Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Xian-Ming Chen | -- | 2609 | 2023-02-24 02:33:23 | | | |
| 2 | Catherine Yang | Meta information modification | 2609 | 2023-02-24 02:39:11 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Deng, S.; Graham, M.L.; Chen, X. IFN-γ and Intracellular Protozoan Parasites. Encyclopedia. Available online: https://encyclopedia.pub/entry/41607 (accessed on 24 July 2026).
Deng S, Graham ML, Chen X. IFN-γ and Intracellular Protozoan Parasites. Encyclopedia. Available at: https://encyclopedia.pub/entry/41607. Accessed July 24, 2026.
Deng, Silu, Marion L. Graham, Xian-Ming Chen. "IFN-γ and Intracellular Protozoan Parasites" Encyclopedia, https://encyclopedia.pub/entry/41607 (accessed July 24, 2026).
Deng, S., Graham, M.L., & Chen, X. (2023, February 24). IFN-γ and Intracellular Protozoan Parasites. In Encyclopedia. https://encyclopedia.pub/entry/41607
Deng, Silu, et al. "IFN-γ and Intracellular Protozoan Parasites." Encyclopedia. Web. 24 February, 2023.
Copy Citation
IFN-γ is an important cytokine in both innate and adaptive immune responses during intracellular parasite infections. Elevated levels of IFN-γ is detected in both experimental animals and human patients following intracellular protozoal infections. An extensive number of studies support a protective role for IFN-γ against the infection of intracellular protozoan parasites, including Plasmodium, Toxoplasma, Cryptosporidium, Trypanosoma, Leishmania, while a few studies also indicated that IFN-γ contributes to the pathogenesis of parasite infection.
protozoan parasites
host defense
interferon
pathogenesis
signaling pathway
1. IFN-γ Production in Protozoan Parasite Infection
IFN-γ production in intracellular protozoan-infected hosts is predominantly mediated by NK cells [1][2][3][4][5], and CD4+/CD8+ and Ag-specific T cells [6][7][8][9][10] in innate and adaptive immunity, respectively. Other types of immune cells have been reported to produce IFN-γ during protozoal infection (Figure 1); for example, natural killer T (NKT) cells were shown to secrete IFN-γ in the liver of P. yeolii-infected mice [11][12] and a significant proportion of γδ T cells and αβ T cells were reported to produce IFN-γ in the peripheral blood of Plasmodium-infected children [13][14][15]. CD11b+ CD45low microglia and CD11b+ CD45high blood-derived macrophages were identified as the major non-T, non-NK cells expressing IFN-γ in the brain of T. gondii-infected mice, whereas group 1 innate lymphoid cells (ILC1s) were identified to produce IFN-γ in the small intestine in response to the oral infection of T. gondii in addition to NK cells and T cells [16][17][18]. The production of IFN-γ by immune cells can be negatively regulated by anti-inflammatory Th2 cytokines such as IL-10 and IL-4 [19][20][21].
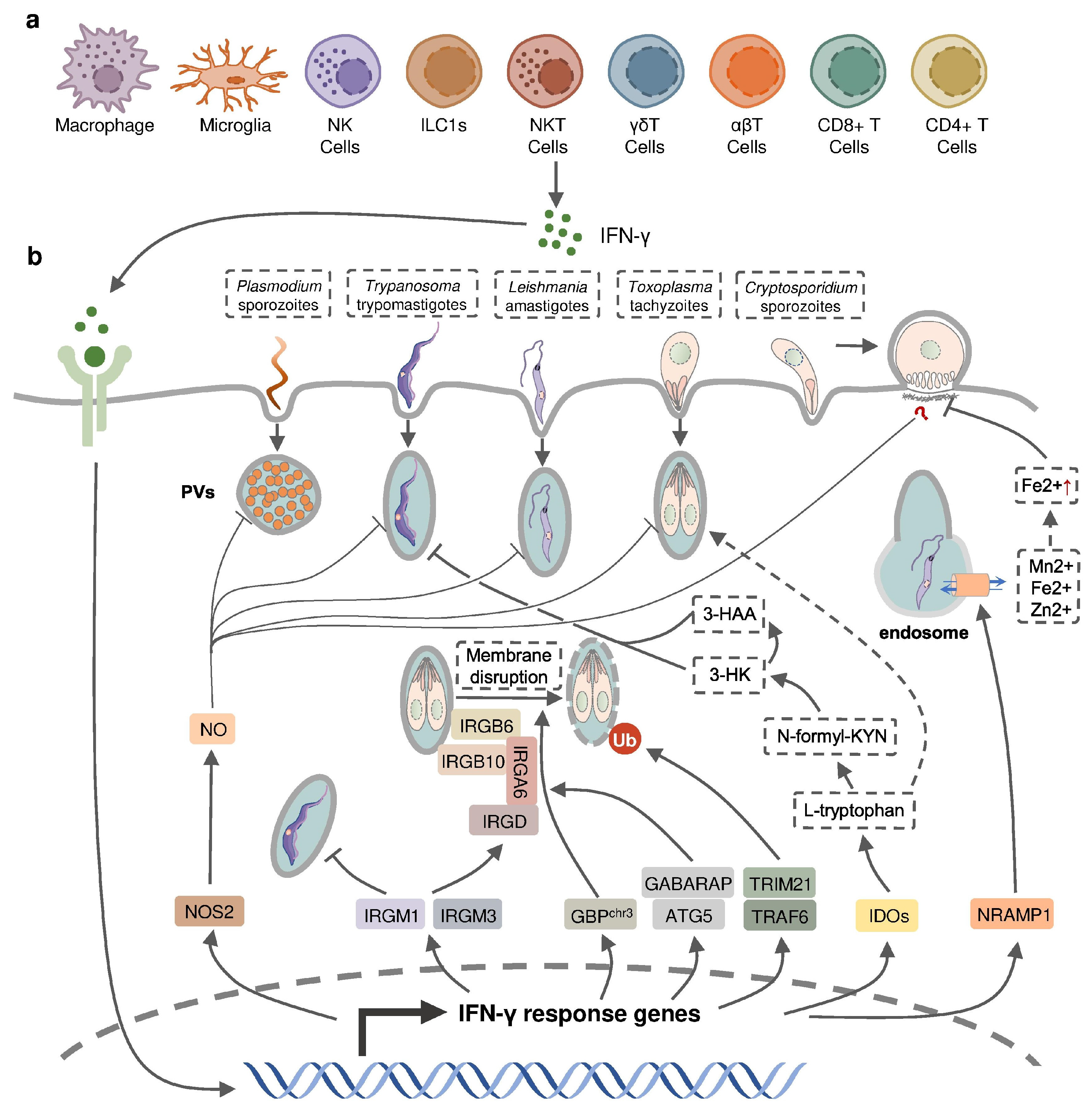
