Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Oncology
The immune microenvironment varies between organs and is influenced by multiple factors, including exposure to external antigens, organ function, and anatomy. Thus, the TIME of metastases differs depending on hosting organs. This variation may account for the different responses to ICIs observed in patients with a similar primary tumour in different metastatic locations. Of the most frequent metastatic locations, the presence of secondary lesions in the liver appears to have the most detrimental impact on the prognosis of patients receiving ICIs.
- tumour immune microenvironment
- radio-immunotherapy combination
- liver metastases
1. Liver Metastases
The liver is exposed to many antigens and microbial products that pass through the portal system. These molecules are removed from the blood system by liver cells to prevent an inflammatory reaction [16,17,18]. A wide variety of immune cells exist in the liver sinusoids, including Kuppfer cells, which produce caspase-3 mediated signals to induce T cells’ apoptosis (such as Fas-ligand [FasL] and TNF-receptor apoptosis-inducing ligand). T cell function is further inhibited by immunosuppressive molecules, including PD-L1, IL-10, and transforming growth factor (TGF)-β, which are secreted by liver-specific antigen-presenting cells (APCs, such as Kuppfer cells, hepatic stellate cells, and sinusoidal endothelial cells). In addition, sinusoidal endothelial cells and natural killer (NK) cells stimulate immunosuppressive regulatory T cells (Tregs), which inhibit the action of T cells. However, this state of tolerance favours the establishment and growth of metastases in the liver [16,17,18,19]. Kuppfer cells, neutrophils, and sinusoidal endothelial cells also contribute to the creation of the liver pre-metastatic niche by secreting adhesion molecules that are recognised by cancer cell receptors.
Once established, liver metastases (among frequent metastatic locations, the liver metastases) induce immune modifications that lead to a “systemic immunosuppressive state”. This phenomenon has been inferred from preclinical models that indicate that LM resection decreases systemic immunosuppression [18]. Studies in mice have revealed a reinforcement of FasL-mediated T cell apoptosis by CD11b+ myeloid-derived suppressor cells (MDSCs), which were particularly enriched in the LM TIME [20]. In addition, mice with LM had increased activation of Tregs (CTLA-4, PD-1, and high inducible co-stimulatory molecule [ICOS]), which is known to contribute to immunosuppression (see Figure 1) [21]. A clinical consequence of this immunosuppressive state is poor prognosis for patients with LM, which has been observed in several cancers regardless of systematic treatment [8,9,22,23,24,25,26,27,28,29].
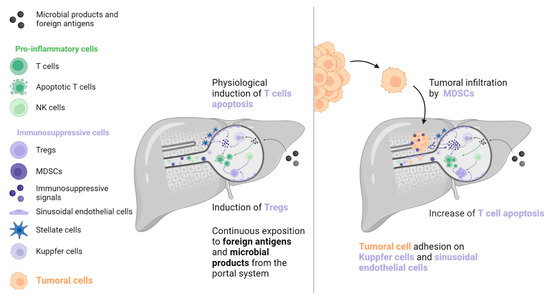
Figure 1. To prevent immune system overactivation, the liver has tolerogenic mechanisms that are primarily mediated by the induction of T cell apoptosis. When a tumour is present, this pathway is strengthened by molecular signals originating from tumoral and TIME cells (such as MDSCs, Kuppfer cells, sinusoidal endothelial cells, and stellate cells). T cell inhibition and depletion may explain the low efficacy of immune checkpoint inhibitors in patients with liver metastases. MDSC: Myeloid-derived suppressive cell, NK: Natural killer, TIME: Tumour immune microenvironment, Treg: Regulator T cell.
