Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Feng Qian | -- | 2544 | 2023-05-04 10:56:38 | | | |
| 2 | Conner Chen | + 4 word(s) | 2548 | 2023-05-06 03:07:07 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Ma, X.; Yang, T.; Liu, L.; Peng, X.; Qian, F.; Tang, F. Ependymal Dysfunctions in the Pathogenesis of Neurodegenerative Diseases. Encyclopedia. Available online: https://encyclopedia.pub/entry/43750 (accessed on 23 July 2026).
Ma X, Yang T, Liu L, Peng X, Qian F, Tang F. Ependymal Dysfunctions in the Pathogenesis of Neurodegenerative Diseases. Encyclopedia. Available at: https://encyclopedia.pub/entry/43750. Accessed July 23, 2026.
Ma, Xin-Yu, Ting-Ting Yang, Lian Liu, Xiao-Chun Peng, Feng Qian, Feng-Ru Tang. "Ependymal Dysfunctions in the Pathogenesis of Neurodegenerative Diseases" Encyclopedia, https://encyclopedia.pub/entry/43750 (accessed July 23, 2026).
Ma, X., Yang, T., Liu, L., Peng, X., Qian, F., & Tang, F. (2023, May 04). Ependymal Dysfunctions in the Pathogenesis of Neurodegenerative Diseases. In Encyclopedia. https://encyclopedia.pub/entry/43750
Ma, Xin-Yu, et al. "Ependymal Dysfunctions in the Pathogenesis of Neurodegenerative Diseases." Encyclopedia. Web. 04 May, 2023.
Copy Citation
The neuron loss caused by the progressive damage to the nervous system is proposed to be the main pathogenesis of neurodegenerative diseases. Ependyma is a layer of ciliated ependymal cells that participates in the formation of the brain-cerebrospinal fluid barrier (BCB). However, as the protective barrier lining the brain ventricles, the ependyma is extremely vulnerable to cytotoxic and cytolytic immune responses. When the ependyma is damaged, the integrity of BCB is destroyed, and the cerebrospinal fluid (CSF)flow and material exchange is affected, leading to brain microenvironment imbalance, which plays a vital role in the pathogenesis of neurodegenerative diseases.
neurodegenerative diseases
radiation-induced brain injury
ependyma
1. The Physiological Function of Ependyma
Ependymal cells (ECs) line the ventricles and the central canal of the spinal cord [1], forming the brain’s ventricular epithelium and a niche for neural stem cells in the ventricular–subventricular zone (V-SVZ) [2]. These dormant stem cells can be elicited for differentiation and migration after activation [3]. A single-cell transcriptomic study has distinguished the ECs from the ependymal neural stem cells in the V-SVZ and verified no stem cell profile in ECs [4]. In the model of spinal cord injury, ECs showed a limited contribution in astrocytic scar-forming [5]. These findings challenged the hypothesis that the mature EC could function as a neural stem cell. Most mouse immature ECs are derived from radial glial cells around embryonic days 14–16, and then differentiated and matured with cilia formation in neonatal age [6][7]. There are three subtypes of ECs, multi-ciliated, bi-ciliated and mono-ciliated ECs [8]. The bi-ciliated and mono-ciliated ECs indicate the subtypes of tanycytes—the specified ECs. The coordinated beating of those propeller-like motile cilia protruded from ECs into the brain ventricles generates a directional cerebrospinal fluid (CSF) flow, which is essential for various physiological processes [9] (Figure 1A). To organize the formation of CSF, single-layer epithelial cells cover capillaries at the bottom of the lateral ventricle, the top of the third ventricle and the lower part of the fourth ventricle near the inferior medullary velum to form the choroid plexus [10] (Figure 1B). The stroma inside the choroid plexus is a part of the pia mater. As the wall of the ventricular system, ECs and astrocytes from the brain parenchyma form a well-controlled filtration membrane, the BCB, which promotes the bi-directional substance exchange between the BIF and CSF, keeps the brain tissue toxicant-free and in physiological balance [2][3][9][11][12][13] (Figure 1C).
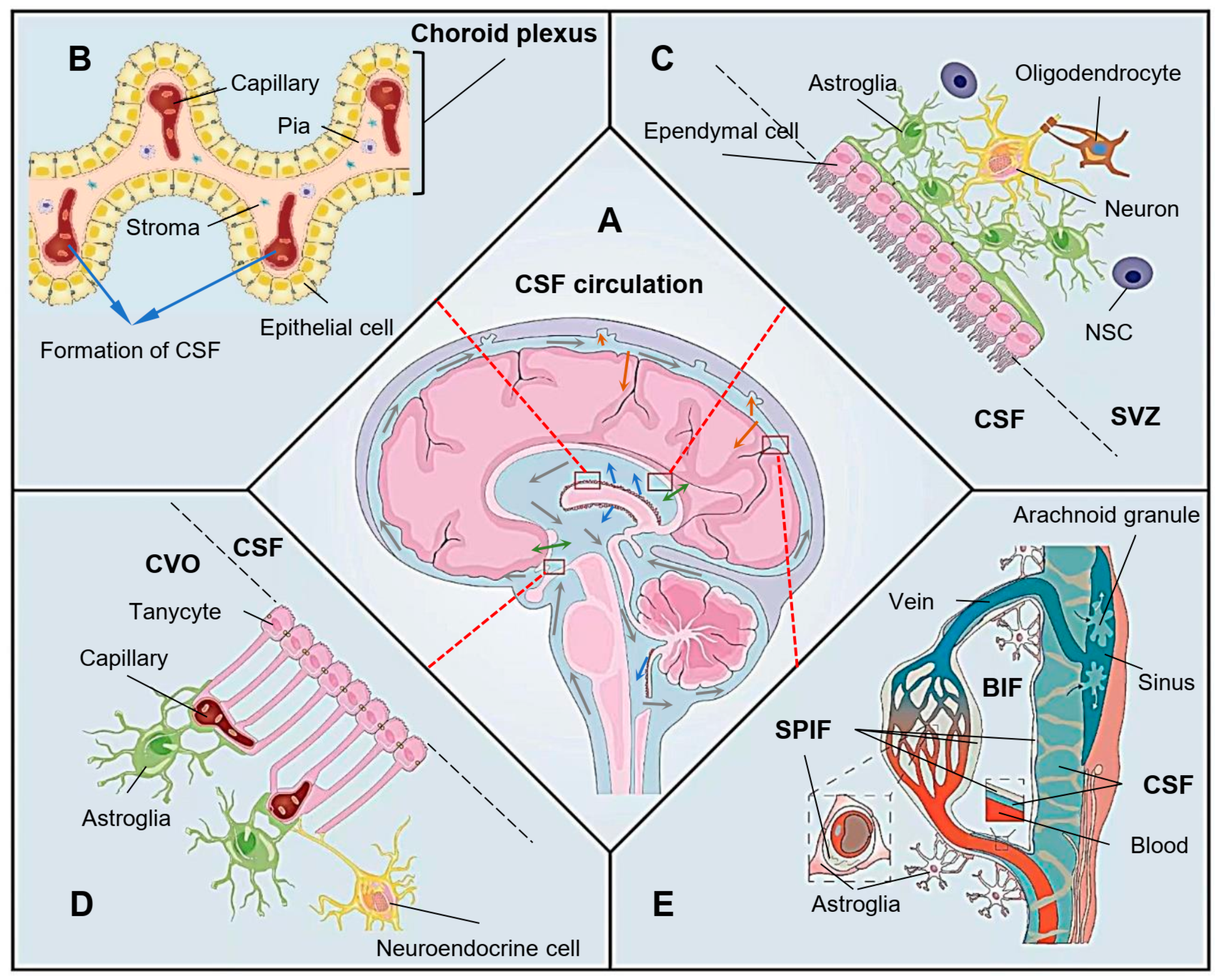
