncological response, in contrast to LM [16][17][18][19][20][21][22][23]. One hypothesis for this discrepancy is that T cells, the main effectors of ICI activity, play a less significant role in normal tissue homeostasis, pre-metastatic niche formation, and metastatic growth in these organs compared to their role in the liver. Therefore, their preservation in non-liver organs would support the maintenance of ICI efficacy, as evidenced by higher ORR in patients with lymph node (LN) metastases [18][19][23]. Indeed, LNs are the sites in which the antitumoral response takes place. LNs are continuously stimulated by tumoral neoantigens, which are processed by dendritic cells to induce CD4+ and CD8+ antitumoral responses favourable to ICI treatment [43][44][45]. However, this situation creates a state of local chronic inflammation that favours the recruitment of immunosuppressive cells, such as MDSCs, tumour-associated macrophages (TAMs), and immature dendritic cells. In turn, this environment is favourable for the implantation and proliferation of cancer cells. This condition is further exacerbated by immunosuppressive signals that are released from the primary tumour, particularly IL-10 and TGF-β (see Figure 5) [44][46][47].
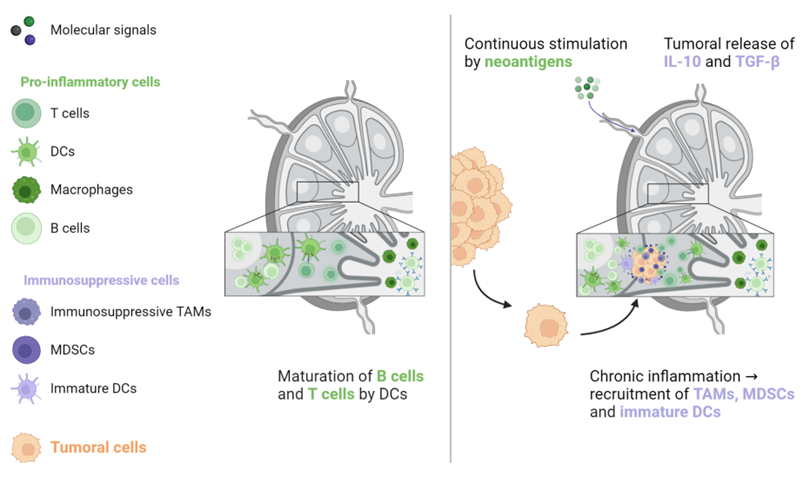
Figure 5. The activation of naive B and T lymphocytes takes place in lymph nodes. When these lymphocytes are continuously triggered by neoantigens from the primary tumour, antitumoral immunity is counterbalanced by immunosuppressive cells, including TAMs, MDSCs, and immature DCs. In addition, the primary tumour generates immunosuppressive signals that promote tumoral cell implantation and proliferation. DC: Dendritic cell, MDSC: Myeloid-derived suppressive cell, TAM: Tumour-associated macrophage, TGF-β: Transforming growth factor-β.
3. Heterogeneity
In addition to metastatic location, TIME heterogeneity is another factor that may affect treatment outcomes, especially in the era of ICIs [5][48][49][50][51][52]. In several malignancies, including NSCLC, melanoma, renal cell carcinoma, and CRC, there is widespread acceptance of the existence of a gene-expression heterogeneity between primary tumours and their metastases, which may impact treatment results. [48][53][54][55]. In many cancers, the TIME in metastases is more immunosuppressive than that of the primary tumours [54][56][57][58][59][60][61][62][63][64][65].
