 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Marilena Pariano | -- | 2148 | 2023-03-06 16:34:21 | | | |
| 2 | Lindsay Dong | Meta information modification | 2148 | 2023-03-07 09:54:19 | | |
Video Upload Options
Developing therapeutics for inflammatory diseases is challenging due to physiological mucosal barriers, systemic side effects, and the local microbiota. In the search for novel methods to overcome some of these problems, drug delivery systems that improve tissue-targeted drug delivery and modulate the microbiota are highly desirable. Microbial metabolites are known to regulate immune responses, an observation that has resulted in important conceptual advances in areas such as metabolite pharmacology and metabolite therapeutics. Indeed, the doctrine of “one molecule, one target, one disease” that has dominated the pharmaceutical industry in the 20th century is being replaced by developing therapeutics which simultaneously manipulate multiple targets through novel formulation approaches, including the multitarget-directed ligands. Thus, metabolites may not only represent biomarkers for disease development, but also, being causally linked to human diseases, an unexploited source of therapeutics.
1. Introduction
2. Metabolite-Based Therapeutics: The Postbiotic Concept
3. Tissue-Specific Drug Delivery
The field of drug delivery systems, including routes of delivery and delivery vehicles, has advanced dramatically in the past few decades, thanks to the increased understanding of the physiological barriers for efficient drug delivery and the development of several new modes of drug delivery that have entered clinical practice. These include lipid nanoparticles (essential for effective delivery of mRNA), the microneedle patch (for sustained delivery to low-income people or people with mental-health conditions), the robotic pill (for oral drug delivery of complex drugs), and cellular delivery via red blood cells or bacteria, to name a few [8]. However, despite improving the drug’s efficacy, side effects may still occur because the unwanted drugs interact with healthy organs or tissues. Thus, targeted drug delivery approaches are of great interest in the pharmaceutical sciences, as they enable the concentrated delivery of a drug compound to its desired target—increasing efficacy and reducing off-target effects as compared to conventional formulations [8]. DDS have enabled the development of many pharmaceutical products that improve patient health by enhancing the delivery of a therapeutic to its target site, minimizing off-target accumulation and facilitating patient compliance [9].
4. The Ubiquitous AhR Signaling Pathway
5. The Tryptophan-Indole Microbial Pathway
6. Turning Indoles into Therapeutics via Targeted Delivery Technologies
References
- Pascal Andreu, V.; Roel-Touris, J.; Dodd, D.; Fischbach, M.A.; Medema, M.H. The gutSMASH web server: Automated identification of primary metabolic gene clusters from the gut microbiota. Nucleic Acids Res. 2021, 49, W263–W270.
- Krautkramer, K.A.; Fan, J.; Backhed, F. Gut microbial metabolites as multi-kingdom intermediates. Nat. Rev. Microbiol. 2021, 19, 77–94.
- Gilbert, J.A.; Blaser, M.J.; Caporaso, J.G.; Jansson, J.K.; Lynch, S.V.; Knight, R. Current understanding of the human microbiome. Nat. Med. 2018, 24, 392–400.
- Hosseinkhani, F.; Heinken, A.; Thiele, I.; Lindenburg, P.W.; Harms, A.C.; Hankemeier, T. The contribution of gut bacterial metabolites in the human immune signaling pathway of non-communicable diseases. Gut Microbes 2021, 13, 1–22.
- Levy, M.; Blacher, E.; Elinav, E. Microbiome, metabolites and host immunity. Curr. Opin. Microbiol. 2017, 35, 8–15.
- Levy, M.; Kolodziejczyk, A.A.; Thaiss, C.A.; Elinav, E. Dysbiosis and the immune system. Nat. Rev. Immunol. 2017, 17, 219–232.
- Zheng, X.; Xie, G.; Zhao, A.; Zhao, L.; Yao, C.; Chiu, N.H.; Jia, W. The footprints of gut microbial–Mammalian co-metabolism. J. Proteome Res. 2011, 10, 5512–5522.
- Allen, T.M.; Cullis, P.R. Drug delivery systems: Entering the mainstream. Science 2004, 303, 1818–1822.
- May, M. Why drug delivery is the key to new medicines. Nat. Med. 2022, 28, 1100–1102.
- Rothhammer, V.; Quintana, F.J. The aryl hydrocarbon receptor: An environmental sensor integrating immune responses in health and disease. Nat. Rev. Immunol. 2019, 19, 184–197.
- Stockinger, B.; Meglio, P.D.; Gialitakis, M.; Duarte, J.H. The aryl hydrocarbon receptor: Multitasking in the immune system. Annu. Rev. Immunol. 2014, 32, 403–432.