Figure 1. Role of the ependyma in cerebrospinal fluid (CSF) flow dynamics. (A) Generation, substance exchange, circulation and draining of CSF. The blue arrows indicate the generation of CSF from the choroid plexus; the grey arrows indicate the direction of CSF flow through the brain ventricular system and subarachnoid space; the green arrows indicate the substance exchange of CSF and brain interstitial fluid (BIF) or blood at the ventricular wall; the orange arrows indicate the draining of CSF at the perivascular spaces and arachnoid granules. (B) The formation of CSF from the choroid plexus, which is formed by a single layer of epithelial cells; the stroma derived from pia mater; and the capillary endothelium. (C) Ependyma and astroglia form the ventricular wall, which functions as the brain–CSF barrier. The neural stem cell (NSC) located in the niche of subventricular zone (SVZ) may proliferate to repair the damaged ependyma and regenerate astroglia to restore the barrier. (D) Tanycytes interact with astroglia and blood vessels to form a three-directional interface facilitating substance exchange among CSF, BIF and blood. These “brain windows” of the circumventricular organs (CVOs) play a vital role in the transportation of hypothalamic regulatory peptides and other factors. (E) CSF flows into the perivascular space and drains into the subpial interstitial fluid (SPIF), which can exchange with the BIF through the astroglial barrier and the blood through the endothelium. The CSF here also acts as a glymphatic system to introduce immune supervision and facilitate waste clearance from the BIF. Arachnoid granules may function as the location for “dirty” CSF draining back into the vein sinus or lymphatic pathway.
Tanycytes are highly specialized ECs that play a vital role in forming the ependyma of the circumventricular organs (CVOs) [14]. Often described as “brain windows”, the CVOs are rich capillary networks closely contacted with tanycytes and continued with the neighboring choroid plexus. This unique structure allows a potential functional relationship of the capillary system with CSF. For example, the median eminence (ME) is a well-known CVO located in the tuberal region of the hypothalamus [15] (Figure 1D). Tanycytes are bi-ciliated or mono-ciliated ECs with less motility than multi-ciliated ECs, but have long processes that can across the hypothalamic parenchyma and link the ventricular and vascular compartments directly [8][16][17]. In the ME, the adjacent tanycytes adhere with each other by various tight junction proteins, including ZO-1, occludin, claudin 1 and claudin 5, to prevent the free passage of molecules through the paracellular pathway [18][19][20]. These tight junctions of tanycytes at the ventricular surface of the CVO can prevent the diffusion of blood-borne molecules into the CSF, even if those molecules have permeated into the parenchyma of the ME through the vasculature surface of the CVO. Tanycytes also take part in forming the BBB between the hypothalamic parenchyma and capillary to maintain the microenvironment surrounding those neuroendocrine cells and facilitate the release of hypothalamic regulatory peptides [16][17][20] (Figure 1D). In addition, the tanycytes may play a vital role in metabolic homeostasis by secreting or transporting circulatory fibroblast growth factor 21 (FGF21) into the central nervous system [21]. Unlike the stricter BBB formed intactly by astrocytes, tanycytes may provide a “window” for brain invasion while promoting substance exchange at the vasculature interface of the CVO [20][22].
The CSF produced in the brain ventricular system flows into the subarachnoid space through the median and lateral foramen of the fourth ventricle [23]. From here, the pia mater replaces the ependyma to form BCB. CSF circulating in the subarachnoid space drains into the subpial interstitial fluid (SPIF) from the perivascular spaces and exchanges substances with the BIF through the astroglial barrier or the blood through the capillary [24][25][26] (Figure 1E). Arachnoid granulations, which have been considered as the main pathway for absorbing CSF into venous sinus, may function as glymphatic–lymphatic coupling structures together with the newly unveiled subarachnoid lymphatic-like membrane (SLYM) [27][28][29]. The CSF-glymphatic communication through SLYM supervises the immune status of CSF and presents the information to the lymphatic and/or blood system through arachnoid granulations. Although the pia mater and ependyma develop from different origins, they both contribute much in maintaining the delicate balance of CSF dynamic flow and biochemical homeostasis in the brain microenvironment [30].
