 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Sherin Bakhashab | -- | 2690 | 2023-03-14 15:10:28 | | | |
| 2 | Lindsay Dong | Meta information modification | 2690 | 2023-03-15 06:46:35 | | |
Video Upload Options
Prostate cancer (PC) is a common malignancy and is one of the leading causes of cancer-related death in men worldwide. Osteosarcoma (OS) is the most common bone cancer, representing 20–40% of all bone malignancy cases. Cancer metastasis is a process by which malignant tumor cells detach from the primary tumor site via a cascade of processes and migrate to secondary sites through the blood circulation or lymphatic system to colonize and form secondary tumors. PC has a specific affinity to the bone based on the “seed and soil” theory; once PC reach the bone, it becomes incurable. Several studies have identified long noncoding RNAs (lncRNAs) as potential targets for cancer therapy or as diagnostic and prognostic biomarkers. The dysregulation of various lncRNAs has been found in various cancer types, including PC, OS, and metastasis.
1. Introduction
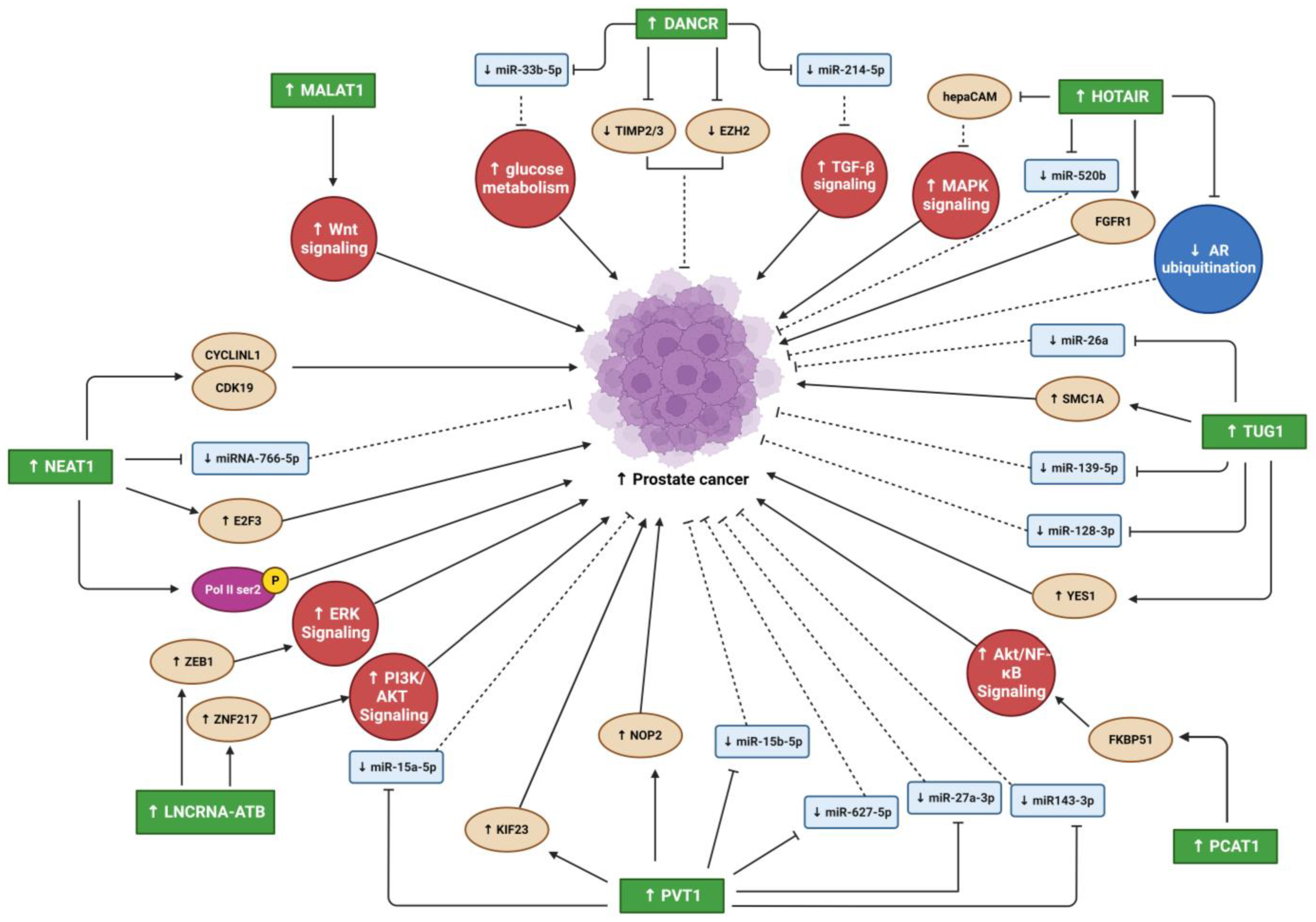
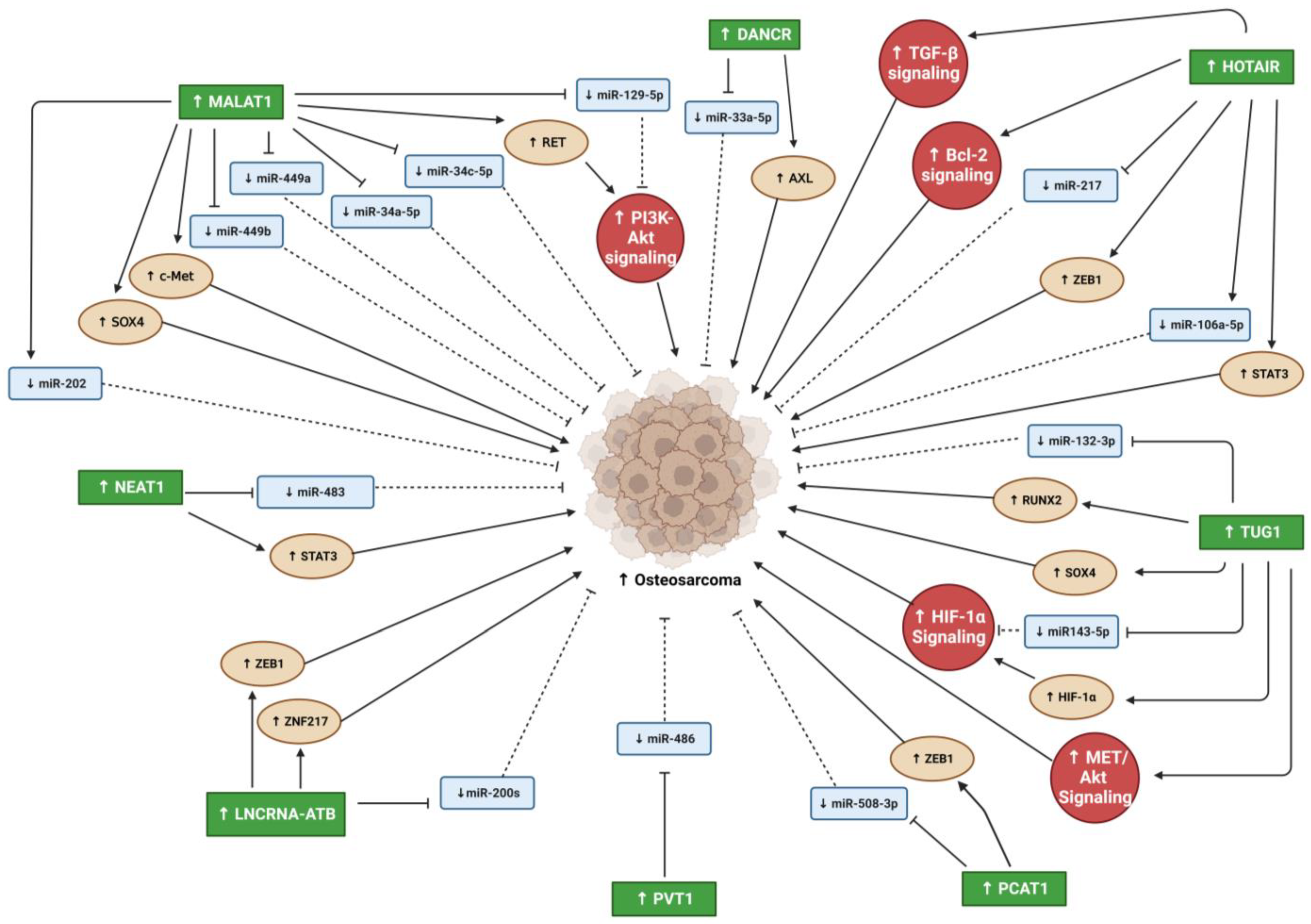
2. Oncogenic lncRNAs in PC and OS Malignancy
2.1. Differentiation Antagonizing Non-Protein Coding RNA
2.2. Metastasis Associated Lung Adenocarcinoma Transcript 1
2.3. Nuclear Enriched Abundant Transcript 1
2.4. HOX Transcript Antisense RNA
2.5. Taurine Upregulated Gene 1
2.6. PC Associated Transcript 1
2.7. LncRNA-Activated by TGF-β
2.8. Plasmacytoma Variant Translocation 1
3. Conclusions
References
- Wang, L.; Lu, B.; He, M.; Wang, Y.; Wang, Z.; Du, L. Prostate Cancer Incidence and Mortality: Global Status and Temporal Trends in 89 Countries From 2000 to 2019. Front. Public Health 2022, 10, 811044.
