 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Isa Domingues Serrano | -- | 4375 | 2023-03-10 12:00:28 | | | |
| 2 | Lindsay Dong | Meta information modification | 4375 | 2023-03-13 02:26:13 | | |
Video Upload Options
The first research on the insect Galleria mellonella was published 85 years ago, and the larva is now widely used as a model to study infections caused by bacterial and fungal pathogens, for screening new antimicrobials, to study the adjacent immune response in co-infections or in host-pathogen interaction, as well as in a toxicity model. The immune system of the G. mellonella model shows remarkable similarities with mammals. Furthermore, results from G. mellonella correlate positively with mammalian models and with other invertebrate models. Unlike other invertebrate models, G. mellonella can withstand temperatures of 37 °C, and its handling and experimental procedures are simpler. Despite having some disadvantages, G. mellonella is a virtuous in vivo model to be used in preclinical studies, as an intermediate model between in vitro and mammalian in vivo studies, and is a great example on how to apply the bioethics principle of the 3Rs (Replacement, Reduction, and Refinement) in animal experimentation.
1. Introduction
2. Characterization of Galleria mellonella
3. Immune System of Galleria mellonella
3.1. Opsonins
3.2. Antimicrobial Peptides
3.3. Melanine
3.4. Extracellular Nucleic Acid
3.5. Influence of Diet on Larval Immune Health
4. Galleria mellonella In Vivo Model—Experimental Design Considerations
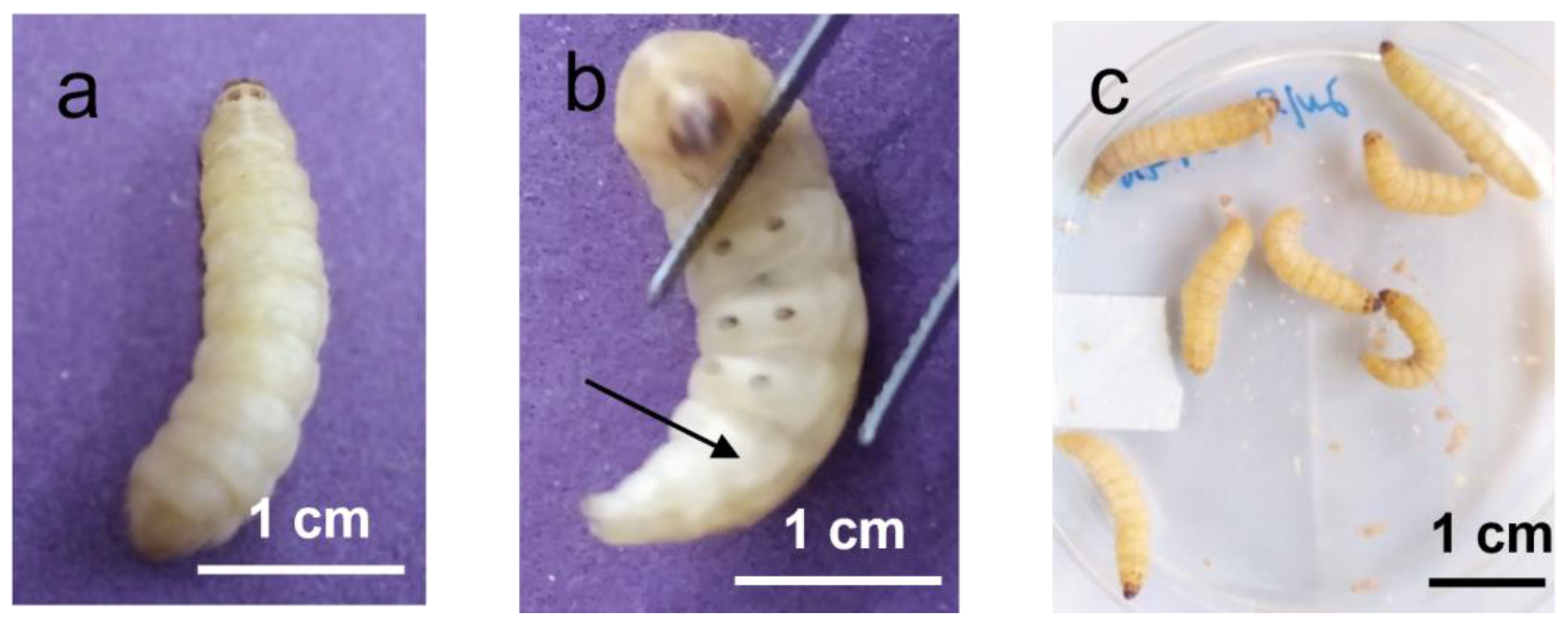
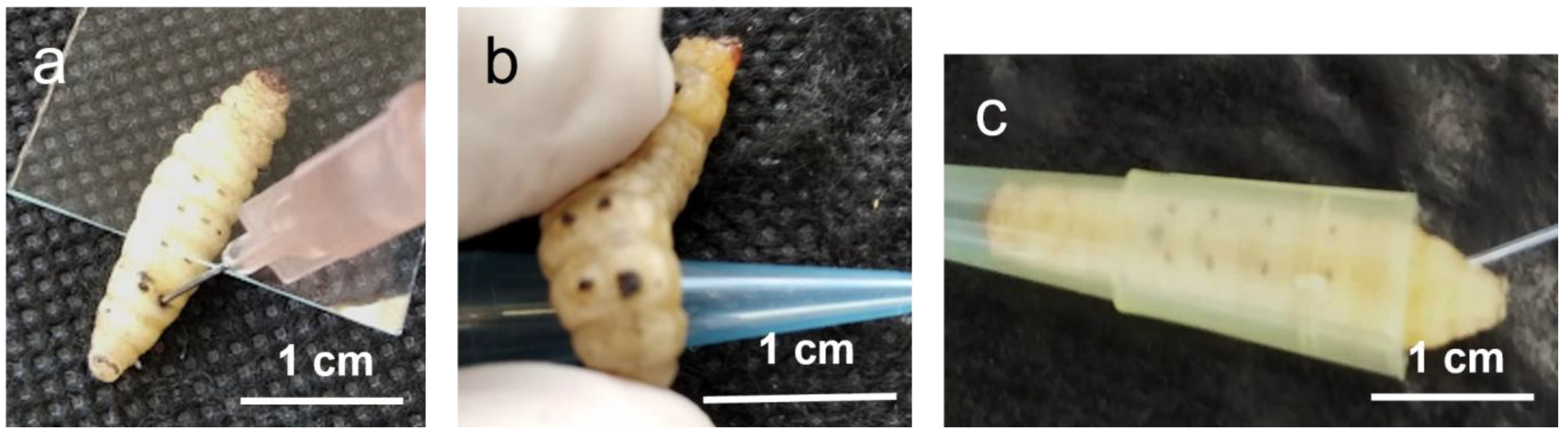
5. Gram-Positive and Gram-Negative Bacteria Causing Disease in Humans
6. Comparison with Other Invertebrate and Mammalian Models
7. Conclusions
References
- Jorjão, A.L.; Oliveira, L.D.; Scorzoni, L.; Figueiredo-Godoi, L.M.A.; Cristina A Prata, M.; Jorge, A.O.C.; Junqueira, J.C. From moths to caterpillars: Ideal conditions for Galleria mellonella rearing for in vivo microbiological studies. Virulence 2018, 9, 383–389.
- Pereira, M.F.; Rossi, C.C.; da Silva, G.C.; Rosa, J.N.; Bazzolli, D.M.S. Galleria mellonella as an infection model: An in-depth look at why it works and practical considerations for successful application. Pathog. Dis. 2020, 78, ftaa056.
- Tsai, C.J.; Loh, J.M.; Proft, T. Galleria mellonella infection models for the study of bacterial diseases and for antimicrobial drug testing. Virulence 2016, 7, 214–229.