- Dong, F.; Perdew, G.H. The aryl hydrocarbon receptor as a mediator of host-microbiota interplay. Gut Microbes 2020, 12, 1859812.
- Roager, H.M.; Licht, T.R. Microbial tryptophan catabolites in health and disease. Nat. Commun. 2018, 9, 3294.
- Medvedev, A.; Buneeva, O. Tryptophan Metabolites as Mediators of Microbiota-Gut-Brain Communication: Focus on Isatin. Front. Behav. Neurosci. 2022, 16, 922274.
- Liu, Y.; Hou, Y.; Wang, G.; Zheng, X.; Hao, H. Gut microbial metabolites of aromatic amino acids as signals in host–microbe interplay. Trends Endocrinol. Metab. 2020, 31, 818–834.
- Ye, X.; Li, H.; Anjum, K.; Zhong, X.; Miao, S.; Zheng, G.; Li, L. Dual role of indoles derived from intestinal microbiota on human health. Front. Immunol. 2022, 13, 903526.
- Yokoyama, Y.; Ichiki, T.; Yamakawa, T.; Tsuji, Y.; Kuronuma, K.; Takahashi, S.; Nakase, H. Impaired tryptophan metabolism in the gastrointestinal tract of patients with critical coronavirus disease 2019. Front. Med. 2022, 9, 941422.
- Su, X.; Gao, Y.; Yang, R. Gut Microbiota-Derived Tryptophan Metabolites Maintain Gut and Systemic Homeostasis. Cells 2022, 11, 2296.
- Hubbard, T.D.; Murray, I.A.; Bisson, W.H.; Lahoti, T.S.; Gowda, K.; Amin, S.G.; Perdew, G.H. Adaptation of the human aryl hydrocarbon receptor to sense microbiota-derived indoles. Sci. Rep. 2015, 5, 12689.
- Zelante, T.; Iannitti, R.G.; Cunha, C.; De Luca, A.; Giovannini, G.; Pieraccini, G.; Romani, L. Tryptophan catabolites from microbiota engage aryl hydrocarbon receptor and balance mucosal reactivity via interleukin-22. Immunity 2013, 39, 372–385.
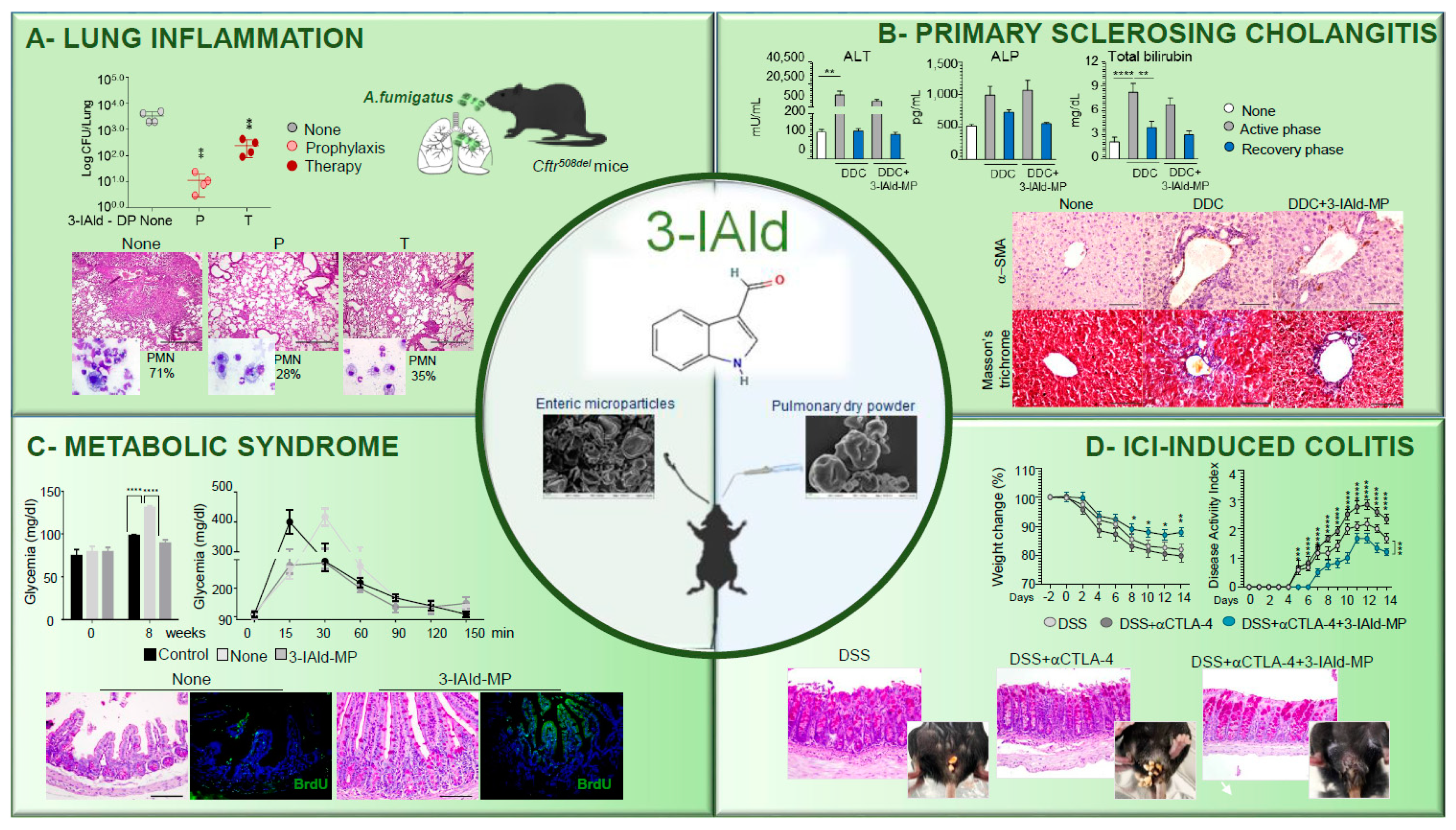
- Zelante, T.; Puccetti, M.; Giovagnoli, S.; Romani, L. Regulation of host physiology and immunity by microbial indole-3-aldehyde. Curr. Opin. Immunol. 2021, 70, 27–32.
- Puccetti, M.; Pariano, M.; Borghi, M.; Barola, C.; Moretti, S.; Galarini, R.; Giovagnoli, S. Enteric formulated indole-3-carboxaldehyde targets the aryl hydrocarbon receptor for protection in a murine model of metabolic syndrome. Int. J. Pharm. 2021, 602, 120610.