- Valery, P.C.; Laversanne, M.; Bray, F. Bone cancer incidence by morphological subtype: A global assessment. Cancer Causes Control 2015, 26, 1127–1139.
- Ghafouri-Fard, S.; Shirvani-Farsani, Z.; Hussen, B.M.; Taheri, M. The critical roles of lncRNAs in the development of osteosarcoma. Biomed. Pharmacother. 2021, 135, 111217.
- Rojas, G.A.; Hubbard, A.K.; Diessner, B.J.; Ribeiro, K.B.; Spector, L.G. International trends in incidence of osteosarcoma (1988–2012). Int. J. Cancer 2021, 149, 1044–1053.
- Singh, A.; Malvankar, S.; Ravi Kumar, Y.S.; Seervi, M.; Srivastava, R.K.; Verma, B. Role of various non-coding RNAs in EMT, cancer, and metastasis: Recent trends and future perspective. Adv. Cancer Biol.-Metastasis 2022, 4, 100039.
- Nie, H.; Liao, Z.; Wang, Y.; Zhou, J.; He, X.; Ou, C. Exosomal long non-coding RNAs: Emerging players in cancer metastasis and potential diagnostic biomarkers for personalized oncology. Genes Dis. 2021, 8, 769–780.
- Castaneda, M.; den Hollander, P.; Kuburich, N.A.; Rosen, J.M.; Mani, S.A. Mechanisms of cancer metastasis. Semin. Cancer Biol. 2022, 87, 17–31.
- Tay, K.J.; Moul, J.W.; Armstrong, A.J. Management of Prostate Cancer in the Elderly. Clin. Geriatr. Med. 2016, 32, 113–132.
- Li, Y.; Egranov, S.D.; Yang, L.; Lin, C. Molecular mechanisms of long noncoding RNAs-mediated cancer metastasis. Genes Chromosom. Cancer 2019, 58, 200–207.
- Kahn, D.; Weiner, G.J.; Ben-Haim, S.; Boles Ponto, L.L.; Madsen, M.T.; Bushnell, D.L.; Watkins, G.L.; Argenyi, E.A.; Hichwa, R.D. Positron Emission Tomographic Measurement of Bone Marrow Blood Flow to the Pelvis and Lumbar Vertebrae in Young Normal Adults. Blood 1994, 83, 958–963.
- Guise, T.A. The vicious cycle of bone metastases. J. Musculoskelet. Neuronal Interact. 2002, 2, 570–572.
- Guise, T.A.; Mohammad, K.S.; Clines, G.; Stebbins, E.G.; Wong, D.H.; Higgins, L.S.; Vessella, R.; Corey, E.; Padalecki, S.; Suva, L.; et al. Basic mechanisms responsible for osteolytic and osteoblastic bone metastases. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2006, 12, 6213s–6216s.
- Fizazi, K.; Yang, J.; Peleg, S.; Sikes, C.R.; Kreimann, E.L.; Daliani, D.; Olive, M.; Raymond, K.A.; Janus, T.J.; Logothetis, C.J.; et al. Prostate cancer cells-osteoblast interaction shifts expression of growth/survival-related genes in prostate cancer and reduces expression of osteoprotegerin in osteoblasts. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2003, 9, 2587–2597.
- Ottewell, P.D. The role of osteoblasts in bone metastasis. J. Bone Oncol. 2016, 5, 124–127.