- Cutuli, M.A.; Petronio Petronio, G.; Vergalito, F.; Magnifico, I.; Pietrangelo, L.; Venditti, N.; Di Marco, R. Galleria mellonella as a consolidated in vivo model hosts: New developments in antibacterial strategies and novel drug testing. Virulence 2019, 10, 527–541.
- Smith, T.L. Genetical Studies on the Wax Moth Galleria mellonella Linn. Genetics 1938, 23, 115–137.
- Ingrao, F.; Belli, N. Galleria mellonella test in the study of the antituberculosis activity of thiosemicarbazones. Lotta Contro La Tuberc. 1951, 21, 282–287.
- Morellini, M.; Avegno, R.P. Antimycotic drugs evaluated with the Galleria mellonella test. Ann. Ist. Carlo Forlanini 1957, 17, 149–154.
- Smissman, E.E.; Beck, S.D.; Boots, M.R. Growth Inhibition of Insects and a Fungus by Indole-3-Acetonitrile. Science 1961, 133, 462.
- Kurstak, E.; Vega, C.E. Bacterial infection due to Salmonella typhimurium in an invertebrate, Galleria mellonella L. Can. J. Microbiol. 1968, 14, 233–237.
- Fuchs, B.B.; Mylonakis, E. Using non-mammalian hosts to study fungal virulence and host defense. Curr. Opin. Microbiol. 2006, 9, 346–351.
- Younghusband, H.B.; Lee, P.E. Virus-cell studies of tipula iridescent virus in Galleria mellonella L. I. Electron microscopy of infection and synthesis of tipula iridescent virus in hemocytes. Virology 1969, 38, 247–254.
- Garzon, S.; Charpentier, G.; Kurstak, E. Morphogenesis of the nodamura virus in the larbae of the lepidopteran Galleria mellonella (L.). Arch. Virol. 1978, 56, 61–76.
- Fargues, J.; Bon, M.C. Influence of temperature preferences of two Paecilomyces fumosoroseus lineages on their co-infection pattern. J. Invertebr. Pathol. 2004, 87, 94–104.
- Lange, A.; Beier, S.; Huson, D.H.; Parusel, R.; Iglauer, F.; Frick, J.S. Genome Sequence of Galleria mellonella (Greater Wax Moth). Genome Announc. 2018, 6, e01220-17.
- Heitmueller, M.; Billion, A.; Dobrindt, U.; Vilcinskas, A.; Mukherjee, K. Epigenetic Mechanisms Regulate Innate Immunity against Uropathogenic and Commensal-Like Escherichia coli in the Surrogate Insect Model Galleria mellonella. Infect. Immun. 2017, 85, e00336-17.
- Sheehan, G.; Kavanagh, K. Proteomic analysis of the responses of candida albicans during infection of Galleria mellonella larvae. J. Fungi 2019, 5, 7.
- Mukherjee, K.; Vilcinskas, A. Development and immunity-related microRNAs of the lepidopteran model host Galleria mellonella. BMC Genom. 2014, 15, 705.
- Champion, O.L.; Wagley, S.; Titball, R.W. Galleria mellonella as a model host for microbiological and toxin research. Virulence 2016, 7, 840–845.
- Scoble, M.J. The Lepidoptera: Form, Function, and Diversity; Oxford University Press: Oxford, UK, 1992; p. 404.
- Kwadha, C.A.; Ong’amo, G.O.; Ndegwa, P.N.; Raina, S.K.; Fombong, A.T. The Biology and Control of the Greater Wax Moth, Galleria mellonella. Insects 2017, 8, 61.
- Gulati, R.; Kaushik, H.D. Enemies of honeybees and their management—A review. Agric. Rev. 2004, 25, 189–200.
- Smith, T.L. External Morphology of the Larva, Pupa, and Adult of the Wax Moth, Galleria Mellonella L. J. Kans. Entomol. Soc. 1965, 38, 287–310.