Figure 1. IFN-γ-inducible cell-autonomous defense against intracellular protozoan parasite infections. (a) IFN-γ can be produced by multiple cell types, including immune cells, during infection and (b) acts on infected host cells to eliminate intracellular parasite by nitric oxide (NO) production, the disruption of parasitophorous vacuoles (PVs) through IFN-inducible GTPase, the restriction of ion assimilation by NRAMP1, and the inhibition of nutrient acquisition by IDOs.
Table 1. Effects of IFN-γ in host following intracellular protozoan parasite infections.
| Protective Effects | |||
| Parasite Species | Treatments and Findings | Effects | Ref. |
| P. falciparum | Recombinant IFN-γ in human hepatocyte cell culture | Hepatic schizont development ↓ | [22] |
| IFN-γ production in infected children | Occurrence of high-density and clinical episode of infection ↓ | [14] | |
| High IFN-γ level in infected patients | Occurrence of cerebral malaria ↓ | [23] | |
| P. berghei | Recombinant IFN-γ in rats, mice and human hepatocyte cell culture | Hepatic schizont development ↓ | [22] |
| Monoclonal IFN-γ neutralizing antibody in mice | Parasitemia ↑ | [24] | |
| Early IFN-γ production in infected mice | Occurrence of cerebral malaria ↓ | [25] | |
| P. vivax | Recombinant IFN-γ in chimpanzees | Parasitemia ↓ | [22] |
| P. Chabaudi | Recombinant IFN-γ in mice. | Parasitemia ↓ Intraerythrocytic parasites ↓ |
[22] |
| IFN-γ−/− mice | Parasitemia ↑ | [26] | |
| P. cynomolgi | Recombinant IFN-γ in rhesus monkey | Hepatic schizont development ↓ | [22] |
| P. yoelii | Recombinant IFN-γ in mice | Parasitemia ↓ | [27] |
| IFN-γR−/− mice | Infection burden ↑ | [28] | |
| IFN-γ−/− mice | Parasitemia ↑ | [26] | |
| T. b. brucei | IFN-γ−/− mice | Parasitemia ↑ Survival time ↓ |
[10] |
| T. b. rhodesiense | IFN-γ−/− mice | Parasitemia ↑ Survival time ↓ |
[29] |
| L. major | IFN-γR−/− mice | Larger and progressing lesions | [30] |
| L. amazonesis | IFN-γ−/− mice | Devastating lesions in late infection stages | [31] |
| T. gondii | IFN-γ−/− mice | Survival ↓ Infection burden ↑ |
[32] |
| C. parvum | Recombinant IFN-γ in intestinal enterocytes cell culture | Infection burden ↓ | [33] |
| IFN-γ−/− mice | Survival ↓ Occurrence of chronic infection ↑ |
[34] | |
| Pathogenic Effects | |||
| Parasite Species | Treatments and Findings | Effects | Ref. |
| P. berghei | Late IFN-γ production in infected mice | Occurrence of cerebral malaria ↑ | [25] |
| Large amount of IFN-γ produced in infected mice | Susceptibility to cerebral malaria ↑ | [35][36] | |
| IFN-γR−/− mice | No cerebral malaria development | [37][38] | |
| IFN-γ−/− mice | No cerebral malaria development | [39][40] | |
| Monoclonal IFN-γ neutralizing antibody in mice | Occurrence of cerebral malaria ↓ Survival time ↑ |
[41] | |
| P. yoelii, P. chabaudi, P. berghei |
Overproduction of IFN-γ | Development of Tfh and GC B cell response ↓ Humoral immunity ↓ Autoimmune anemia ↑ |
[42][43] [44][45] [46] |
| T. congolense | Overproducing IFN-γ in mice | Survival time ↓ | [47] |
| Reducing production of IFN-γ in mice | Survival time ↑ | [48] | |
| Monoclonal IFN-γ neutralizing antibody in mice | Susceptibility ↓ | [49] | |
2. IFN-γ in Host Defense against Protozoan Parasites