2. Ependymal Dysfunctions in the Pathogenesis of Neurodegenerative Diseases
Neurodegenerative diseases are associated with the abnormal transportation of metabolites or other substances among intracellular fluid, interstitial fluid, CSF and blood in the brain [31]. CSF is mainly produced in the choroid plexus and transported from the lateral ventricle to the third ventricle, aqueduct and fourth ventricle, and then is re-inhaled in the subarachnoid space [7]. As a fluid clearing pathway in the brain, the glymphatic–lymphatic pathway helps to drain the CSF from the subarachnoid space into the perivascular spaces of penetrating arteries, also known as Virchow–Robin spaces [32][33][34]. From these perivascular spaces, CSF can finally return to the brain parenchyma and/or the cerebral vasculature [32]. This lymphatic pathway dominates during sleep. It has been reported that the clearance rate of harmful metabolites (such as Aβ) during sleep was significantly higher than that during awaking [35][36]. During sleep, the BIF secreted from astroglia dilutes the extracellular metabolites and washes them away by the increased BIF advection in the larger interstitial space [37]. Recent research has demonstrated that the etiology of AD and other neurodegenerative diseases may involve the abnormal expression of lipoproteins from the reactive astrocytes, such as the intensively studied APOE4, and the neurotoxic lipids they transport [38][39][40]. These toxic lipids may disturb lipid metabolism in brain tissue and destroy the membrane structures, especially the ependyma. The tanycytes have been reported to have an important role in regulating lipid metabolism [41]. The aged mouse EC possesses more lipid droplet accumulation and loses its barrier function [42]. This metabolic alteration in EC can cause the aging of EC, the dysfunction of ependyma and cognitive impairment [43]. Neuron stem cell and other progenitors in the subependymal area, such as the SVZ, can repair the damaged ependyma; however, they sometimes induce gliosis on the surface of the ventricular wall [31][44][45]. The cross-talk between astroglial and microglia activation, perivascular macrophage migration and immune cell infiltration in SVZ after brain injury may affect periventricular interstitial fluid homeostasis and impair ependymal function [46][47].
The filtration of water through ependyma is mainly controlled by aquaporin 4 (AQP4), the most abundant aquaporin in the mammalian brain [10]. Increased AQP4 expression was detected at the gliosis site of ependyma that impaired the CSF/BIF dynamic balance and the clearance of interstitial solutes [31][48]. On the other hand, deletion of AQP4 can obviously prevent the cytotoxic edema after stroke [48][49]. The abnormal expression of AQP4 is also involved in the dysfunction of the lymphatic pathway in animal models of traumatic brain injury, AD and stroke [50]. A higher AQP4 level was found in the ECs after subarachnoid hemorrhage, and the expression level of AQP4 was related with the severity of hydrocephalus [51]. The autoimmune antibodies from the patients of neuromyelitis optica can target AQP4 on the surface of ECs to trigger the functional impairment and inflammatory response in ependyma [52]. There is no doubt that AQP4 variation is associated with genetic susceptibility to PD [53]. The choroid plexus epithelium also expresses other AQPs including AQP1, AQP5 and AQP7, which more or less contribute to CSF production [10].
The normal activity of the ependymal motile cilia ensures the necessary CSF circulation to maintain brain homoeostasis, wash out toxins, deliver signal molecules and orient the migration of new-born neurons [54]. However, the molecular mechanism underlying the maintenance of ependymal motile cilia remains unclear [55]. The highly conserved cilia project from the apical surface and the zonula adherens on the lateral surface of ECs to move the overlying fluid by coordinated beating [12]. These cilia arise from the basal bodies, which are docked on the cell surfaces and rotationally polarized toward the CSF [56][57]. It has been suggested that this complementary polarization of the ependymal cilia should be regulated by the planar cell polarity pathway, which coordinates cell behavior in a plane of tissue cells [58][59]. The motile cilia dyskinesia can cause chronic recurrent respiratory infections, infertility, hydrocephalus and laterality defect [60][61][62]. Defective ependymal cilia motility is associated with the hydrocephalus, increased intracranial pressure and many neurological diseases [63][64]. Ciliary defects in mouse ECs can disrupt the CSF flow and lead to hydrocephalus and disoriented neuroblast migration in the SVZ [65][66]. Connexin 43 (Cx43), the dominating connexin of gap junction in the brain, plays a vital role in maintaining ependymal cilia [1]. Deletion of Cx43 can reduce the ciliary activity of ECs in zebrafish and mouse [1]. Possibly, the absence of Cx43 may affect the polarization of the ependymal cilia through the planar cell polarity pathway.
The neurodegenerative diseases share similar changes in the brain at the early stage, such as hydrocephalus [67] and ventricular broadening [64]. To date, the final diagnosis of AD can only be made by histopathological detection of Aβ plaques and neurofibrillary tangles post mortem [68]. For the purpose of early diagnosis and prevention of AD, positron-emission tomography (PET) has been used to analyze the synaptic dysfunction and cerebral Aβ load in the brain of AD model mice [69]. The data indicated that the glucose metabolism was decreased and the Aβ deposition was increased in AD mouse brain. The decreased glucose metabolism in AD may be due to the dysfunction of those glucose transporters expressed in the BBB, choroid plexus and ependyma [70][71]. Interestingly, high glucose or fructose concentration can directly stimulate the expression of brain-derived neurotrophic factor (BDNF) in the mouse microglia SIM-A9 cell [72]. Besides Aβ accumulation, more activated microglia have also been reported in AD animal models and in patients [73][74]. The Aβ plaque can activate microglia to form the plaque-microglial complex, and then significantly alter the gene expression and biological function of the surrounding astrocyte and oligodendrocyte precursor cell [75]. Furthermore, vascular risk factors such as hypercholesterolemia and hyperglycemia may also be involved in the genesis of AD and other neurodegenerative diseases [35]. The severity of cerebral atherosclerosis and/or arteriolosclerosis are associated with cognitive dysfunction [76]. Improving Aβ clearance along the perivascular pathway may provide a feasible therapeutic approach to control the progression of AD [77]. Recent research demonstrated that the CSF macrophages near the border of brain parenchyma had a role in regulating CSF flow dynamics by delicate clearance of the extra accumulated extracellular matrix proteins [78]. The single-nucleus RNA sequencing data obtained from the AD patients and the animal model of AD demonstrated abnormal transcriptomic alterations in these macrophages [78]. Intracisternal injection of macrophage colony-stimulating factor can improve the function of CSF macrophages and restore the CSF flow, implicating a new strategy to counteract the deficient CSF dynamics [78].
Similarly, the early diagnosis of PD, especially the premotor phase, is difficult in a clinic setting. Intracellular accumulation of the α-syn aggregates is the major pathological change of PD [79]. A previous study demonstrated that the changes in sleep-related oscillations should be an early consequence of abnormal α-syn aggregation in the mouse model [80]. The lymphatic pathway helps to drain the harmful substances in the cerebral interstitial fluid and CSF through the perivascular spaces of penetrating arteries, especially during sleep [30]. Chronic sleep deprivation or circadian disruption may cause lymphatic pathway dysfunction in the brain. The consequent abnormal accumulation of α-syn or other harmful substances caused by this BCB dysfunction will consequently result in AD, PD, depression and anxiety [80][81][82][83].