Particularly regarding ICIs, LM-mediated immunosuppression has a significant impact on oncological outcomes [8,20,29,30,31,32,33,34,35,36]. A retrospective study by Tumeh and colleagues included two cohorts of 223 and 113 metastatic melanoma patients, respectively, and one cohort of 165 metastatic NSCLC patients treated with pembrolizumab monotherapy [8]. Compared to lung and brain metastases, LM were found to be an independent poor prognostic factor significantly associated with a shorter progression-free survival (PFS) in all three cohorts. The lower density of CD8+ T cells observed in LM TIME compared to that observed in non-liver metastases may account for this difference. Moreover, non-liver metastases from patients with synchronous LM had a lower CD8+ cells density than those from patients with no LM [8]. In a study by Yu and colleagues, LM were also associated with a reduced objective response rate (ORR) and a reduced overall survival (OS) in melanoma (n = 182) and NSCLC (n = 279) patients treated with ICIs, compared to those of patients with brain or lung metastases. Interestingly, this association was not observed for targeted therapy in melanoma patients or chemotherapy in NSCLC patients [20]. A retrospective report by Osorio and colleagues evaluated metastatic NSCLC (n = 214) and dMMR carcinoma (n = 78) patients treated with PD-1 inhibitors and found that the liver was the most frequent site of progression in both cohorts (NSCLC: 24.0%; dMMR carcinoma: 29.7%). Although the response to PD-1 blockade was uniform among synchronous metastases, progression was found to be heterogeneous. The authors hypothesised that the immunotolerance of the TIME in LM explained why the liver was the first progression site following PD-1 blockade therapy [36].
The negative impact of LM has been observed even in cancers with a poor intrinsic response to ICIs. Wang and colleagues conducted a study of 95 MSS CRC patients treated with PD-(L)1 immunotherapy. Patients without LM had a superior ORR (19.5%) and PFS (4.0 months) compared to those with LM, for whom no objective response or increased PFS (1.5 months) were demonstrated [9,36]. Similarly, a reduced response rate was observed in patients with LM who received anti-PD-1 and anti-CTLA-4 combination therapy. In a retrospective cohort of 140 metastatic melanoma patients who received first-line treatment with dual ICIs, the best outcomes (ORR and PFS) were found in patients with lung metastases, whereas comparatively poor results (ORR, PFS, and OS) were observed in patients with LM [37].
2. Non-Liver Metastases
The lungs, like the liver, are a common metastatic site because they are also exposed to a variety of foreign molecules (e.g., tobacco, traffic pollution, inhaled microorganisms). Thus, tolerance mechanisms are necessary to prevent inappropriate immune reactions. In the lungs, such mechanisms include epithelial cells, airway macrophages, mucociliary action, and commensal microbioma [38]. These mechanisms have the potential to favour a pre-metastatic niche formation, often due to overexpression of the cytokine CCL2, which is derived from the primary tumour and leads resident macrophages and endothelial cells to produce endogenous Toll-like receptor 4 ligands such as S100A8 and SAA3. These effectors facilitate the recruitment of MDSCs and increase vascular permeability, supporting tumoral cell implantation [39]. In addition, γδT cells account for up to 20% of T cells in the lung ecosystem and may promote tumour growth by inducing neoangiogenesis and evasion by restricting the action of APCs, recruiting MDSCs, and secreting immunosuppressive molecules such as IL-4, IL-10, and TGF-β (see Figure 2) [40].
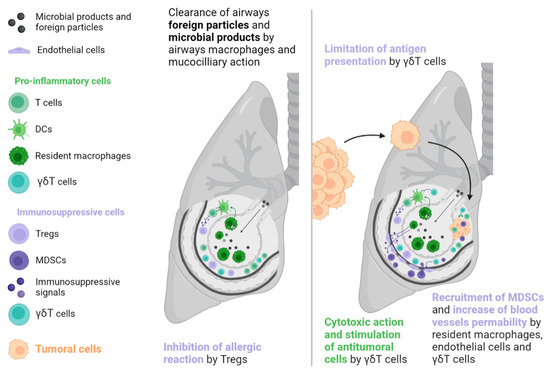
Figure 2. Tolerogenic mechanisms in the lungs involve less T cell apoptosis than those in the liver. In the presence of a tumour, a particular lymphocyte population called γδT cells can have ambiguous effects on tumour development. These cells exert a direct cytotoxic action and an indirect antitumoral action via the stimulation of T cells. However, they can also facilitate tumour development through the secretion of immunosuppressive signals, recruitment of immunosuppressive cells, promotion of neoangiogenesis, and limitation of antigen presentation. DC: Dendritic cell, MDSC: Myeloid-derived suppressive cell, Treg: Regulator T cell.