- Dvorak, Z.; Sokol, H.; Mani, S. Drug Mimicry: Promiscuous Receptors PXR and AhR, and Microbial Metabolite Interactions in the Intestine. Trends Pharm. Sci. 2020, 41, 900–908.
- Vyhlídalová, B.; Krasulová, K.; Pečinková, P.; Marcalíková, A.; Vrzal, R.; Zemánková, L.; Dvořák, Z. Gut microbial catabolites of tryptophan are ligands and agonists of the aryl hydrocarbon receptor: A detailed characterization. Int. J. Mol. Sci. 2020, 21, 2614.
- Puccetti, M.; Pariano, M.; Renga, G.; Santarelli, I.; D’Onofrio, F.; Bellet, M.M.; Giovagnoli, S. Targeted drug delivery technologies potentiate the overall therapeutic efficacy of an indole derivative in a mouse cystic fibrosis setting. Cells 2021, 10, 1601.
- Puccetti, M.; Giovagnoli, S.; Zelante, T.; Romani, L.; Ricci, M. Development of novel indole-3-aldehyde–loaded gastro-resistant spray-dried microparticles for postbiotic small intestine local delivery. J. Pharm. Sci. 2018, 107, 2341–2353.
- D’Onofrio, F.; Renga, G.; Puccetti, M.; Pariano, M.; Bellet, M.M.; Santarelli, I.; Romani, L. Indole-3-carboxaldehyde restores gut mucosal integrity and protects from liver fibrosis in murine sclerosing cholangitis. Cells 2021, 10, 1622.
- Renga, G.; Nunzi, E.; Pariano, M.; Puccetti, M.; Bellet, M.M.; Pieraccini, G.; Costantini, C. Optimizing therapeutic outcomes of immune checkpoint blockade by a microbial tryptophan metabolite. J. Immunother. Cancer 2022, 10, e003725.
- Puccetti, M.; Dos Reis, L.G.; Pariano, M.; Costantini, C.; Renga, G.; Ricci, M.; Giovagnoli, S. Development and in vitro-in vivo performances of an inhalable indole-3-carboxaldehyde dry powder to target pulmonary inflammation and infection. Int. J. Pharm. 2021, 607, 121004.
- Vesterhus, M.; Karlsen, T.H. Emerging therapies in primary sclerosing cholangitis: Pathophysiological basis and clinical opportunities. J. Gastroenterol. 2020, 55, 588–614.
- Natividad, J.M.; Agus, A.; Planchais, J.; Lamas, B.; Jarry, A.C.; Martin, R.; Sokol, H. Impaired aryl hydrocarbon receptor ligand production by the gut microbiota is a key factor in metabolic syndrome. Cell Metab. 2018, 28, 737–749.