Metastases located in the same organ share similar TIME characteristics, regardless of the primary tumour. García-Mulero and colleagues reported the gene signature of 374 metastases located in four organs (lung, liver, brain, and bone) of various primary tumours. The TIME of lung metastases was the most pro-immunogenic, with a high score for effector cells and a low score for suppressor cells, followed by bone, brain, and liver metastases [66]. The organ affected by metastases may not account for all sources of TIME heterogeneity, but it may partly explain some TIME variations. A study by Angelova and colleagues assessed spatial and temporal TIME heterogeneities in two metastatic CRC patients with a long disease course, due to the availability of multiple tumour samples. Within the same patient, the TIME differed between metastases and between different areas from the same lesion. Additionally, the TIME may vary considerably between metastases that have a parent-child relationship, based on their similar mutational profiles. Globally, the rate of pro-immunogenic mutations seems to decrease in late-onset lesions, which favours the development of immune escape mechanisms and promotes disease progression. However, high immunoscore values, which reflect a high immune infiltration, can also be found in tumour samples collected in later onset of metastatic diseases [67]. Similar TIME heterogeneity has also been reported for metastases arising from the same organ in melanoma and ovarian cancer [68][69].
References
- Crispe, I.N. The liver as a lymphoid organ. Annu. Rev. Immunol. 2009, 27, 147–163.
- Doherty, D.G. Immunity, tolerance and autoimmunity in the liver: A comprehensive review. J. Autoimmun. 2016, 66, 60–75.
- Lee, J.C.; Green, M.D.; Huppert, L.A.; Chow, C.; Pierce, R.H.; Daud, A.I. The Liver–Immunity Nexus and Cancer Immunotherapy. Clin. Cancer Res. 2022, 28, 5–12.
- Ciner, A.T.; Jones, K.; Muschel, R.J.; Brodt, P. The unique immune microenvironment of liver metastases: Challenges and opportunities. Semin. Cancer Biol. 2021, 71, 143–156.
- Yu, J.; Green, M.D.; Li, S.; Sun, Y.; Journey, S.N.; Choi, J.E.; Rizvi, S.M.; Qin, A.; Waninger, J.J.; Lang, X.; et al. Liver metastasis restrains immunotherapy efficacy via macrophage-mediated T cell elimination. Nat. Med. 2021, 27, 152–164.
- Lee, J.C.; Mehdizadeh, S.; Smith, J.; Young, A.; Mufazalov, I.A.; Mowery, C.T.; Daud, A.; Bluestone, J.A. Regulatory T cell control of systemic immunity and immunotherapy response in liver metastasis. Sci. Immunol. 2020, 5, eaba0759.
- Tumeh, P.C.; Hellmann, M.D.; Hamid, O.; Tsai, K.K.; Loo, K.L.; Gubens, M.A.; Rosenblum, M.; Harview, C.L.; Taube, J.M.; Handley, N.; et al. Liver Metastasis and Treatment Outcome with Anti-PD-1 Monoclonal Antibody in Patients with Melanoma and NSCLC. Cancer Immunol. Res. 2017, 5, 417–424.
- Wang, C.; Sandhu, J.; Ouyang, C.; Ye, J.; Lee, P.P.; Fakih, M. Clinical Response to Immunotherapy Targeting Programmed Cell Death Receptor 1/Programmed Cell Death Ligand 1 in Patients With Treatment-Resistant Microsatellite Stable Colorectal Cancer With and Without Liver Metastases. JAMA Netw. Open 2021, 4, e2118416.
- Patanaphan, V.; Salazar, O.M.; Risco, R. Breast cancer: Metastatic patterns and their prognosis. South. Med. J. 1988, 81, 1109–1112.
- Hoang, T.; Dahlberg, S.; Sandler, A.B.; Brahmer, J.R.; Schiller, J.H.; Johnson, D.H. Prognostic models to predict survival in non-small-cell lung cancer patients treated with first-line paclitaxel and carboplatin with or without bevacizumab. J. Thorac. Oncol. 2012, 7, 1361–1368.
- Wu, K.-L.; Tsai, M.-J.; Yang, C.-J.; Chang, W.-A.; Hung, J.-Y.; Yen, C.-J.; Shen, C.-H.; Kuo, T.-Y.; Lee, J.-Y.; Chou, S.-H.; et al. Liver metastasis predicts poorer prognosis in stage IV lung adenocarcinoma patients receiving first-line gefitinib. Lung Cancer 2015, 88, 187–194.