- Ellis, J.; Graham, J.; Mortensen, A. Standard methods for wax moth research. J. Apic. Res. 2013, 52, 1–17.
- Nielsen, R.A.; Brister, C.D. Greater Wax Moth: Behavior of Larvae. Ann. Entomol. Soc. Am. 1979, 726, 811–815.
- Charrière, J.-D.; Imdorf, A. Protection of honey combs from wax moth damage. Am. Bee J. 1999, 139, 627–630.
- Singkum, P.; Suwanmanee, S.; Pumeesat, P.; Luplertlop, N. A powerful in vivo alternative model in scientific research: Galleria mellonella. Acta Microbiol. Immunol. Hung. 2019, 66, 31–55.
- Pereira, M.F.; Rossi, C.C.; Vieira de Queiroz, M.; Martins, G.F.; Isaac, C.; Bossé, J.T.; Li, Y.; Wren, B.W.; Terra, V.S.; Cuccui, J.; et al. Galleria mellonella is an effective model to study Actinobacillus pleuropneumoniae infection. Microbiology 2015, 161, 387–400.
- Hillyer, J.F.; Pass, G. The Insect Circulatory System: Structure, Function, and Evolution. Annu. Rev. Entomol. 2020, 65, 121–143.
- Durieux, M.F.; Melloul, É.; Jemel, S.; Roisin, L.; Dardé, M.L.; Guillot, J.; Dannaoui, É.; Botterel, F. Galleria mellonella as a screening tool to study virulence factors of Aspergillus fumigatus. Virulence 2021, 12, 818–834.
- Allonsius, C.N.; Van Beeck, W.; De Boeck, I.; Wittouck, S.; Lebeer, S. The microbiome of the invertebrate model host Galleria mellonella is dominated by Enterococcus. Anim. Microbiome 2019, 1, 7.
- Ménard, G.; Rouillon, A.; Cattoir, V.; Donnio, P.Y. Galleria mellonella as a Suitable Model of Bacterial Infection: Past, Present and Future. Front. Cell Infect. Microbiol. 2021, 11, 782733.
- Gohl, P.; LeMoine, C.M.R.; Cassone, B.J. Diet and ontogeny drastically alter the larval microbiome of the invertebrate model. Can. J. Microbiol. 2022, 68, 594–604.
- Lemaitre, B.; Hoffmann, J. The host defense of Drosophila melanogaster. Annu. Rev. Immunol. 2007, 25, 697–743.
- Sheehan, G.; Garvey, A.; Croke, M.; Kavanagh, K. Innate humoral immune defences in mammals and insects: The same, with differences ? Virulence 2018, 9, 1625–1639.
- Kavanagh, K.; Sheehan, G. The use of Galleria mellonella larvae to identify novel antimicrobial agents against fungal species of medical interest. J. Fungi 2018, 4, 113.
- Tojo, S.; Naganuma, F.; Arakawa, K.; Yokoo, S. Involvement of both granular cells and plasmatocytes in phagocytic reactions in the greater wax moth, Galleria mellonella. J. Insect. Physiol. 2000, 46, 1129–1135.
- Lavine, M.D.; Strand, M.R. Insect hemocytes and their role in immunity. Insect Biochem. Mol. Biol. 2002, 32, 1295–1309.
- Browne, N.; Heelan, M.; Kavanagh, K. An analysis of the structural and functional similarities of insect hemocytes and mammalian phagocytes. Virulence 2013, 4, 597–603.
- Arteaga Blanco, L.A.; Crispim, J.S.; Fernandes, K.M.; de Oliveira, L.L.; Pereira, M.F.; Bazzolli, D.M.S.; Martins, G.F. Differential cellular immune response of Galleria mellonella to Actinobacillus pleuropneumoniae. Cell Tissue Res. 2017, 370, 153–168.
- Kordaczuk, J.; Sułek, M.; Mak, P.; Zdybicka-Barabas, A.; Śmiałek, J.; Wojda, I. Cationic protein 8 plays multiple roles in Galleria mellonella immunity. Sci. Rep. 2022, 12, 11737.