IFN-γ appears to be critical in controlling the infections of many intracellular parasites (Table 1). Exogenous IFN-γ was found to significantly diminish infections of Plasmodium in mice, rats, non-human primates, as well as in in vitro human hepatocytes, by inhibiting the parasite DNA replication during liver-stage development [22]. However, Plasmodium has also evolved a strategy to evade the host defense during liver-stage development by suppressing the expression of pro-inflammatory cytokines including IFN-γ in hepatic mononuclear cells [50]. Plasmodium-infected mice administrated with recombinant IFN-γ exhibited a suppressed blood-stage infection with the delayed onset of parasitemia, decreased levels of infected erythrocyte, and increased survival [27][51][52]. The positive effect of endogenous IFN-γ in the host defense against Plasmodium was determined from the increased susceptibility of rats treated with an anti-IFN-γ neutralizing antibody and in mice deficient in IFN-γ or the IFN-γ receptor [24][26][28]. Human studies also indicated a positive correlation between low IFN-γ production by live Plasmodium-stimulated peripheral blood mononuclear cells and the increased occurrence of symptomatic malaria as well as the risk of moderate-to-high-density P. falciparum reinfection [14]. Early IFN-γ production was shown to contribute to the protection against the development of murine cerebral malaria, the most severe neurological complication of Plasmodium infection, in P. berghei-infected mice and peripheral levels of IFN-γ were found to drop just before the onset of both human and murine cerebral malaria [23][25]. IFN-γ can be induced by malaria vaccines, as higher numbers of IFN-γ-producing T cells and increased IFN-γ level were detected in vaccine-treated subjects in several clinical trials [53][54][55]. Vaccination with chemically attenuated, asexual, blood-stage Plasmodium falciparum induces anti-parasitic cellular immune responses involving IFN-γ in Plasmodium-naïve volunteers [56]. Nevertheless, a subunit vaccine targeting Plasmodium falciparum circumsporozoite protein (CSP) activates the host immune responses dominated by parasite specific IgG antibody instead of IFN-γ [57]. Mice with a deficiency of IFN-γ or IFN-γ receptor have a higher susceptibility to L. major infection, accompanied by elevated Th2-type responses compared to the wild-type mice, but IFN-γ-deficient mice do not appear to succumb to L. amazonensis until 2 months post infection, suggesting that IFN-γ is induced at different stages of infection by diverse Leishmania species [30][31][58]. Similarly, IFN-γ or IFN-γ-receptor-deficient mice exhibited high susceptibility to infections by T. b. rhodesiense, T. b. brucei, and T. gondii [10][29][32][59]. While exogenous IFN-γ inhibits the development of C. parvum in cultured enterocytes without the need of immune effector cells, both IFN-γ-deficient and anti-IFN-γ-antibody-treated neonatal mice became more susceptible to C. parvum infection [33][34].