- Ren, Y.; Dai, C.; Zheng, H.; Zhou, F.; She, Y.; Jiang, G.; Fei, K.; Yang, P.; Xie, D.; Chen, C. Prognostic effect of liver metastasis in lung cancer patients with distant metastasis. Oncotarget 2016, 7, 53245–53253.
- Dong, F.; Shen, Y.; Gao, F.; Xu, T.; Wang, X.; Zhang, X.; Zhong, S.; Zhang, M.; Chen, S.; Shen, Z. Prognostic value of site-specific metastases and therapeutic roles of surgery for patients with metastatic bladder cancer: A population-based study. Cancer Manag. Res. 2017, 9, 611–626.
- Pond, G.R.; Sonpavde, G.; de Wit, R.; Eisenberger, M.A.; Tannock, I.F.; Armstrong, A.J. The Prognostic Importance of Metastatic Site in Men with Metastatic Castration-resistant Prostate Cancer. Eur. Urol. 2014, 65, 3–6.
- Fukuoka, S.; Hara, H.; Takahashi, N.; Kojima, T.; Kawazoe, A.; Asayama, M.; Yoshii, T.; Kotani, D.; Tamura, H.; Mikamoto, Y.; et al. Regorafenib Plus Nivolumab in Patients With Advanced Gastric or Colorectal Cancer: An Open-Label, Dose-Escalation, and Dose-Expansion Phase Ib Trial (REGONIVO, EPOC1603). J. Clin. Oncol. 2020, 38, 2053–2061.
- Topalian, S.L.; Hodi, F.S.; Brahmer, J.R.; Gettinger, S.N.; Smith, D.C.; McDermott, D.F.; Powderly, J.D.; Sosman, J.A.; Atkins, M.B.; Leming, P.D.; et al. Five-Year Survival and Correlates Among Patients With Advanced Melanoma, Renal Cell Carcinoma, or Non-Small Cell Lung Cancer Treated With Nivolumab. JAMA Oncol. 2019, 5, 1411–1420.
- Bilen, M.A.; Shabto, J.M.; Martini, D.J.; Liu, Y.; Lewis, C.; Collins, H.; Akce, M.; Kissick, H.; Carthon, B.C.; Shaib, W.L.; et al. Sites of metastasis and association with clinical outcome in advanced stage cancer patients treated with immunotherapy. BMC Cancer 2019, 19, 857.
- Nishino, M.; Ramaiya, N.H.; Chambers, E.S.; Adeni, A.E.; Hatabu, H.; Jänne, P.A.; Hodi, F.S.; Awad, M.M. Immune-related response assessment during PD-1 inhibitor therapy in advanced non-small-cell lung cancer patients. J. Immunother. Cancer 2016, 4, 84.
- Schmid, S.; Diem, S.; Li, Q.; Krapf, M.; Flatz, L.; Leschka, S.; Desbiolles, L.; Klingbiel, D.; Jochum, W.; Früh, M. Organ-specific response to nivolumab in patients with non-small cell lung cancer (NSCLC). Cancer Immunol. Immunother. 2018, 67, 1825–1832.
- Sasaki, A.; Nakamura, Y.; Mishima, S.; Kawazoe, A.; Kuboki, Y.; Bando, H.; Kojima, T.; Doi, T.; Ohtsu, A.; Yoshino, T.; et al. Predictive factors for hyperprogressive disease during nivolumab as anti-PD1 treatment in patients with advanced gastric cancer. Gastric Cancer 2019, 22, 793–802.
- Botticelli, A.; Cirillo, A.; Scagnoli, S.; Cerbelli, B.; Strigari, L.; Cortellini, A.; Pizzuti, L.; Vici, P.; De Galitiis, F.; Di Pietro, F.R.; et al. The Agnostic Role of Site of Metastasis in Predicting Outcomes in Cancer Patients Treated with Immunotherapy. Vaccines 2020, 8, 203.
- Wang, X.; Ji, Q.; Yan, X.; Lian, B.; Si, L.; Chi, Z.; Sheng, X.; Kong, Y.; Mao, L.; Bai, X.; et al. The Impact of Liver Metastasis on Anti-PD-1 Monoclonal Antibody Monotherapy in Advanced Melanoma: Analysis of Five Clinical Studies. Front. Oncol. 2020, 10, 546604.