HD is a genetic neurodegenerative disease caused by the abnormal expansion of the CAG trinucleotide repeat in the huntingtin gene, which leads to a polyglutamine strand at the N-terminus of huntingtin protein [84]. Current therapeutic strategies designed for HD focus on reducing cytoplasmic aggregation of the mutant huntingtin protein [85]. Most cases of ALS are also characterized by the abnormal cytoplasmic aggregation of different proteins including TAR DNA binding protein 43 (TDP-43), Cu–Zn superoxide dismutase (SOD1), ubiquitin/p62 and others [86][87][88]. Unlike the HD, many genetic mutants have been identified in the ALS patients. Therefore, it is complicated to explain the pathogenesis of ALS. Ageing or exogenous risk factors may accelerate these inherited sensitivities and cause the onset of neurodegeneration [86]. Without considering the initiation of neuron damage, the activation of microglia and astroglia may contribute to the progressive motor neuron loss in ALS [86][87]. In human HD brains, the inflammatory activation of astroglia in the caudate nucleus and the subependymal layer was indicated by the co-localization of RAGE with its ligands and the nucleus translocation of NF-κB [89].
Under most circumstance, preventing neuroinflammation at an early stage can improve the cognitive impairment [90][91]. However, inhibition of the proinflammatory kinase IKKβ accelerates HD progression in mice because IKKβ has a role in phosphorylating huntingtin [92]. A review article suggested that the activation of microglia and astroglia in brain tissue may promote the BBB restoration, limit the blood-derived immune cell infiltration, trap the infiltrated T cells and achieve the early resolution of neuroinflammation in multiple sclerosis [93]. The age-related cerebral microvascular dysfunction and microbleeding destroy the integrity of BBB and allow the entry of peripheral neurotoxic substances, macrophages and neutrophils [94][95][96][97][98][99]. These factors can activate microglia and astroglia in the brain to release pro-inflammatory cytokines that induce chronic neuroinflammation and further brain injury. The anti-inflammatory reagent OKN-007 has shown effects on reversing lipopolysaccharide (LPS)-induced long-term neuroinflammatory responses and BBB impairment [100].
References
- Zhang, J.; Chandrasekaran, G.; Li, W.; Kim, D.Y.; Jeong, I.Y.; Lee, S.H.; Liang, T.; Bae, J.Y.; Choi, I.; Kang, H.; et al. Wnt-PLC-IP(3)-Connexin-Ca(2+) axis maintains ependymal motile cilia in zebrafish spinal cord. Nat. Commun. 2020, 11, 1860.
- Lim, D.A.; Alvarez-Buylla, A. The Adult Ventricular-Subventricular Zone (V-SVZ) and Olfactory Bulb (OB) Neurogenesis. Cold Spring Harb. Perspect. Biol. 2016, 8, a018820.
- Luo, Y.; Coskun, V.; Liang, A.; Yu, J.; Cheng, L.; Ge, W.; Shi, Z.; Zhang, K.; Li, C.; Cui, Y.; et al. Single-cell transcriptome analyses reveal signals to activate dormant neural stem cells. Cell 2015, 161, 1175–1186.
- Shah, P.T.; Stratton, J.A.; Stykel, M.G.; Abbasi, S.; Sharma, S.; Mayr, K.A.; Koblinger, K.; Whelan, P.J.; Biernaskie, J. Single-Cell Transcriptomics and Fate Mapping of Ependymal Cells Reveals an Absence of Neural Stem Cell Function. Cell 2018, 173, 1045–1057.e9.
- Ren, Y.; Ao, Y.; O’Shea, T.M.; Burda, J.E.; Bernstein, A.M.; Brumm, A.J.; Muthusamy, N.; Ghashghaei, H.T.; Carmichael, S.T.; Cheng, L.; et al. Ependymal cell contribution to scar formation after spinal cord injury is minimal, local and dependent on direct ependymal injury. Sci. Rep. 2017, 7, 41122.
- Spassky, N.; Merkle, F.T.; Flames, N.; Tramontin, A.D.; Garcia-Verdugo, J.M.; Alvarez-Buylla, A. Adult ependymal cells are postmitotic and are derived from radial glial cells during embryogenesis. J. Neurosci. 2005, 25, 10–18.
- Louvi, A.; Grove, E.A. Cilia in the CNS: The quiet organelle claims center stage. Neuron 2011, 69, 1046–1060.
- Mirzadeh, Z.; Kusne, Y.; Duran-Moreno, M.; Cabrales, E.; Gil-Perotin, S.; Ortiz, C.; Chen, B.; Garcia-Verdugo, J.M.; Sanai, N.; Alvarez-Buylla, A. Bi- and uniciliated ependymal cells define continuous floor-plate-derived tanycytic territories. Nat. Commun. 2017, 8, 13759.
- Roales-Bujan, R.; Paez, P.; Guerra, M.; Rodriguez, S.; Vio, K.; Ho-Plagaro, A.; Garcia-Bonilla, M.; Rodriguez-Perez, L.M.; Dominguez-Pinos, M.D.; Rodriguez, E.M.; et al. Astrocytes acquire morphological and functional characteristics of ependymal cells following disruption of ependyma in hydrocephalus. Acta Neuropathol. 2012, 124, 531–546.
- Municio, C.; Carrero, L.; Antequera, D.; Carro, E. Choroid Plexus Aquaporins in CSF Homeostasis and the Glymphatic System: Their Relevance for Alzheimer’s Disease. Int. J. Mol. Sci. 2023, 24, 878.
- Bethlehem, R.A.I.; Seidlitz, J.; White, S.R.; Vogel, J.W.; Anderson, K.M.; Adamson, C.; Adler, S.; Alexopoulos, G.S.; Anagnostou, E.; Areces-Gonzalez, A.; et al. Brain charts for the human lifespan. Nature 2022, 604, 525–533.
- Del Bigio, M.R. Ependymal cells: Biology and pathology. Acta Neuropathol. 2010, 119, 55–73.
- Johanson, C.; Stopa, E.; McMillan, P.; Roth, D.; Funk, J.; Krinke, G. The distributional nexus of choroid plexus to cerebrospinal fluid, ependyma and brain: Toxicologic/pathologic phenomena, periventricular destabilization, and lesion spread. Toxicol. Pathol. 2011, 39, 186–212.
- Gross, P.M.; Weindl, A. Peering through the windows of the brain. J. Cereb. Blood Flow Metab. 1987, 7, 663–672.