IFN-inducible cell-autonomous defense, including parasiticidal activity mediated by nitic oxide (NO), the disruption of parasitophorous vacuoles (PVs) related to IFN-inducible GTPase, the restriction of ion assimilation by natural resistance-associated macrophage protein 1 (NRAMP1), and the inhibition of nutrient acquisition by indoleamine 2,3-dioxygenases (IDOs), is critical for the restriction of parasite growth in infected cells and the elimination of the parasite inside the parasite-containing subcellular compartments [60] (Figure 1). Previous studies have underlined the role of nitric oxide synthase 2 (NOS2, iNOS)-NO in IFN-γ-mediated parasiticidal activity against intracellular protozoan parasites. IFN-γ-induced NO production showed the most evident parasiticidal activity against T. cruzi trypomastigotes and L. major amastigotes in IFN-γ-activated macrophages and P. falciparum as well as P. yoelii sporozoites in human and mouse hepatocytes, respectively [28][61][62][63]. Correspondingly, Nos2-deficient mice were highly susceptible to these pathogens [58][61][62]. NO production was reported to be induced by IFN-γ in hosts infected with T. gondii. Nevertheless, IFN-inducible NO might play a limited role and function at later time points in hosts infected with tachyzoites of the less virulent type II T. gondii, whereas it was essential in parasite control in virulent type I-T. gondii-strain-infected hosts [64]. C. parvum-infected mice had an increased level of NOS2 which was partially attributed to activated IFN-γ signaling [34]. A slightly longer infection period was observed in C. parvum-infected neonatal Nos2-deficient mice [65]. Nonetheless, several other studies with human enterocytes and mouse models indicated that the protective action of IFN-γ against C. parvum infection is independent of NO activity [33][66][67]. Thus, the precise mechanisms of NO-mediated antiprotozoal activity are still incompletely understood.
IFN-inducible immunity-related GTPases (IRGs) are a subfamily of IFN-γ-inducible GTPases characterized by a particular molecular weight between 21 to 47 kDa. They defend host cells against protozoa by targeting the PVs. IRGs can be divided into two groups—GKS-containing IRGs (IRGA, IRGB, IRGC or IRGD groups) and GMS-containing IRGs (IRGMs)—based on their canonical-lysine-containing (lysine, K) and non-canonical (methionine, M) G1 motifs (GxxG[K/M]S/T) within the conserved amino-terminal catalytic GTPase domain [68]. IRGM1, IRGM3 and IRGA6 enhance the IFN-γ-induced control of avirulent T. gondii strain in macrophages and astrocytes, which might account for the limited effect of NO in hosts infected with this parasite [69][70][71][72][73][74]. In host cells infected with avirulent T. gondii, IRGM proteins that act as guanine nucleotide dissociation inhibitors under an uninfected status are released from the endoplasmic reticulum which “turns on” the GKS-containing IRGs to target the PV [68]. IRGM1 also contributes to the elimination of T. cruzi infection in macrophages as IRGM1 KO macrophages displayed a defective intracellular killing of T. cruzi amastigotes [75]. A hierarchical model revealed the intrinsic order of IRGs when they are recruited to PV which indicated that IRGB6 and possibly IRGB10 bind to the vacuole before IRGA6, while IRGD is loaded last [76]. The recruitment of these molecules leads to vesiculation, membrane disruption, and sometimes necroptosis of the targeted PVs.
The other subfamily of IFN-γ-inducible GTPases, guanylate-binding proteins (GBPs), consists of a set of proteins with molecular weights between 65–73 kDa, comprising 7 and 11 members in humans and mice, respectively [77]. GBP genes are categorized into two clusters located on chromosome 3 and chromosome 5 in mice. Mice lacking GBPs on chromosome 3 (GBPchr3), including GBP1, GBP2, GBP3, GBP5 and GBP7, were highly susceptible to T. gondii infection due to the insufficient disruption of the PVs [78]. Moreover, macrophages lacking GBPchr3 showed defective loading of IRGB6 on the T. gondii PV membrane (PVM), suggesting that GBPs and IRGs coordinate with each other in PV targeting [78]. Compared with GBPchr3-deficient mice, mice deficient in GBP1 or GBP2 exhibited a milder decline of survival rates following T. gondii infection, indicating the complementary roles of each GBP on chromosome 3 in the host defense against parasite infection [79][80]. On the contrary, GBP1 was not recruited to T. cruzi compartments suggesting a protozoan specificity of GBP-mediated PV disruption [81]. The structural and biochemical cues of IFN-γ-inducible GTPase for targeting these molecules to the PV and whether membrane disruption is due to a direct effect of IRG activity, or a result of some intermediary molecules, remains unclear. Recent evidence suggests the C-terminal isoprenylation of GBP2 regulates the recruitment of GBP2 to the PVM by recognizing the ubiquitination on the PVM, which differentiates between the host membrane and the PVM [82]. E3 ligases such as TRAF6 and TRIM21 modulate ubiquitination of T. gondii PVM following IFN-γ treatment, whereas the dependent effect of these molecules on the IFN-γ-mediated elimination of T. gondii is controversial [83][84][85]. Interestingly, the distribution of GBPs in the host cytoplasm triggering the disruption of PVM is also regulated by ubiquitin-like autophagy proteins, such as autophagy-regulated gene 5 (ATG5) and GABARAP autophagy proteins, in an autophagy-independent fashion [86][87].