- Osorio, J.C.; Arbour, K.C.; Le, D.T.; Durham, J.N.; Plodkowski, A.J.; Halpenny, D.F.; Ginsberg, M.S.; Sawan, P.; Crompton, J.G.; Yu, H.A.; et al. Lesion-Level Response Dynamics to Programmed Cell Death Protein (PD-1) Blockade. J. Clin. Oncol. 2019, 37, 3546–3555.
- Pires da Silva, I.; Lo, S.; Quek, C.; Gonzalez, M.; Carlino, M.S.; Long, G.V.; Menzies, A.M. Site-specific response patterns, pseudoprogression, and acquired resistance in patients with melanoma treated with ipilimumab combined with anti–PD-1 therapy. Cancer 2020, 126, 86–97.
- Lloyd, C.M.; Marsland, B.J. Lung Homeostasis: Influence of Age, Microbes, and the Immune System. Immunity 2017, 46, 549–561.
- Maru, Y. The lung metastatic niche. J. Mol. Med. 2015, 93, 1185–1192.
- Cheng, M.; Hu, S. Lung-resident γδ T cells and their roles in lung diseases. Immunology 2017, 151, 375–384.
- Hernandez, R.K.; Wade, S.W.; Reich, A.; Pirolli, M.; Liede, A.; Lyman, G.H. Incidence of bone metastases in patients with solid tumors: Analysis of oncology electronic medical records in the United States. BMC Cancer 2018, 18, 44.
- Fornetti, J.; Welm, A.L.; Stewart, S.A. Understanding the Bone in Cancer Metastasis. J. Bone Miner. Res. 2018, 33, 2099–2113.
- Baschuk, N.; Rautela, J.; Parker, B.S. Bone specific immunity and its impact on metastasis. Bonekey Rep. 2015, 4, 665.
- Zhao, E.; Xu, H.; Wang, L.; Kryczek, I.; Wu, K.; Hu, Y.; Wang, G.; Zou, W. Bone marrow and the control of immunity. Cell. Mol. Immunol. 2012, 9, 11–19.
- Ell, B.; Kang, Y. SnapShot: Bone Metastasis. Cell 2012, 151, 690–690.e1.
- Galea, I.; Bechmann, I.; Perry, V.H. What is immune privilege (not)? Trends Immunol. 2007, 28, 12–18.
- Arvanitis, C.D.; Ferraro, G.B.; Jain, R.K. The blood-brain barrier and blood-tumour barrier in brain tumours and metastases. Nat. Rev. Cancer 2020, 20, 26–41.
- Farber, S.H.; Tsvankin, V.; Narloch, J.L.; Kim, G.J.; Salama, A.K.S.; Vlahovic, G.; Blackwell, K.L.; Kirkpatrick, J.P.; Fecci, P.E. Embracing rejection: Immunologic trends in brain metastasis. Oncoimmunology 2016, 5, e1172153.
- Kisler, K.; Zlokovic, B.V. How the brain regulates its own immune system. Nat. Neurosci. 2022, 25, 532–534.
- Prinz, M.; Jung, S.; Priller, J. Microglia Biology: One Century of Evolving Concepts. Cell 2019, 179, 292–311.
- Wu, S.Y.; Watabe, K. The roles of microglia/macrophages in tumor progression of brain cancer and metastatic disease. Front. Biosci. 2017, 22, 1805–1829.
- Di Giacomo, A.M.; Ascierto, P.A.; Pilla, L.; Santinami, M.; Ferrucci, P.F.; Giannarelli, D.; Marasco, A.; Rivoltini, L.; Simeone, E.; Nicoletti, S.V.; et al. Ipilimumab and fotemustine in patients with advanced melanoma (NIBIT-M1): An open-label, single-arm phase 2 trial. Lancet Oncol. 2012, 13, 879–886.