- Casimiro, S.; Ferreira, A.R.; Mansinho, A.; Alho, I.; Costa, L. Molecular Mechanisms of Bone Metastasis: Which Targets Came from the Bench to the Bedside? Int. J. Mol. Sci. 2016, 17, 1415.
- David Roodman, G.; Silbermann, R. Mechanisms of osteolytic and osteoblastic skeletal lesions. BoneKEy Rep. 2015, 4, 753.
- Buijs, J.T.; Rentsch, C.A.; van der Horst, G.; van Overveld, P.G.; Wetterwald, A.; Schwaninger, R.; Henriquez, N.V.; Ten Dijke, P.; Borovecki, F.; Markwalder, R.; et al. BMP7, a putative regulator of epithelial homeostasis in the human prostate, is a potent inhibitor of prostate cancer bone metastasis in vivo. Am. J. Pathol. 2007, 171, 1047–1057.
- Chaffer, C.L.; Thompson, E.W.; Williams, E.D. Mesenchymal to epithelial transition in development and disease. Cells Tissues Organs 2007, 185, 7–19.
- van der Pluijm, G. Epithelial plasticity, cancer stem cells and bone metastasis formation. Bone 2011, 48, 37–43.
- Yu, X.; Yustein, J.T.; Xu, J. Research models and mesenchymal/epithelial plasticity of osteosarcoma. Cell Biosci. 2021, 11, 94.
- Sannino, G.; Marchetto, A.; Kirchner, T.; Grünewald, T.G.P. Epithelial-to-Mesenchymal and Mesenchymal-to-Epithelial Transition in Mesenchymal Tumors: A Paradox in Sarcomas? Cancer Res. 2017, 77, 4556–4561.
- Aird, J.; Baird, A.-M.; Lim, M.C.J.; McDermott, R.; Finn, S.P.; Gray, S.G. Carcinogenesis in prostate cancer: The role of long non-coding RNAs. Non-Coding RNA Res. 2018, 3, 29–38.
- Mukhametov, U.; Lyulin, S.; Borzunov, D.; Sufianova, G.; Shumadalova, A.; Zhang, D.; Gareev, I. Functions of the bone morphogenetic protein signaling pathway through non-coding RNAs. Non-Coding RNA Res. 2022, 7, 178–183.
- Sikora, M.; Marycz, K.; Smieszek, A. Small and Long Non-coding RNAs as Functional Regulators of Bone Homeostasis, Acting Alone or Cooperatively. Mol. Ther.-Nucleic Acids 2020, 21, 792–803.
- Volders, P.J.; Anckaert, J.; Verheggen, K.; Nuytens, J.; Martens, L.; Mestdagh, P.; Vandesompele, J. LNCipedia 5: Towards a reference set of human long non-coding RNAs. Nucleic Acids Res. 2019, 47, D135–D139.
- Liu, L.; Li, Z.; Liu, C.; Zou, D.; Li, Q.; Feng, C.; Jing, W.; Luo, S.; Zhang, Z.; Ma, L. LncRNAWiki 2.0: A knowledgebase of human long non-coding RNAs with enhanced curation model and database system. Nucleic Acids Res. 2022, 50, D190–D195.
- Wang, M.; Gu, J.; Zhang, X.; Yang, J.; Zhang, X.; Fang, X. Long Non-coding RNA DANCR in Cancer: Roles, Mechanisms, and Implications. Front. Cell Dev. Biol. 2021, 9, 753706.
- Jia, J.; Li, F.; Tang, X.S.; Xu, S.; Gao, Y.; Shi, Q.; Guo, W.; Wang, X.; He, D.; Guo, P. Long noncoding RNA DANCR promotes invasion of prostate cancer through epigenetically silencing expression of TIMP2/3. Oncotarget 2016, 7, 37868–37881.
- Wang, Y.Y.; Chen, C. lncRNA-DANCR Promotes Taxol Resistance of Prostate Cancer Cells through Modulating the miR-33b-5p-LDHA Axis. Dis. Mrk. 2022, 2022, 9516774.
- Deng, H.; Zhu, B.; Dong, Z.; Jiang, H.; Zhao, X.; Wu, S. miR-214-5p targeted by LncRNA DANCR mediates TGF-β signaling pathway to accelerate proliferation, migration and inhibit apoptosis of prostate cancer cells. Am. J. Transl. Res. 2021, 13, 2224–2240.