Bone is another common metastatic site in various solid malignancies, particularly prostate cancer, NSCLC, renal cell carcinoma, and breast cancer [41]. The bone environment is considered immune tolerant due to the abundance of Tregs and MDSCs and the lack of NK and cytotoxic T cells [42,43,44]. Primary tumour cells can generate systemic molecular signals that interact with osteoblasts and osteoclasts to promote angiogenesis, cancer cell adhesion, and bone resorption. The combination of these actions generates a pre-metastatic bone niche (see Figure 3) [43,45].
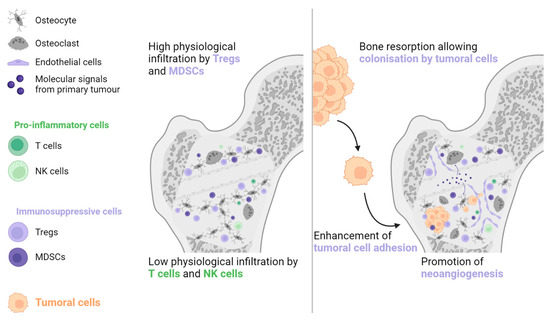
Figure 3. The physiological abundance of Tregs and MDSCs, combined with the paucity of antitumoral immune cells, is the main reason why bones are frequently invaded by metastatic lesions. The primary tumour generates molecular signals, which contribute to the formation of a pre-metastatic niche by increasing bone resorption, stimulating neoangiogenesis, and enhancing tumour cell adhesion. MDSC: Myeloid-derived suppressive cell, NK: Natural killer, Treg: Regulator T cell.
For a long time, the brain was considered an immune-depleted organ due to the presence of immunosuppressive molecules and low T cell infiltration in patients with primary brain tumours [46,47,48]. However, lymphatic vessels have been recently found in regular brain sinuses, transporting immune cells and molecules to the cervical lymph nodes, the first lymphatic relay [47]. A large population of immune cells has also been found in the dura and meninges around the brain, coming through channels directly from the skull bone marrow [49]. Additionally, the microglia, a specialised population of resident macrophages in the brain, is considered the key responder to all types of brain aggressions. It phagocytoses cellular wastes, migrates to the site of disease, secretes pro-inflammatory factors such as IL-1β and tumour-necrosis factor (TNF)-α, and has a direct cytotoxic effect [48,50]. However, prolonged microglial activation induces chronic inflammation and destruction of normal brain cells (astrocytes and neurons), which creates a local environment that favours metastatic invasion (see Figure 4) [48,50,51]. Although patients with brain metastases are usually excluded from pivotal ICIs trials, retrospective data from NSCLC, renal cell carcinoma, and melanoma patients suggest that brain metastases are not a prognostic factor for poor ORR or OS [52,53,54,55].
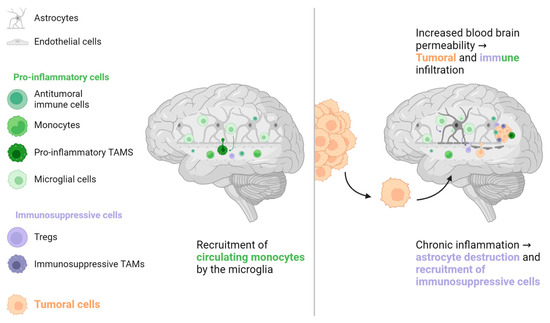
Figure 4. The brain has its own population of macrophages, the microglia. These cells, the main responders to brain injury or aggression, stimulate the recruitment of immune cells, mainly monocytes, from systemic circulation. In the case of chronic inflammation, microglial cells can directly damage normal brain cells such as astrocytes, which are crucial for preserving the blood-brain barrier. Increased blood-brain barrier permeability can promote tumour cell migration and recruitment of immune cells, some of which have an immunosuppressive phenotype that favours tumour cell implantation. TAM: Tumour-associated macrophage, Treg: Regulator T cell.