- Di Giacomo, A.; Ascierto, P.; Queirolo, P.; Pilla, L.; Ridolfi, R.; Santinami, M.; Testori, A.; Simeone, E.; Guidoboni, M.; Maurichi, A.; et al. Three-year follow-up of advanced melanoma patients who received ipilimumab plus fotemustine in the Italian Network for Tumor Biotherapy (NIBIT)-M1 phase II study. Ann. Oncol. 2015, 26, 798–803.
- De Giorgi, U.; Cartenì, G.; Giannarelli, D.; Basso, U.; Galli, L.; Cortesi, E.; Caserta, C.; Pignata, S.; Sabbatini, R.; Bearz, A.; et al. Safety and efficacy of nivolumab for metastatic renal cell carcinoma: Real-world results from an expanded access programme. BJU Int. 2019, 123, 98–105.
- Teixeira Loiola de Alencar, V.; Guedes Camandaroba, M.P.; Pirolli, R.; Fogassa, C.A.Z.; Cordeiro de Lima, V.C. Immunotherapy as Single Treatment for Patients With NSCLC With Brain Metastases: A Systematic Review and Meta-Analysis—The META-L-BRAIN Study. J. Thorac. Oncol. 2021, 16, 1379–1391.
- Girard, J.P.; Moussion, C.; Förster, R. HEVs, lymphatics and homeostatic immune cell trafficking in lymph nodes. Nat. Rev. Immunol. 2012, 12, 762–773.
- Sleeman, J.P. The lymph node pre-metastatic niche. J. Mol. Med. 2015, 93, 1173–1184.
- Lee, H.; Na, K.J.; Choi, H. Differences in Tumor Immune Microenvironment in Metastatic Sites of Breast Cancer. Front. Oncol. 2021, 11, 649004.
- Wallace, A.C.; Hollenberg, N.K. The Transplantability of Tumours by Intravenous and Intralymphatic Routes. Br. J. Cancer 1965, 19, 338–342.
- Cady, B. Regional lymph node metastases, a singular manifestation of the process of clinical metastases in cancer: Contemporary animal research and clinical reports suggest unifying concepts. Cancer Treat. Res. 2007, 135, 185–201.
- Reuben, A.; Spencer, C.N.; Prieto, P.A.; Gopalakrishnan, V.; Reddy, S.M.; Miller, J.P.; Mao, X.; De Macedo, M.P.; Chen, J.; Song, X.; et al. Genomic and immune heterogeneity are associated with differential responses to therapy in melanoma. NPJ Genom. Med. 2017, 2, 10.
- Şenbabaoğlu, Y.; Gejman, R.S.; Winer, A.G.; Liu, M.; Van Allen, E.M.; de Velasco, G.; Miao, D.; Ostrovnaya, I.; Drill, E.; Luna, A.; et al. Tumor immune microenvironment characterization in clear cell renal cell carcinoma identifies prognostic and immunotherapeutically relevant messenger RNA signatures. Genome Biol. 2016, 17, 231.
- Oliver, A.J.; Keam, S.P.; von Scheidt, B.; Zanker, D.J.; Harrison, A.J.; Tantalo, D.G.; Darcy, P.K.; Kershaw, M.H.; Slaney, C.Y. Primary and metastatic breast tumors cross-talk to influence immunotherapy responses. OncoImmunology 2020, 9, 1802979.
- Lin, Q.; Ren, L.; Jian, M.; Xu, P.; Li, J.; Zheng, P.; Feng, Q.; Yang, L.; Ji, M.; Wei, Y.; et al. The mechanism of the premetastatic niche facilitating colorectal cancer liver metastasis generated from myeloid-derived suppressor cells induced by the S1PR1–STAT3 signaling pathway. Cell Death Dis. 2019, 10, 693.
- Remark, R.; Alifano, M.; Cremer, I.; Lupo, A.; Dieu-Nosjean, M.-C.; Riquet, M.; Crozet, L.; Ouakrim, H.; Goc, J.; Cazes, A.; et al. Characteristics and clinical impacts of the immune environments in colorectal and renal cell carcinoma lung metastases: Influence of tumor origin. Clin. Cancer Res. 2013, 19, 4079–4091.