- Langlet, F.; Mullier, A.; Bouret, S.G.; Prevot, V.; Dehouck, B. Tanycyte-like cells form a blood-cerebrospinal fluid barrier in the circumventricular organs of the mouse brain. J. Comp. Neurol. 2013, 521, 3389–3405.
- Bolborea, M.; Langlet, F. What is the physiological role of hypothalamic tanycytes in metabolism? Am. J. Physiol. Regul. Integr. Comp. Physiol. 2021, 320, R994–R1003.
- Takemura, S.; Isonishi, A.; Horii-Hayashi, N.; Tanaka, T.; Tatsumi, K.; Komori, T.; Yamamuro, K.; Yamano, M.; Nishi, M.; Makinodan, M.; et al. Juvenile social isolation affects the structure of the tanycyte-vascular interface in the hypophyseal portal system of the adult mice. Neurochem. Int. 2023, 162, 105439.
- Mullier, A.; Bouret, S.G.; Prevot, V.; Dehouck, B. Differential distribution of tight junction proteins suggests a role for tanycytes in blood-hypothalamus barrier regulation in the adult mouse brain. J. Comp. Neurol. 2010, 518, 943–962.
- Langlet, F.; Levin, B.E.; Luquet, S.; Mazzone, M.; Messina, A.; Dunn-Meynell, A.A.; Balland, E.; Lacombe, A.; Mazur, D.; Carmeliet, P.; et al. Tanycytic VEGF-A boosts blood-hypothalamus barrier plasticity and access of metabolic signals to the arcuate nucleus in response to fasting. Cell Metab. 2013, 17, 607–617.
- Miyata, S. Glial functions in the blood-brain communication at the circumventricular organs. Front. Neurosci. 2022, 16, 991779.
- Geller, S.; Arribat, Y.; Netzahualcoyotzi, C.; Lagarrigue, S.; Carneiro, L.; Zhang, L.; Amati, F.; Lopez-Mejia, I.C.; Pellerin, L. Tanycytes Regulate Lipid Homeostasis by Sensing Free Fatty Acids and Signaling to Key Hypothalamic Neuronal Populations via FGF21 Secretion. Cell Metab. 2019, 30, 833–844.e7.
- Bentivoglio, M.; Kristensson, K.; Rottenberg, M.E. Circumventricular Organs and Parasite Neurotropism: Neglected Gates to the Brain? Front. Immunol. 2018, 9, 2877.
- Adigun, O.O.; Al-Dhahir, M.A. Anatomy, Head and Neck, Cerebrospinal Fluid. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022.
- Weller, R.O.; Sharp, M.M.; Christodoulides, M.; Carare, R.O.; Mollgard, K. The meninges as barriers and facilitators for the movement of fluid, cells and pathogens related to the rodent and human CNS. Acta Neuropathol. 2018, 135, 363–385.
- Rasmussen, M.K.; Mestre, H.; Nedergaard, M. Fluid transport in the brain. Physiol. Rev. 2022, 102, 1025–1151.
- Mestre, H.; Verma, N.; Greene, T.D.; Lin, L.A.; Ladron-de-Guevara, A.; Sweeney, A.M.; Liu, G.; Thomas, V.K.; Galloway, C.A.; de Mesy Bentley, K.L.; et al. Periarteriolar spaces modulate cerebrospinal fluid transport into brain and demonstrate altered morphology in aging and Alzheimer’s disease. Nat. Commun. 2022, 13, 3897.
- Shah, T.; Leurgans, S.E.; Mehta, R.I.; Yang, J.; Galloway, C.A.; de Mesy Bentley, K.L.; Schneider, J.A.; Mehta, R.I. Arachnoid granulations are lymphatic conduits that communicate with bone marrow and dura-arachnoid stroma. J. Exp. Med. 2023, 220, e20220618.
- Mollgard, K.; Beinlich, F.R.M.; Kusk, P.; Miyakoshi, L.M.; Delle, C.; Pla, V.; Hauglund, N.L.; Esmail, T.; Rasmussen, M.K.; Gomolka, R.S.; et al. A mesothelium divides the subarachnoid space into functional compartments. Science 2023, 379, 84–88.
- Bissenas, A.; Fleeting, C.; Patel, D.; Al-Bahou, R.; Patel, A.; Nguyen, A.; Woolridge, M.; Angelle, C.; Lucke-Wold, B. CSF Dynamics: Implications for Hydrocephalus and Glymphatic Clearance. Curr. Res. Med. Sci. 2022, 1, 24–42.
- Xie, L.; Kang, H.; Xu, Q.; Chen, M.J.; Liao, Y.; Thiyagarajan, M.; O’Donnell, J.; Christensen, D.J.; Nicholson, C.; Iliff, J.J.; et al. Sleep drives metabolite clearance from the adult brain. Science 2013, 342, 373–377.
- Shook, B.A.; Lennington, J.B.; Acabchuk, R.L.; Halling, M.; Sun, Y.; Peters, J.; Wu, Q.; Mahajan, A.; Fellows, D.W.; Conover, J.C. Ventriculomegaly associated with ependymal gliosis and declines in barrier integrity in the aging human and mouse brain. Aging Cell 2014, 13, 340–350.
- Aspelund, A.; Antila, S.; Proulx, S.T.; Karlsen, T.V.; Karaman, S.; Detmar, M.; Wiig, H.; Alitalo, K. A dural lymphatic vascular system that drains brain interstitial fluid and macromolecules. J. Exp. Med. 2015, 212, 991–999.
- Ahn, J.H.; Cho, H.; Kim, J.H.; Kim, S.H.; Ham, J.S.; Park, I.; Suh, S.H.; Hong, S.P.; Song, J.H.; Hong, Y.K.; et al. Meningeal lymphatic vessels at the skull base drain cerebrospinal fluid. Nature 2019, 572, 62–66.
- Hannocks, M.J.; Pizzo, M.E.; Huppert, J.; Deshpande, T.; Abbott, N.J.; Thorne, R.G.; Sorokin, L. Molecular characterization of perivascular drainage pathways in the murine brain. J. Cereb. Blood Flow Metab. 2018, 38, 669–686.
- Toledo, J.B.; Arnold, S.E.; Raible, K.; Brettschneider, J.; Xie, S.X.; Grossman, M.; Monsell, S.E.; Kukull, W.A.; Trojanowski, J.Q. Contribution of cerebrovascular disease in autopsy confirmed neurodegenerative disease cases in the National Alzheimer’s Coordinating Centre. Brain 2013, 136, 2697–2706.