NRAMP1 is a highly hydrophobic integral membrane phosphoglycoprotein (~100 kD), expressed primarily in the late endosomal/early endosomal compartment of macrophages and polymorphonuclear leukocytes as a membrane transporter [88]. It has been shown to transport divalent ion cations, such as Mn2+, Fe2+ and Zn2+, to prevent the intracellular pathogens from these divalent metals essential for parasite survival [88][89]. NRAMP1 has been identified as an essential factor in the host defense against L. donovani, but the intrinsic mechanism remains unclear [90]. A previous study has revealed the correlation between cellular Fe2+ concentration and the IFN-γ-induced inhibition of C. parvum infection in intestinal enterocytes, but whether NRAMP1 is involved has not been investigated [33].
IDOs, IDO1 and IDO2, are both IFN-inducible, haem-containing oxidoreductases that degrade L-tryptophan to generate N-formylkynurenine (N-formyl-KYN) in the kynurenine pathway [91]. The removal of L-tryptophan restricted the growth of T. gondii in several IFN-γ-stimulated human cell lines including fibroblasts, lung epithelial cells, glioblastoma cells, retinal pigment epithelial cells, and macrophages [92][93][94][95][96][97][98]. Moreover, increased susceptibility to T. gondii occurred in mice with a double deficiency of IDO1 and IDO2 but not in IDO1-deficient mice, suggesting a significant role of both IDOs in the restriction of T. gondii infection in vivo [99]. IDOs may also control T. cruzi infection through the downstream kynurenine catabolites 3-hydroxykynurenine (3-HK) and 3-hydroxyanthranilic acid (3-HAA), which are likely to be harmful to T. cruzi amastigotes and trypomastigotes [100].
Additionally, IFN-γ could increase the expression of endothelial vascular cell adhesion molecule 1 (VCAM-1) to facilitate the recruitment of CD8+ T cells in the brain of mice chronically infected with T. gondii and enhance the cytotoxic potential of CD8+ T cells by inducing NO, which contributes to the host defense against parasites in the brain [17][101][102]. IFN-γ has been reported to modulate B-cell-mediated humoral immunity in Plasmodium infection via modulating the class-switching of antibody-producing B cells as IFN-γ-deficient mice produce less parasite-specific IgM, IgG3 and cytophilic IgG2a than wild-type mice [103].
3. IFN-γ in the Pathogenesis of Protozoan Infection
In contrast to the protective effect of IFN-γ, the response has also been reported to be involved in the pathogenesis of protozoan infection (Table 1). Although IFN-γ production as early as 24 h p.i. prevented the occurrence of experimental cerebral malaria in Plasmodium-infected mice, mice with late IFN-γ production at 3 to 4 days p.i. were found to develop severe experimental cerebral malaria [25]. IFN-γ mRNA accumulation was detected in mice susceptible to cerebral malaria [35]. The suppressed development of cerebral malaria was observed in Plasmodium-infected mice following the administration of anti-IFN-γ monoclonal antibody, treatment with IFN-γ-suppressive IL-10, inhibition of IFN-γ production, or deficiency of the IFN-γ receptor [36][37][39][41]. IFN-γ may mediate the development of experimental cerebral malaria through various mechanisms. IFN-γ, together with tumor necrosis factor (TNF) and lymphotoxin α, enhance the activation and apoptosis of the brain endothelium through the activation of endothelial cell and subsequently increased local binding of platelets [41][104][105]. IFN-γ is also necessary for the recruitment of CD8+ T cells in the brain by inducing the expression of canonical adhesion molecules, such as ICAM-1, CXCL9, and CXCL10. Accumulated CD8+ T cells mediate the immune responses against infected red blood cells sequestered in the brain and the lungs in susceptible mice, facilitating development of experimental CM [38][40]. The precise effect of IFN-γ in the development of cerebral malaria is still controversial. T. congolense highly susceptible BALB/c mice displayed significantly higher plasma IFN-γ levels compared to infected parasite-resistant C57BL/6 mice [49][106][107][108]. The IFN-γ-mediated accumulation and activation of erythrophagocytic myeloid cells led to acute anemia, liver injury, and a reduced survival time in T. brucei or T. congolense-infected BALB/c mice [47][48][109]. The overproduction of IFN-γ, induced by blocking IL-10R, shortened the survival time of both C57BL/6 and BALB/c mice following T. congolense infection [47]. T. congolense-susceptible BALB/c mice could switch to a relatively resistant-like phenotype by the neutralization of IFN-γ or by reducing the production of IFN-γ through the depletion of IL-12 during T. congolense infection [48][49]. Moreover, IFN-γ is also a crucial mediator in the humoral immunity that may exacerbate infection outcomes, leading to parasite-associated autoimmune disorders and chronic infection. Plasmodium DNA could induce autoreactive responses against erythrocytes by activating a population of B cells expressing CD11c and T-bet, in which IFN-γ acted as an essential factor, together with parasitic DNA to promote the expansion of autoreactive T-bet+ B cells, a major producer of autoantibodies promoting malarial anemia [46]. IFN-γ also contributed to the inhibition of T follicular helper cell differentiation during sever malaria infection, resulting in an impaired germ center B cell response and inefficient production of antibody-secreting plasma cells [42][43][44][45].