- Jiang, N.; Wang, X.; Xie, X.; Liao, Y.; Liu, N.; Liu, J.; Miao, N.; Shen, J.; Peng, T. lncRNA DANCR promotes tumor progression and cancer stemness features in osteosarcoma by upregulating AXL via miR-33a-5p inhibition. Cancer Lett. 2017, 405, 46–55.
- Li, Z.X.; Zhu, Q.N.; Zhang, H.B.; Hu, Y.; Wang, G.; Zhu, Y.S. MALAT1: A potential biomarker in cancer. Cancer Manag. Res. 2018, 10, 6757–6768.
- Zhang, J.; Hao, X.; Yin, M.; Xu, T.; Guo, F. Long non-coding RNA in osteogenesis: A new world to be explored. Bone Jt. Res. 2019, 8, 73–80.
- Deng, J.; Tang, J.; Wang, G.; Zhu, Y.S. Long Non-Coding RNA as Potential Biomarker for Prostate Cancer: Is It Making a Difference? Int. J. Environ. Res. Public Health 2017, 14, 270.
- Ren, S.; Liu, Y.; Xu, W.; Sun, Y.; Lu, J.; Wang, F.; Wei, M.; Shen, J.; Hou, J.; Gao, X.; et al. Long noncoding RNA MALAT-1 is a new potential therapeutic target for castration resistant prostate cancer. J. Urol. 2013, 190, 2278–2287.
- Yu, X.; Li, Z.; Zheng, H.; Chan, M.T.; Wu, W.K. NEAT1: A novel cancer-related long non-coding RNA. Cell Prolif. 2017, 50, e12329.
- Wen, S.; Wei, Y.; Zen, C.; Xiong, W.; Niu, Y.; Zhao, Y. Long non-coding RNA NEAT1 promotes bone metastasis of prostate cancer through N6-methyladenosine. Mol. Cancer 2020, 19, 171.
- Zhao, W.; Zhu, X.; Jin, Q.; Lin, B.; Ji, R. The lncRNA NEAT1/miRNA-766-5p/E2F3 Regulatory Axis Promotes Prostate Cancer Progression. J. Oncol. 2022, 2022, 1866972.
- Hajjari, M.; Salavaty, A. HOTAIR: An oncogenic long non-coding RNA in different cancers. Cancer Biol. Med. 2015, 12, 1–9.
- Li, T.; Liu, N.; Gao, Y.; Quan, Z.; Hao, Y.; Yu, C.; Li, L.; Yuan, M.; Niu, L.; Luo, C.; et al. Long noncoding RNA HOTAIR regulates the invasion and metastasis of prostate cancer by targeting hepaCAM. Br. J. Cancer 2021, 124, 247–258.
- Wang, B.; Qu, X.L.; Liu, J.; Lu, J.; Zhou, Z.Y. HOTAIR promotes osteosarcoma development by sponging miR-217 and targeting ZEB1. J. Cell. Physiol. 2019, 234, 6173–6181.
- Zheng, H.; Min, J. Role of Long Noncoding RNA HOTAIR in the Growth and Apoptosis of Osteosarcoma Cell MG-63. BioMed Res. Int. 2016, 2016, 5757641.
- Guo, J.; Dou, D.; Zhang, T.; Wang, B. HOTAIR Promotes Cisplatin Resistance of Osteosarcoma Cells by Regulating Cell Proliferation, Invasion, and Apoptosis via miR-106a-5p/STAT3 Axis. Cell Transplant. 2020, 29, 963689720948447.
- Li, Z.; Shen, J.; Chan, M.T.; Wu, W.K. TUG1: A pivotal oncogenic long non-coding RNA of human cancers. Cell Prolif. 2016, 49, 471–475.
- Yang, B.; Tang, X.; Wang, Z.; Sun, D.; Wei, X.; Ding, Y. TUG1 promotes prostate cancer progression by acting as a ceRNA of miR-26a. Biosci. Rep. 2018, 38, BSR20180677.
- Sheng, K.; Li, Y. LncRNA TUG1 promotes the development of osteosarcoma through RUNX2. Exp. Ther. Med. 2019, 18, 3002–3008.
- Zhou, Q.; Hu, T.; Xu, Y. Anticancer potential of TUG1 knockdown in cisplatin-resistant osteosarcoma through inhibition of MET/Akt signalling. J. Drug Target. 2020, 28, 204–211.