- Iliff, J.J.; Wang, M.; Liao, Y.; Plogg, B.A.; Peng, W.; Gundersen, G.A.; Benveniste, H.; Vates, G.E.; Deane, R.; Goldman, S.A.; et al. A paravascular pathway facilitates CSF flow through the brain parenchyma and the clearance of interstitial solutes, including amyloid beta. Sci. Transl. Med. 2012, 4, 147ra111.
- Thomas, J.H. Theoretical analysis of wake/sleep changes in brain solute transport suggests a flow of interstitial fluid. Fluids Barriers CNS 2022, 19, 30.
- Jackson, R.J.; Meltzer, J.C.; Nguyen, H.; Commins, C.; Bennett, R.E.; Hudry, E.; Hyman, B.T. APOE4 derived from astrocytes leads to blood-brain barrier impairment. Brain 2022, 145, 3582–3593.
- Blanchard, J.W.; Akay, L.A.; Davila-Velderrain, J.; von Maydell, D.; Mathys, H.; Davidson, S.M.; Effenberger, A.; Chen, C.Y.; Maner-Smith, K.; Hajjar, I.; et al. APOE4 impairs myelination via cholesterol dysregulation in oligodendrocytes. Nature 2022, 611, 769–779.
- Guttenplan, K.A.; Weigel, M.K.; Prakash, P.; Wijewardhane, P.R.; Hasel, P.; Rufen-Blanchette, U.; Munch, A.E.; Blum, J.A.; Fine, J.; Neal, M.C.; et al. Neurotoxic reactive astrocytes induce cell death via saturated lipids. Nature 2021, 599, 102–107.
- Kim, S.; Kim, N.; Park, S.; Jeon, Y.; Lee, J.; Yoo, S.J.; Lee, J.W.; Moon, C.; Yu, S.W.; Kim, E.K. Tanycytic TSPO inhibition induces lipophagy to regulate lipid metabolism and improve energy balance. Autophagy 2020, 16, 1200–1220.
- Muthusamy, N.; Sommerville, L.J.; Moeser, A.J.; Stumpo, D.J.; Sannes, P.; Adler, K.; Blackshear, P.J.; Weimer, J.M.; Ghashghaei, H.T. MARCKS-dependent mucin clearance and lipid metabolism in ependymal cells are required for maintenance of forebrain homeostasis during aging. Aging Cell 2015, 14, 764–773.
- Muthusamy, N.; Williams, T.I.; O’Toole, R.; Brudvig, J.J.; Adler, K.B.; Weimer, J.M.; Muddiman, D.C.; Ghashghaei, H.T. Phosphorylation-dependent proteome of Marcks in ependyma during aging and behavioral homeostasis in the mouse forebrain. Geroscience 2022, 44, 2077–2094.
- Del Carmen Gomez-Roldan, M.; Perez-Martin, M.; Capilla-Gonzalez, V.; Cifuentes, M.; Perez, J.; Garcia-Verdugo, J.M.; Fernandez-Llebrez, P. Neuroblast proliferation on the surface of the adult rat striatal wall after focal ependymal loss by intracerebroventricular injection of neuraminidase. J. Comp. Neurol. 2008, 507, 1571–1587.
- Luo, J.; Shook, B.A.; Daniels, S.B.; Conover, J.C. Subventricular zone-mediated ependyma repair in the adult mammalian brain. J. Neurosci. 2008, 28, 3804–3813.
- Garcia-Bonilla, M.; Castaneyra-Ruiz, L.; Zwick, S.; Talcott, M.; Otun, A.; Isaacs, A.M.; Morales, D.M.; Limbrick, D.D., Jr.; McAllister, J.P., 2nd. Acquired hydrocephalus is associated with neuroinflammation, progenitor loss, and cellular changes in the subventricular zone and periventricular white matter. Fluids Barriers CNS 2022, 19, 17.
- Piehl, N.; van Olst, L.; Ramakrishnan, A.; Teregulova, V.; Simonton, B.; Zhang, Z.; Tapp, E.; Channappa, D.; Oh, H.; Losada, P.M.; et al. Cerebrospinal fluid immune dysregulation during healthy brain aging and cognitive impairment. Cell 2022, 185, 5028–5039.e13.
- Sofroniew, M.V. Molecular dissection of reactive astrogliosis and glial scar formation. Trends Neurosci. 2009, 32, 638–647.
- Haj-Yasein, N.N.; Vindedal, G.F.; Eilert-Olsen, M.; Gundersen, G.A.; Skare, O.; Laake, P.; Klungland, A.; Thoren, A.E.; Burkhardt, J.M.; Ottersen, O.P.; et al. Glial-conditional deletion of aquaporin-4 (Aqp4) reduces blood-brain water uptake and confers barrier function on perivascular astrocyte endfeet. Proc. Natl. Acad. Sci. USA 2011, 108, 17815–17820.
- Abbott, N.J.; Pizzo, M.E.; Preston, J.E.; Janigro, D.; Thorne, R.G. The role of brain barriers in fluid movement in the CNS: Is there a ‘glymphatic’ system? Acta Neuropathol. 2018, 135, 387–407.
- Long, C.Y.; Huang, G.Q.; Du, Q.; Zhou, L.Q.; Zhou, J.H. The dynamic expression of aquaporins 1 and 4 in rats with hydrocephalus induced by subarachnoid haemorrhage. Folia Neuropathol. 2019, 57, 182–195.
- Bigotte, M.; Gimenez, M.; Gavoille, A.; Deligiannopoulou, A.; El Hajj, A.; Croze, S.; Goumaidi, A.; Malleret, G.; Salin, P.; Giraudon, P.; et al. Ependyma: A new target for autoantibodies in neuromyelitis optica? Brain Commun. 2022, 4, fcac307.
- Sun, X.; Tian, Q.; Yang, Z.; Liu, Y.; Li, C.; Hou, B.; Xie, A. Association of AQP4 single nucleotide polymorphisms (rs335929 and rs2075575) with Parkinson’s disease: A case-control study. Neurosci. Lett. 2023, 797, 137062.
- Zappaterra, M.W.; Lehtinen, M.K. The cerebrospinal fluid: Regulator of neurogenesis, behavior, and beyond. Cell Mol. Life Sci. 2012, 69, 2863–2878.
- Jimenez, A.J.; Dominguez-Pinos, M.D.; Guerra, M.M.; Fernandez-Llebrez, P.; Perez-Figares, J.M. Structure and function of the ependymal barrier and diseases associated with ependyma disruption. Tissue Barriers 2014, 2, e28426.