- Gerlinger, M.; Rowan, A.J.; Horswell, S.; Math, M.; Larkin, J.; Endesfelder, D.; Gronroos, E.; Martinez, P.; Matthews, N.; Stewart, A.; et al. Intratumor Heterogeneity and Branched Evolution Revealed by Multiregion Sequencing. N. Engl. J. Med. 2012, 366, 883–892.
- Zhang, Y.; Song, J.; Zhao, Z.; Yang, M.; Chen, M.; Liu, C.; Ji, J.; Zhu, D. Single-cell transcriptome analysis reveals tumor immune microenvironment heterogenicity and granulocytes enrichment in colorectal cancer liver metastases. Cancer Lett. 2020, 470, 84–94.
- Rotow, J.; Bivona, T.G. Understanding and targeting resistance mechanisms in NSCLC. Nat. Rev. Cancer 2017, 17, 637–658.
- Mizuno, R.; Kawada, K.; Itatani, Y.; Ogawa, R.; Kiyasu, Y.; Sakai, Y. The Role of Tumor-Associated Neutrophils in Colorectal Cancer. Int. J. Mol. Sci. 2019, 20, 529.
- Valls, A.F.; Knipper, K.; Giannakouri, E.; Sarachaga, V.; Hinterkopf, S.; Wuehrl, M.; Shen, Y.; Radhakrishnan, P.; Klose, J.; Ulrich, A.; et al. VEGFR1+ Metastasis–Associated Macrophages Contribute to Metastatic Angiogenesis and Influence Colorectal Cancer Patient Outcome. Clin. Cancer Res. 2019, 25, 5674–5685.
- Ogiya, R.; Niikura, N.; Kumaki, N.; Bianchini, G.; Kitano, S.; Iwamoto, T.; Hayashi, N.; Yokoyama, K.; Oshitanai, R.; Terao, M.; et al. Comparison of tumor-infiltrating lymphocytes between primary and metastatic tumors in breast cancer patients. Cancer Sci. 2016, 107, 1730–1735.
- Son, S.-M.; Woo, C.G.; Kim, D.H.; Yun, H.Y.; Kim, H.; Kim, H.K.; Yang, Y.; Kwon, J.; Kwon, M.; Kim, T.-Y.; et al. Distinct tumor immune microenvironments in primary and metastatic lesions in gastric cancer patients. Sci. Rep. 2020, 10, 14293.
- Wang, W.; Ye, L.-F.; Bao, H.; Hu, M.-T.; Han, M.; Tang, H.-M.; Ren, C.; Wu, X.; Shao, Y.; Wang, F.-H.; et al. Heterogeneity and evolution of tumour immune microenvironment in metastatic gastroesophageal adenocarcinoma. Gastric Cancer 2022, 25, 1017–1030.
- Baine, M.K.; Turcu, G.; Zito, C.R.; Adeniran, A.J.; Camp, R.L.; Chen, L.; Kluger, H.M.; Jilaveanu, L.B. Characterization of tumor infiltrating lymphocytes in paired primary and metastatic renal cell carcinoma specimens. Oncotarget 2015, 6, 24990–25002.
- Kudo, Y.; Haymaker, C.; Zhang, J.; Reuben, A.; Duose, D.; Fujimoto, J.; Roy-Chowdhuri, S.; Soto, L.S.; Dejima, H.; Parra, E.; et al. Suppressed immune microenvironment and repertoire in brain metastases from patients with resected non-small-cell lung cancer. Ann. Oncol. 2019, 30, 1521–1530.
- Song, C.W.; Glatstein, E.; Marks, L.B.; Emami, B.; Grimm, J.; Sperduto, P.W.; Kim, M.-S.; Hui, S.; Dusenbery, K.E.; Cho, L.C. Biological Principles of Stereotactic Body Radiation Therapy (SBRT) and Stereotactic Radiation Surgery (SRS): Indirect Cell Death. Int. J. Radiat. Oncol. Biol. Phys. 2021, 110, 21–34.