- Wu, G.; Liu, Y.; Ding, Y.; Yi, Y. Ultrastructural and functional characterization of circulating hemocytes from Galleria mellonella larva: Cell types and their role in the innate immunity. Tissue Cell 2016, 48, 297–304.
- Pech, L.L.; Strand, M.R. Granular cells are required for encapsulation of foreign targets by insect haemocytes. J. Cell Sci. 1996, 109, 2053–2060.
- Schmit, A.R.; Ratcliffe, N.A. The encapsulation of foreign tissue implants in Galleria mellonella larvae. J. Insect Physiol. 1977, 23, 175–184.
- Insua, J.L.; Llobet, E.; Moranta, D.; Pérez-Gutiérrez, C.; Tomás, A.; Garmendia, J.; Bengoechea, J.A. Modeling Klebsiella pneumoniae pathogenesis by infection of the wax moth Galleria mellonella. Infect. Immun. 2013, 81, 3552–3565.
- Barnoy, S.; Gancz, H.; Zhu, Y.; Honnold, C.L.; Zurawski, D.V.; Venkatesan, M.M. The Galleria mellonella larvae as an in vivo model for evaluation of Shigella virulence. Gut Microbes 2017, 8, 335–350.
- Kim, C.H.; Shin, Y.P.; Noh, M.Y.; Jo, Y.H.; Han, Y.S.; Seong, Y.S.; Lee, I.H. An insect multiligand recognition protein functions as an opsonin for the phagocytosis of microorganisms. J. Biol. Chem. 2010, 285, 25243–25250.
- Wang, Q.; Ren, M.; Liu, X.; Xia, H.; Chen, K. Peptidoglycan recognition proteins in insect immunity. Mol. Immunol. 2019, 106, 69–76.
- Park, S.Y.; Kim, C.H.; Jeong, W.H.; Lee, J.H.; Seo, S.J.; Han, Y.S.; Lee, I.H. Effects of two hemolymph proteins on humoral defense reactions in the wax moth, Galleria mellonella. Dev. Comp. Immunol. 2005, 29, 43–51.
- Zdybicka-Barabas, A.; Stączek, S.; Mak, P.; Skrzypiec, K.; Mendyk, E.; Cytryńska, M. Synergistic action of Galleria mellonella apolipophorin III and lysozyme against Gram-negative bacteria. Biochim. Biophys. Acta 2013, 1828, 1449–1456.
- Stączek, S.; Zdybicka-Barabas, A.; Mak, P.; Sowa-Jasiłek, A.; Kedracka-Krok, S.; Jankowska, U.; Suder, P.; Wydrych, J.; Grygorczuk, K.; Jakubowicz, T.; et al. Studies on localization and protein ligands of Galleria mellonella apolipophorin III during immune response against different pathogens. J. Insect Physiol. 2018, 105, 18–27.
- Riddell, D.R.; Graham, A.; Owen, J.S. Apolipoprotein E inhibits platelet aggregation through the L-arginine:nitric oxide pathway. Implications for vascular disease. J. Biol. Chem. 1997, 272, 89–95.
- Yu, X.Q.; Kanost, M.R. Binding of hemolin to bacterial lipopolysaccharide and lipoteichoic acid. An immunoglobulin superfamily member from insects as a pattern-recognition receptor. Eur. J. Biochem. 2002, 269, 1827–1834.
- Shaik, H.A.; Sehnal, F. Hemolin expression in the silk glands of Galleria mellonella in response to bacterial challenge and prior to cell disintegration. J. Insect Physiol. 2009, 55, 781–787.
- Mowlds, P.; Coates, C.; Renwick, J.; Kavanagh, K. Dose-dependent cellular and humoral responses in Galleria mellonella larvae following beta-glucan inoculation. Microbes Infect. 2010, 12, 146–153.
- Toke, O. Antimicrobial peptides: New candidates in the fight against bacterial infections. Biopolymers 2005, 80, 717–735.