The precise role of IFN-γ in the host defense against protozoan parasite infection and in the pathogenesis of infection can be different during the infection of different pathogens, at different infectious stages, or in hosts with different intrinsic immune statues. Therefore, close attention to the alteration in IFN-γ levels and the IFN-γ-mediated immune response is necessary for timely adjustments of therapeutic strategies and predictions of prognosis of infection.
References
- Horowitz, A.; Riley, E.M. Activation of Human NK Cells by Plasmodium-Infected Red Blood Cells. Methods Mol. Biol. 2013, 923, 447–464.
- Barakat, F.M.; McDonald, V.; Di Santo, J.P.; Korbel, D.S. Roles for NK Cells and an NK Cell-Independent Source of Intestinal Gamma Interferon for Innate Immunity to Cryptosporidium Parvum Infection. Infect. Immun. 2009, 77, 5044–5049.
- Onyilagha, C.; Kuriakose, S.; Ikeogu, N.; Kung, S.K.P.; Uzonna, J.E. NK Cells Are Critical for Optimal Immunity to Experimental Trypanosoma Congolense Infection. J. Immunol. 2019, 203, 964–971.
- Artavanis-Tsakonas, K.; Riley, E.M. Innate Immune Response to Malaria: Rapid Induction of IFN-γ from Human NK Cells by Live Plasmodium Falciparum-Infected Erythrocytes. J. Immunol. 2002, 169, 2956–2963.
- Becker, I.; Salaiza, N.; Aguirre, M.; Delgado, J.; Carrillo-Carrasco, N.; Kobeh, L.G.; Ruiz, A.; Cervantes, R.; Torres, A.P.; Cabrera, N.; et al. Leishmania Lipophosphoglycan (LPG) Activates NK Cells through Toll-like Receptor-2. Mol. Biochem. Parasitol. 2003, 130, 65–74.
- Filipe-Santos, O.; Pescher, P.; Breart, B.; Lippuner, C.; Aebischer, T.; Glaichenhaus, N.; Späth, G.F.; Bousso, P. A Dynamic Map of Antigen Recognition by CD4 T Cells at the Site of Leishmania Major Infection. Cell Host Microbe 2009, 6, 23–33.
- Walther, M.; Jeffries, D.; Finney, O.C.; Njie, M.; Ebonyi, A.; Deininger, S.; Lawrence, E.; Ngwa-Amambua, A.; Jayasooriya, S.; Cheeseman, I.H.; et al. Distinct Roles for FOXP3+ and FOXP3− CD4+ T Cells in Regulating Cellular Immunity to Uncomplicated and Severe Plasmodium Falciparum Malaria. PLoS Pathog. 2009, 5, e1000364.
- Ong’echa, J.M.O.; Lal, A.A.; Terlouw, D.J.; Ter Kuile, F.O.; Kariuki, S.K.; Udhayakumar, V.; Orago, A.S.S.; Hightower, A.W.; Nahlen, B.L.; Shi, Y.P. Association of Interferon-Gamma Responses to Pre-Erythrocytic Stage Vaccine Candidate Antigens of Plasmodium Falciparum in Young Kenyan Children with Improved Hemoglobin Levels: XV. Asembo Bay Cohort Project. Am. J. Trop. Med. Hyg. 2003, 68, 590–597.
- Wang, X.; Kang, H.; Kikuchi, T.; Suzuki, Y. Gamma Interferon Production, but Not Perforin-Mediated Cytolytic Activity, of T Cells Is Required for Prevention of Toxoplasmic Encephalitis in BALB/c Mice Genetically Resistant to the Disease. Infect. Immun. 2004, 72, 4432–4438.