- Zhou, J.; Gong, Z.; Jia, Q.; Wu, Y.; Yang, Z.-Z.; Zhu, B. Programmed death ligand 1 expression and CD8+ tumor-infiltrating lymphocyte density differences between paired primary and brain metastatic lesions in non-small cell lung cancer. Biochem. Biophys. Res. Commun. 2018, 498, 751–757.
- Schlam, I.; Church, S.E.; Hether, T.D.; Chaldekas, K.; Hudson, B.M.; White, A.M.; Maisonet, E.; Harris, B.T.; Swain, S.M. The tumor immune microenvironment of primary and metastatic HER2—Positive breast cancers utilizing gene expression and spatial proteomic profiling. J. Transl. Med. 2021, 19, 480.
- García-Mulero, S.; Alonso, M.H.; Pardo, J.; Santos, C.; Sanjuan, X.; Salazar, R.; Moreno, V.; Piulats, J.M.; Sanz-Pamplona, R. Lung metastases share common immune features regardless of primary tumor origin. J. Immunother. Cancer 2020, 8, e000491.
- Angelova, M.; Mlecnik, B.; Vasaturo, A.; Bindea, G.; Fredriksen, T.; Lafontaine, L.; Buttard, B.; Morgand, E.; Bruni, D.; Jouret-Mourin, A.; et al. Evolution of Metastases in Space and Time under Immune Selection. Cell 2018, 175, 751–765.e16.
- Jiménez-Sánchez, A.; Memon, D.; Pourpe, S.; Veeraraghavan, H.; Li, Y.; Vargas, H.A.; Gill, M.B.; Park, K.J.; Zivanovic, O.; Konner, J.; et al. Heterogeneous Tumor-Immune Microenvironments among Differentially Growing Metastases in an Ovarian Cancer Patient. Cell 2017, 170, 927–938.e20.
- Ascierto, M.L.; Makohon-Moore, A.; Lipson, E.J.; Taube, J.M.; McMiller, T.L.; Berger, A.E.; Fan, J.; Kaunitz, G.J.; Cottrell, T.R.; Kohutek, Z.A.; et al. Transcriptional Mechanisms of Resistance to Anti–PD-1 Therapy. Clin. Cancer Res. 2017, 23, 3168–3180.
- Galon, J.; Mlecnik, B.; Bindea, G.; Angell, H.K.; Berger, A.; Lagorce, C.; Lugli, A.; Zlobec, I.; Hartmann, A.; Bifulco, C.; et al. Towards the introduction of the ‘Immunoscore’ in the classification of malignant tumours. J. Pathol. 2014, 232, 199–209.
- Mlecnik, B.; Van Den Eynde, M.; Bindea, G.; Church, S.E.; Vasaturo, A.; Fredriksen, T.; Lafontaine, L.; Haicheur, N.; Marliot, F.; Debetancourt, D.; et al. Comprehensive Intrametastatic Immune Quantification and Major Impact of Immunoscore on Survival. J. Natl. Cancer Inst. 2018, 110, 97–108.
- Pagès, F.; Mlecnik, B.; Marliot, F.; Bindea, G.; Ou, F.-S.; Bifulco, C.; Lugli, A.; Zlobec, I.; Rau, T.T.; Berger, M.D.; et al. International validation of the consensus Immunoscore for the classification of colon cancer: A prognostic and accuracy study. Lancet 2018, 391, 2128–2139.
- Van den Eynde, M.; Mlecnik, B.; Bindea, G.; Fredriksen, T.; Church, S.E.; Lafontaine, L.; Haicheur, N.; Marliot, F.; Angelova, M.; Vasaturo, A.; et al. The Link between the Multiverse of Immune Microenvironments in Metastases and the Survival of Colorectal Cancer Patients. Cancer Cell 2018, 34, 1012–1026.e3.
More
Information
Subjects:
Oncology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
519
Revisions:
2 times
(View History)
Update Date:
10 May 2023
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No