- Diamond, G.; Beckloff, N.; Weinberg, A.; Kisich, K.O. The roles of antimicrobial peptides in innate host defense. Curr. Pharm. Des. 2009, 15, 2377–2392.
- Serrano, I. Antimicrobial Peptides. In Frontiers in Antimicrobial Agents—The Challenging of Antibiotic Resistance in the Development of New Therapeutics; Oliveira, M., Serrano, I., Eds.; Bentham Science Publishers: Sharjah, United Arab Emirates, 2015; Volume 1, pp. 33–68.
- Brown, S.E.; Howard, A.; Kasprzak, A.B.; Gordon, K.H.; East, P.D. The discovery and analysis of a diverged family of novel antifungal moricin-like peptides in the wax moth Galleria mellonella. Insect Biochem. Mol. Biol. 2008, 38, 201–212.
- Hoffmann, J.A.; Reichhart, J.M.; Hetru, C. Innate immunity in higher insects. Curr. Opin. Immunol. 1996, 8, 8–13.
- Cytryńska, M.; Mak, P.; Zdybicka-Barabas, A.; Suder, P.; Jakubowicz, T. Purification and characterization of eight peptides from Galleria mellonella immune hemolymph. Peptides 2007, 28, 533–546.
- Zitzmann, J.; Weidner, T.; Czermak, P. Optimized expression of the antimicrobial protein Gloverin from Galleria mellonella using stably transformed Drosophila melanogaster S2 cells. Cytotechnology 2017, 69, 371–389.
- Sowa-Jasiłek, A.; Zdybicka-Barabas, A.; Stączek, S.; Wydrych, J.; Skrzypiec, K.; Mak, P.; Deryło, K.; Tchórzewski, M.; Cytryńska, M. Galleria mellonella lysozyme induces apoptotic changes in Candida albicans cells. Microbiol. Res. 2016, 193, 121–131.
- Brown, S.E.; Howard, A.; Kasprzak, A.B.; Gordon, K.H.; East, P.D. A peptidomics study reveals the impressive antimicrobial peptide arsenal of the wax moth Galleria mellonella. Insect Biochem. Mol. Biol. 2009, 39, 792–800.
- Lu, A.; Zhang, Q.; Zhang, J.; Yang, B.; Wu, K.; Xie, W.; Luan, Y.X.; Ling, E. Insect prophenoloxidase: The view beyond immunity. Front. Physiol. 2014, 5, 252.
- Altincicek, B.; Stötzel, S.; Wygrecka, M.; Preissner, K.T.; Vilcinskas, A. Host-derived extracellular nucleic acids enhance innate immune responses, induce coagulation, and prolong survival upon infection in insects. J. Immunol. 2008, 181, 2705–2712.
- Banville, N.; Browne, N.; Kavanagh, K. Effect of nutrient deprivation on the susceptibility of Galleria mellonella larvae to infection. Virulence 2012, 3, 497–503.
- Champion, O.L.; Titball, R.W.; Bates, S. Standardization of G. mellonella Larvae to Provide Reliable and Reproducible Results in the Study of Fungal Pathogens. J. Fungi 2018, 4, 108.
- Harding, C.; Schroeder, G.; Collins, J.; Frankel, G. Use of Galleria mellonella as a Model Organism to Study Legionella pneumophila Infection. J. Vis. Exp. JoVE 2013, 22, e50964.
- Ramarao, N.; Nielsen-Leroux, C.; Lereclus, D. The insect Galleria mellonella as a powerful infection model to investigate bacterial pathogenesis. J. Vis. Exp. 2012, 11, e4392.
- Desbois, A.P.; Coote, P.J. Wax moth larva (Galleria mellonella): An in vivo model for assessing the efficacy of antistaphylococcal agents. J. Antimicrob. Chemother. 2011, 66, 1785–1790.