- Namangala, B.; Noël, W.; De Baetselier, P.; Brys, L.; Beschin, A. Relative Contribution of Interferon-γ and Interleukin-10 to Resistance to Murine African Trypanosomosis. J. Infect. Dis. 2001, 183, 1794–1800.
- Miller, J.L.; Sack, B.K.; Baldwin, M.; Vaughan, A.M.; Kappe, S.H.I. Interferon-Mediated Innate Immune Responses against Malaria Parasite Liver Stages. Cell Rep. 2014, 7, 436–447.
- Soulard, V.; Roland, J.; Sellier, C.; Gruner, A.C.; Leite-de-Moraes, M.; Franetich, J.-F.; Rénia, L.; Cazenave, P.-A.; Pied, S. Primary Infection of C57BL/6 Mice with Plasmodium Yoelii Induces a Heterogeneous Response of NKT Cells. Infect. Immun. 2007, 75, 2511–2522.
- Hviid, L.; Kurtzhals, J.A.L.; Adabayeri, V.; Loizon, S.; Kemp, K.; Goka, B.Q.; Lim, A.; Mercereau-Puijalon, O.; Akanmori, B.D.; Behr, C. Perturbation and Proinflammatory Type Activation of Vδ1+ Γδ T Cells in African Children with Plasmodium Falciparum Malaria. Infect. Immun. 2001, 69, 3190–3196.
- D’Ombrain, M.C.; Robinson, L.J.; Stanisic, D.I.; Taraika, J.; Bernard, N.; Michon, P.; Mueller, I.; Schofield, L. Association of Early Interferon-γ Production with Immunity to Clinical Malaria: A Longitudinal Study among Papua New Guinean Children. Clin. Infect. Dis. 2008, 47, 1380–1387.
- Pamplona, A.; Silva-Santos, B. Γδ T Cells in Malaria: A Double-edged Sword. FEBS J. 2021, 288, 1118–1129.
- Suzuki, Y.; Claflin, J.; Wang, X.; Lengi, A.; Kikuchi, T. Microglia and Macrophages as Innate Producers of Interferon-Gamma in the Brain Following Infection with Toxoplasma Gondii. Int. J. Parasitol. 2005, 35, 83–90.
- Wang, X.; Suzuki, Y. Microglia Produce IFN-γ Independently from T Cells During Acute Toxoplasmosis in the Brain. J. Interferon Cytokine Res. 2007, 27, 599–605.
- Klose, C.S.N.; Flach, M.; Möhle, L.; Rogell, L.; Hoyler, T.; Ebert, K.; Fabiunke, C.; Pfeifer, D.; Sexl, V.; Fonseca-Pereira, D.; et al. Differentiation of Type 1 ILCs from a Common Progenitor to All Helper-like Innate Lymphoid Cell Lineages. Cell 2014, 157, 340–356.
- Torre, D.; Speranza, F.; Giola, M.; Matteelli, A.; Tambini, R.; Biondi, G. Role of Th1 and Th2 Cytokines in Immune Response to Uncomplicated Plasmodium Falciparum Malaria. Clin. Vaccine Immunol. 2002, 9, 348–351.
- Loevenich, K.; Ueffing, K.; Abel, S.; Hose, M.; Matuschewski, K.; Westendorf, A.M.; Buer, J.; Hansen, W. DC-Derived IL-10 Modulates Pro-Inflammatory Cytokine Production and Promotes Induction of CD4+IL-10+ Regulatory T Cells during Plasmodium Yoelii Infection. Front. Immunol. 2017, 8, 152.
- Ivanova, D.L.; Denton, S.L.; Fettel, K.D.; Sondgeroth, K.S.; Munoz Gutierrez, J.; Bangoura, B.; Dunay, I.R.; Gigley, J.P. Innate Lymphoid Cells in Protection, Pathology, and Adaptive Immunity During Apicomplexan Infection. Front. Immunol. 2019, 10, 196.
- Doolan, D.L.; Hoffman, S.L. The Complexity of Protective Immunity against Liver-Stage Malaria. J. Immunol. 2000, 165, 1453–1462.
- Prakash, D.; Fesel, C.; Jain, R.; Cazenave, P.-A.; Mishra, G.C.; Pied, S. Clusters of Cytokines Determine Malaria Severity in Plasmodium Falciparum–Infected Patients from Endemic Areas of Central India. J. Infect. Dis. 2006, 194, 198–207.