Studies that have examined ICIs in the metastatic setting do not report lung, bone, or brain metastases as an independent poor prognostic factor for oncological response, in contrast to LM [29,30,31,32,33,34,35,36]. One hypothesis for this discrepancy is that T cells, the main effectors of ICI activity, play a less significant role in normal tissue homeostasis, pre-metastatic niche formation, and metastatic growth in these organs compared to their role in the liver. Therefore, their preservation in non-liver organs would support the maintenance of ICI efficacy, as evidenced by higher ORR in patients with lymph node (LN) metastases [31,32,36]. Indeed, LNs are the sites in which the antitumoral response takes place. LNs are continuously stimulated by tumoral neoantigens, which are processed by dendritic cells to induce CD4+ and CD8+ antitumoral responses favourable to ICI treatment [56,57,58]. However, this situation creates a state of local chronic inflammation that favours the recruitment of immunosuppressive cells, such as MDSCs, tumour-associated macrophages (TAMs), and immature dendritic cells. In turn, this environment is favourable for the implantation and proliferation of cancer cells. This condition is further exacerbated by immunosuppressive signals that are released from the primary tumour, particularly IL-10 and TGF-β (see Figure 5) [57,59,60].
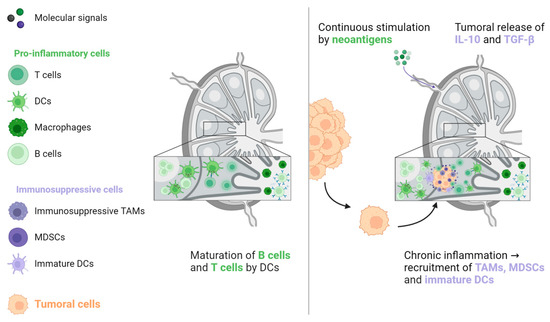
Figure 5. The activation of naive B and T lymphocytes takes place in lymph nodes. When these lymphocytes are continuously triggered by neoantigens from the primary tumour, antitumoral immunity is counterbalanced by immunosuppressive cells, including TAMs, MDSCs, and immature DCs. In addition, the primary tumour generates immunosuppressive signals that promote tumoral cell implantation and proliferation. DC: Dendritic cell, MDSC: Myeloid-derived suppressive cell, TAM: Tumour-associated macrophage, TGF-β: Transforming growth factor-β.
3. Heterogeneity
In addition to metastatic location, TIME heterogeneity is another factor that may affect treatment outcomes, especially in the era of ICIs [20,61,62,63,64,65]. In several malignancies, including NSCLC, melanoma, renal cell carcinoma, and CRC, there is widespread acceptance of the existence of a gene-expression heterogeneity between primary tumours and their metastases, which may impact treatment results. [61,66,67,68]. In many cancers, the TIME in metastases is more immunosuppressive than that of the primary tumours [67,69,70,71,72,73,74,75,76,77,78].
Metastases located in the same organ share similar TIME characteristics, regardless of the primary tumour. García-Mulero and colleagues reported the gene signature of 374 metastases located in four organs (lung, liver, brain, and bone) of various primary tumours. The TIME of lung metastases was the most pro-immunogenic, with a high score for effector cells and a low score for suppressor cells, followed by bone, brain, and liver metastases [79]. The organ affected by metastases may not account for all sources of TIME heterogeneity, but it may partly explain some TIME variations. A study by Angelova and colleagues assessed spatial and temporal TIME heterogeneities in two metastatic CRC patients with a long disease course, due to the availability of multiple tumour samples. Within the same patient, the TIME differed between metastases and between different areas from the same lesion. Additionally, the TIME may vary considerably between metastases that have a parent-child relationship, based on their similar mutational profiles. Globally, the rate of pro-immunogenic mutations seems to decrease in late-onset lesions, which favours the development of immune escape mechanisms and promotes disease progression. However, high immunoscore values, which reflect a high immune infiltration, can also be found in tumour samples collected in later onset of metastatic diseases [80]. Similar TIME heterogeneity has also been reported for metastases arising from the same organ in melanoma and ovarian cancer [81,82].
TIME-based classification systems have been created to account for the effect of TIME heterogeneity on patient prognosis. These systems may be more predictive of patient outcome than the AJCC/UICC TNM classification, as is the “immunoscore” in CRC, for instance [83,84,85,86].
This entry is adapted from the peer-reviewed paper 10.3390/biom13040689
This entry is offline, you can click here to edit this entry!