- Mil-Homens, D.; Barahona, S.; Moreira, R.N.; Silva, I.J.; Pinto, S.N.; Fialho, A.M.; Arraiano, C.M. Stress Response Protein BolA Influences Fitness and Promotes Salmonella enterica Serovar Typhimurium Virulence. Appl. Environ. Microbiol. 2018, 84, e02850-17.
- Cotter, G.; Doyle, S.; Kavanagh, K. Development of an insect model for the in vivo pathogenicity testing of yeasts. FEMS Immunol. Med. Microbiol. 2000, 27, 163–169.
- Fredericks, L.R.; Lee, M.D.; Roslund, C.R.; Crabtree, A.M.; Allen, P.B.; Rowley, P.A. The design and implementation of restraint devices for the injection of pathogenic microorganisms into Galleria mellonella. PLoS ONE 2020, 15, e0230767.
- Bismuth, H.; Aussel, L.; Ezraty, B. The greater wax moth, Galleria mellonella to study host-pathogen interactions. Med. Sci. 2019, 35, 346–351.
- Konkel, M.E.; Tilly, K. Temperature-regulated expression of bacterial virulence genes. Microbes Infect. 2000, 2, 157–166.
- Peters, B.M.; Jabra-Rizk, M.A.; O’May, G.A.; Costerton, J.W.; Shirtliff, M.E. Polymicrobial interactions: Impact on pathogenesis and human disease. Clin. Microbiol. Rev. 2012, 25, 193–213.
- Whiley, R.A.; Sheikh, N.P.; Mushtaq, N.; Hagi-Pavli, E.; Personne, Y.; Javaid, D.; Waite, R.D. Differential potentiation of the virulence of the Pseudomonas aeruginosa cystic fibrosis liverpool epidemic strain by oral commensal Streptococci. J. Infect. Dis. 2014, 209, 769–780.
- Scott, J.; Sueiro-Olivares, M.; Ahmed, W.; Heddergott, C.; Zhao, C.; Thomas, R.; Bromley, M.; Latgé, J.P.; Krappmann, S.; Fowler, S.; et al. Pseudomonas aeruginosa-Derived Volatile Sulfur Compounds Promote Distal Aspergillus fumigatus Growth and a Synergistic Pathogen-Pathogen Interaction That Increases Pathogenicity in Co-infection. Front. Microbiol. 2019, 10, 2311.
- Reece, E.; Doyle, S.; Greally, P.; Renwick, J.; McClean, S. Aspergillus fumigatus inhibits Pseudomonas aeruginosa in co-culture: Implications of a mutually antagonistic relationship on virulence and inflammation in the CF airway. Front. Microbiol. 2018, 9, 1205.
- Mukherjee, K.; Domann, E.; Hain, T. The Greater Wax Moth Galleria mellonella as an Alternative Model Host for Human Pathogens; Springer Science+Business Media: Berlin, Germany, 2010; Volume 2, pp. 3–14.
- Harding, C.R.; Schroeder, G.N.; Reynolds, S.; Kosta, A.; Collins, J.W.; Mousnier, A.; Frankel, G. Legionella pneumophila pathogenesis in the Galleria mellonella infection model. Infect. Immun. 2012, 80, 2780–2790.
- Lionakis, M.S. Drosophila and Galleria insect model hosts: New tools for the study of fungal virulence, pharmacology and immunology. Virulence 2011, 2, 521–527.
- Piatek, M.; Sheehan, G.; Kavanagh, K. Galleria mellonella: The Versatile Host for Drug Discovery, In Vivo Toxicity Testing and Characterising Host-Pathogen Interactions. Antibiotics 2021, 10, 1545.
- Jander, G.; Rahme, L.G.; Ausubel, F.M. Positive correlation between virulence of Pseudomonas aeruginosa mutants in mice and insects. J. Bacteriol. 2000, 182, 3843–3845.
- Eisemann, C.; Jorgensen, W.; Merritt, D.; Rice, M.; Cribb, B.; Webb, P.; Zalucki, M. Do insects feel pain?—A biological view. Experientia 1984, 40, 164–167.