- Mirzadeh, Z.; Han, Y.G.; Soriano-Navarro, M.; Garcia-Verdugo, J.M.; Alvarez-Buylla, A. Cilia organize ependymal planar polarity. J. Neurosci. 2010, 30, 2600–2610.
- Wallingford, J.B. Planar cell polarity signaling, cilia and polarized ciliary beating. Curr. Opin. Cell Biol. 2010, 22, 597–604.
- Kishimoto, N.; Sawamoto, K. Planar polarity of ependymal cilia. Differentiation 2012, 83, S86–S90.
- Butler, M.T.; Wallingford, J.B. Planar cell polarity in development and disease. Nat. Rev. Mol. Cell Biol. 2017, 18, 375–388.
- Lucas, J.S.; Davis, S.D.; Omran, H.; Shoemark, A. Primary ciliary dyskinesia in the genomics age. Lancet Respir. Med. 2020, 8, 202–216.
- Mitchison, H.M.; Valente, E.M. Motile and non-motile cilia in human pathology: From function to phenotypes. J. Pathol. 2017, 241, 294–309.
- Reiter, J.F.; Leroux, M.R. Genes and molecular pathways underpinning ciliopathies. Nat. Rev. Mol. Cell Biol. 2017, 18, 533–547.
- Zhang, J.; Williams, M.A.; Rigamonti, D. Genetics of human hydrocephalus. J. Neurol. 2006, 253, 1255–1266.
- Palha, J.A.; Santos, N.C.; Marques, F.; Sousa, J.; Bessa, J.; Miguelote, R.; Sousa, N.; Belmonte-de-Abreu, P. Do genes and environment meet to regulate cerebrospinal fluid dynamics? Relevance for schizophrenia. Front. Cell. Neurosci. 2012, 6, 31.
- Ohata, S.; Alvarez-Buylla, A. Planar Organization of Multiciliated Ependymal (E1) Cells in the Brain Ventricular Epithelium. Trends Neurosci. 2016, 39, 543–551.
- Ohata, S.; Nakatani, J.; Herranz-Perez, V.; Cheng, J.; Belinson, H.; Inubushi, T.; Snider, W.D.; Garcia-Verdugo, J.M.; Wynshaw-Boris, A.; Alvarez-Buylla, A. Loss of Dishevelleds disrupts planar polarity in ependymal motile cilia and results in hydrocephalus. Neuron 2014, 83, 558–571.
- Muller-Schmitz, K.; Krasavina-Loka, N.; Yardimci, T.; Lipka, T.; Kolman, A.G.J.; Robbers, S.; Menge, T.; Kujovic, M.; Seitz, R.J. Normal Pressure Hydrocephalus Associated with Alzheimer’s Disease. Ann. Neurol. 2020, 88, 703–711.
- DeTure, M.A.; Dickson, D.W. The neuropathological diagnosis of Alzheimer’s disease. Mol. Neurodegener. 2019, 14, 32.
- Rice, L.; Bisdas, S. The diagnostic value of FDG and amyloid PET in Alzheimer’s disease-A systematic review. Eur. J. Radiol. 2017, 94, 16–24.
- Koepsell, H. Glucose transporters in brain in health and disease. Pflug. Arch. 2020, 472, 1299–1343.
- An, Y.; Varma, V.R.; Varma, S.; Casanova, R.; Dammer, E.; Pletnikova, O.; Chia, C.W.; Egan, J.M.; Ferrucci, L.; Troncoso, J.; et al. Evidence for brain glucose dysregulation in Alzheimer’s disease. Alzheimer’s Dement. 2018, 14, 318–329.
- Aldhshan, M.S.; Jhanji, G.; Mizuno, T.M. Glucose and fructose directly stimulate brain-derived neurotrophic factor gene expression in microglia. Neuroreport 2022, 33, 583–589.
- Franke, T.N.; Irwin, C.; Bayer, T.A.; Brenner, W.; Beindorff, N.; Bouter, C.; Bouter, Y. In vivo Imaging with (18)F-FDG- and (18)F-Florbetaben-PET/MRI Detects Pathological Changes in the Brain of the Commonly Used 5XFAD Mouse Model of Alzheimer’s Disease. Front. Med. 2020, 7, 529.
- Ohm, D.T.; Fought, A.J.; Martersteck, A.; Coventry, C.; Sridhar, J.; Gefen, T.; Weintraub, S.; Bigio, E.; Mesulam, M.M.; Rogalski, E.; et al. Accumulation of neurofibrillary tangles and activated microglia is associated with lower neuron densities in the aphasic variant of Alzheimer’s disease. Brain Pathol. 2021, 31, 189–204.
- Zeng, H.; Huang, J.; Zhou, H.; Meilandt, W.J.; Dejanovic, B.; Zhou, Y.; Bohlen, C.J.; Lee, S.H.; Ren, J.; Liu, A.; et al. Integrative in situ mapping of single-cell transcriptional states and tissue histopathology in a mouse model of Alzheimer’s disease. Nat. Neurosci. 2023, 26, 430–446.
- Arvanitakis, Z.; Capuano, A.W.; Leurgans, S.E.; Bennett, D.A.; Schneider, J.A. Relation of cerebral vessel disease to Alzheimer’s disease dementia and cognitive function in elderly people: A cross-sectional study. Lancet Neurol. 2016, 15, 934–943.
- Arbel-Ornath, M.; Hudry, E.; Eikermann-Haerter, K.; Hou, S.; Gregory, J.L.; Zhao, L.; Betensky, R.A.; Frosch, M.P.; Greenberg, S.M.; Bacskai, B.J. Interstitial fluid drainage is impaired in ischemic stroke and Alzheimer’s disease mouse models. Acta Neuropathol. 2013, 126, 353–364.
- Drieu, A.; Du, S.; Storck, S.E.; Rustenhoven, J.; Papadopoulos, Z.; Dykstra, T.; Zhong, F.; Kim, K.; Blackburn, S.; Mamuladze, T.; et al. Parenchymal border macrophages regulate the flow dynamics of the cerebrospinal fluid. Nature 2022, 611, 585–593.
- Mollenhauer, B.; Caspell-Garcia, C.J.; Coffey, C.S.; Taylor, P.; Shaw, L.M.; Trojanowski, J.Q.; Singleton, A.; Frasier, M.; Marek, K.; Galasko, D.; et al. Longitudinal CSF biomarkers in patients with early Parkinson disease and healthy controls. Neurology 2017, 89, 1959–1969.