- Yu, X.; Hu, L.; Li, S.; Shen, J.; Wang, D.; Xu, R.; Yang, H. Long non-coding RNA Taurine upregulated gene 1 promotes osteosarcoma cell metastasis by mediating HIF-1α via miR-143-5p. Cell Death Dis. 2019, 10, 280.
- Li, G.; Liu, K.; Du, X. Long Non-Coding RNA TUG1 Promotes Proliferation and Inhibits Apoptosis of Osteosarcoma Cells by Sponging miR-132-3p and Upregulating SOX4 Expression. Yonsei Med. J. 2018, 59, 226–235.
- Ghafouri-Fard, S.; Dashti, S.; Taheri, M. PCAT1: An oncogenic lncRNA in diverse cancers and a putative therapeutic target. Exp. Mol. Pathol. 2020, 114, 104429.
- Shang, Z.; Yu, J.; Sun, L.; Tian, J.; Zhu, S.; Zhang, B.; Dong, Q.; Jiang, N.; Flores-Morales, A.; Chang, C.; et al. LncRNA PCAT1 activates AKT and NF-κB signaling in castration-resistant prostate cancer by regulating the PHLPP/FKBP51/IKKα complex. Nucleic Acids Res. 2019, 47, 4211–4225.
- Zhang, X.; Zhang, Y.; Mao, Y.; Ma, X. The lncRNA PCAT1 is correlated with poor prognosis and promotes cell proliferation, invasion, migration and EMT in osteosarcoma. OncoTargets Ther. 2018, 11, 629–638.
- Chang, L.; Jia, D.L.; Cao, C.S.; Wei, H.; Li, Z.Q. LncRNA PCAT-1 promotes the progression of osteosarcoma via miR-508-3p/ZEB1 axis. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 2517–2527.
- Tang, F.; Xu, Y.; Wang, H.; Bian, E.; Zhao, B. LncRNA-ATB in cancers: What do we know so far? Mol. Biol. Rep. 2020, 47, 4077–4086.
- Xu, S.; Yi, X.M.; Tang, C.P.; Ge, J.P.; Zhang, Z.Y.; Zhou, W.Q. Long non-coding RNA ATB promotes growth and epithelial-mesenchymal transition and predicts poor prognosis in human prostate carcinoma. Oncol. Rep. 2016, 36, 10–22.
- Onagoruwa, O.T.; Pal, G.; Ochu, C.; Ogunwobi, O.O. Oncogenic Role of PVT1 and Therapeutic Implications. Front. Oncol. 2020, 10, 17.
- Wang, W.; Zhou, R.; Wu, Y.; Liu, Y.; Su, W.; Xiong, W.; Zeng, Z. PVT1 Promotes Cancer Progression via MicroRNAs. Front. Oncol. 2019, 9, 609.
- Wu, H.; Tian, X.; Zhu, C. Knockdown of lncRNA PVT1 inhibits prostate cancer progression in vitro and in vivo by the suppression of KIF23 through stimulating miR-15a-5p. Cancer Cell Int. 2020, 20, 283.
- Liu, J.; Li, Y.; Zhang, Q.; Lv, C.; Wang, M.; Jiao, Y.; Wang, C. PVT1 Expression Is a Predictor for Poor Survival of Prostate Cancer Patients. Technol. Cancer Res. Treat. 2021, 20, 1533033820971610.
- Sun, F.; Wu, K.; Yao, Z.; Mu, X.; Zheng, Z.; Sun, M.; Wang, Y.; Liu, Z.; Zhu, Y. Long Noncoding RNA PVT1 Promotes Prostate Cancer Metastasis by Increasing NOP2 Expression via Targeting Tumor Suppressor MicroRNAs. OncoTargets Ther. 2020, 13, 6755–6765.
- Yan, M.; Pan, X.F.; Liu, Y.; Zhao, S.; Gong, W.Q.; Liu, W. Long noncoding RNA PVT1 promotes metastasis via miR-484 sponging in osteosarcoma cells. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 2229–2238.
- Xun, C.; Jiang, D.; Tian, Z.; Yunus, A.; Chen, J. Long noncoding RNA plasmacytoma variant translocation gene 1 promotes epithelial-mesenchymal transition in osteosarcoma. J. Clin. Lab. Anal. 2021, 35, e23587.