- Zhang, R.; Liu, Y.; Chen, Y.; Li, Q.; Marshall, C.; Wu, T.; Hu, G.; Xiao, M. Aquaporin 4 deletion exacerbates brain impairments in a mouse model of chronic sleep disruption. CNS Neurosci. Ther. 2020, 26, 228–239.
- Tobaldini, E.; Costantino, G.; Solbiati, M.; Cogliati, C.; Kara, T.; Nobili, L.; Montano, N. Sleep, sleep deprivation, autonomic nervous system and cardiovascular diseases. Neurosci. Biobehav. Rev. 2017, 74, 321–329.
- Musiek, E.S.; Holtzman, D.M. Mechanisms linking circadian clocks, sleep, and neurodegeneration. Science 2016, 354, 1004–1008.
- Koinis-Mitchell, D.; Rosario-Matos, N.; Ramirez, R.R.; Garcia, P.; Canino, G.J.; Ortega, A.N. Sleep, Depressive/Anxiety Disorders, and Obesity in Puerto Rican Youth. J. Clin. Psychol. Med. Settings 2017, 24, 59–73.
- Walker, F.O. Huntington’s disease. Lancet 2007, 369, 218–228.
- Tabrizi, S.J.; Ghosh, R.; Leavitt, B.R. Huntingtin Lowering Strategies for Disease Modification in Huntington’s Disease. Neuron 2019, 102, 899.
- Mead, R.J.; Shan, N.; Reiser, H.J.; Marshall, F.; Shaw, P.J. Amyotrophic lateral sclerosis: A neurodegenerative disorder poised for successful therapeutic translation. Nat. Rev. Drug Discov. 2023, 22, 185–212.
- Boillee, S.; Vande Velde, C.; Cleveland, D.W. ALS: A disease of motor neurons and their nonneuronal neighbors. Neuron 2006, 52, 39–59.
- Ramos-Campoy, O.; Avila-Polo, R.; Grau-Rivera, O.; Antonell, A.; Clarimon, J.; Rojas-Garcia, R.; Charif, S.; Santiago-Valera, V.; Hernandez, I.; Aguilar, M.; et al. Systematic Screening of Ubiquitin/p62 Aggregates in Cerebellar Cortex Expands the Neuropathological Phenotype of the C9orf72 Expansion Mutation. J. Neuropathol. Exp. Neurol. 2018, 77, 703–709.
- Kim, J.; Waldvogel, H.J.; Faull, R.L.; Curtis, M.A.; Nicholson, L.F. The RAGE receptor and its ligands are highly expressed in astrocytes in a grade-dependant manner in the striatum and subependymal layer in Huntington’s disease. J. Neurochem. 2015, 134, 927–942.
- Sun, J.; Zhang, S.; Zhang, X.; Zhang, X.; Dong, H.; Qian, Y. IL-17A is implicated in lipopolysaccharide-induced neuroinflammation and cognitive impairment in aged rats via microglial activation. J. Neuroinflamm. 2015, 12, 165.
- Yamanaka, D.; Kawano, T.; Nishigaki, A.; Aoyama, B.; Tateiwa, H.; Shigematsu-Locatelli, M.; Locatelli, F.M.; Yokoyama, M. Preventive effects of dexmedetomidine on the development of cognitive dysfunction following systemic inflammation in aged rats. J. Anesth. 2017, 31, 25–35.
- Ochaba, J.; Fote, G.; Kachemov, M.; Thein, S.; Yeung, S.Y.; Lau, A.L.; Hernandez, S.; Lim, R.G.; Casale, M.; Neel, M.J.; et al. IKKbeta slows Huntington’s disease progression in R6/1 mice. Proc. Natl. Acad. Sci. USA 2019, 116, 10952–10961.
- Ruiz, F.; Vigne, S.; Pot, C. Resolution of inflammation during multiple sclerosis. Semin. Immunopathol. 2019, 41, 711–726.
- Toth, P.; Tarantini, S.; Csiszar, A.; Ungvari, Z. Functional vascular contributions to cognitive impairment and dementia: Mechanisms and consequences of cerebral autoregulatory dysfunction, endothelial impairment, and neurovascular uncoupling in aging. Am. J. Physiol. Heart Circ. Physiol. 2017, 312, H1–H20.
- Tarantini, S.; Tran, C.H.T.; Gordon, G.R.; Ungvari, Z.; Csiszar, A. Impaired neurovascular coupling in aging and Alzheimer’s disease: Contribution of astrocyte dysfunction and endothelial impairment to cognitive decline. Exp. Gerontol. 2017, 94, 52–58.
- Sumbria, R.K.; Grigoryan, M.M.; Vasilevko, V.; Paganini-Hill, A.; Kilday, K.; Kim, R.; Cribbs, D.H.; Fisher, M.J. Aging exacerbates development of cerebral microbleeds in a mouse model. J. Neuroinflamm. 2018, 15, 69.
- Zang, L.; Yang, B.; Zhang, M.; Cui, J.; Ma, X.; Wei, L. Trelagliptin Mitigates Macrophage Infiltration by Preventing the Breakdown of the Blood-Brain Barrier in the Brain of Middle Cerebral Artery Occlusion Mice. Chem. Res. Toxicol. 2021, 34, 1016–1023.
- Xiang, P.; Chew, W.S.; Seow, W.L.; Lam, B.W.S.; Ong, W.Y.; Herr, D.R. The S1P(2) receptor regulates blood-brain barrier integrity and leukocyte extravasation with implications for neurodegenerative disease. Neurochem. Int. 2021, 146, 105018.
- Vila Verde, D.; de Curtis, M.; Librizzi, L. Seizure-Induced Acute Glial Activation in the in vitro Isolated Guinea Pig Brain. Front. Neurol. 2021, 12, 607603.
- Towner, R.A.; Saunders, D.; Smith, N.; Gulej, R.; McKenzie, T.; Lawrence, B.; Morton, K.A. Anti-inflammatory agent, OKN-007, reverses long-term neuroinflammatory responses in a rat encephalopathy model as assessed by multi-parametric MRI: Implications for aging-associated neuroinflammation. Geroscience 2019, 41, 483–494.
More
Information
Subjects:
Physiology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.3K
Revisions:
2 times
(View History)
Update Date:
06 May 2023
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No