- Schofield, L.; Villaquiran, J.; Ferreira, A.; Schellekens, H.; Nussenzweig, R.; Nussenzweig, V. γ Interferon, CD8+ T Cells and Antibodies Required for Immunity to Malaria Sporozoites. Nature 1987, 330, 664–666.
- Mitchell, A.J.; Hansen, A.M.; Hee, L.; Ball, H.J.; Potter, S.M.; Walker, J.C.; Hunt, N.H. Early Cytokine Production Is Associated with Protection from Murine Cerebral Malaria. Infect. Immun. 2005, 73, 5645–5653.
- van der Heyde, H.C.; Pepper, B.; Batchelder, J.; Cigel, F.; Weidanz, W.P. The Time Course of Selected Malarial Infections in Cytokine-Deficient Mice. Exp. Parasitol. 1997, 85, 206–213.
- Shear, H.L.; Srinivasan, R.; Nolan, T.; Ng, C. Role of IFN-Gamma in Lethal and Nonlethal Malaria in Susceptible and Resistant Murine Hosts. J. Immunol. 1989, 143, 2038–2044.
- Tsuji, M.; Miyahira, Y.; Nussenzweig, R.S.; Aguet, M.; Reichel, M.; Zavala, F. Development of Antimalaria Immunity in Mice Lacking IFN-Gamma Receptor. J. Immunol. 1995, 154, 5338–5344.
- Hertz, C.J.; Filutowicz, H.; Mansfield, J.M. Resistance to the African Trypanosomes Is IFN-γ Dependent. J. Immunol. 1998, 161, 6775–6783.
- Swihart, K.; Fruth, U.; Messmer, N.; Hug, K.; Behin, R.; Huang, S.; Del Giudice, G.; Aguet, M.; Louis, J.A. Mice from a Genetically Resistant Background Lacking the Interferon γ Receptor Are Susceptible to Infection with Leishmania Major but Mount a Polarized T Helper Cell 1-Type CD4+ T Cell Response. J. Exp. Med. 1995, 181, 961–971.
- Pinheiro, R.O.; Rossi-Bergmann, B. Interferon-Gamma Is Required for the Late but Not Early Control of Leishmania Amazonensis Infection in C57Bl/6 Mice. Mem. Inst. Oswaldo Cruz 2007, 102, 79–82.
- Scharton-Kersten, T.M.; Wynn, T.A.; Denkers, E.Y.; Bala, S.; Grunvald, E.; Hieny, S.; Gazzinelli, R.T.; Sher, A. In the Absence of Endogenous IFN-Gamma, Mice Develop Unimpaired IL-12 Responses to Toxoplasma Gondii While Failing to Control Acute Infection. J. Immunol. 1996, 157, 4045–4054.
- Pollok, R.C.G.; Farthing, M.J.G.; Bajaj-Elliott, M.; Sanderson, I.R.; McDonald, V. Interferon Gamma Induces Enterocyte Resistance against Infection by the Intracellular Pathogen Cryptosporidium Parvum. Gastroenterology 2001, 120, 99–107.
- Lacroix, S.; Mancassola, R.; Naciri, M.; Laurent, F. Cryptosporidium Parvum-Specific Mucosal Immune Response in C57BL/6 Neonatal and Gamma Interferon-Deficient Mice: Role of Tumor Necrosis Factor Alpha in Protection. Infect. Immun. 2001, 69, 1635–1642.
- de Kossodo, S.; Grau, G.E. Profiles of Cytokine Production in Relation with Susceptibility to Cerebral Malaria. J. Immunol. 1993, 151, 4811–4820.
- Kossodo, S.; Monso, C.; Juillard, P.; Velu, T.; Goldman, M.; Grau, G.E. Interleukin-10 Modulates Susceptibility in Experimental Cerebral Malaria. Immunology 1997, 91, 536–540.
- Amani, V.; Vigário, A.M.; Belnoue, E.; Marussig, M.; Fonseca, L.; Mazier, D.; Rénia, L. Involvement of IFN-Gamma Receptor-Medicated Signaling in Pathology and Anti-Malarial Immunity Induced by Plasmodium Berghei Infection. Eur. J. Immunol. 2000, 30, 1646–1655.
- Belnoue, E.; Potter, S.M.; Rosa, D.S.; Mauduit, M.; Grüner, A.C.; Kayibanda, M.; Mitchell, A.J.; Hunt, N.H.; Rénia, L. Control of Pathogenic CD8+ T Cell Migration to the Brain by IFN-γ during Experimental Cerebral Malaria. Parasite Immunol. 2008, 30, 544–553.
- Yañez, D.M.; Manning, D.D.; Cooley, A.J.; Weidanz, W.P.; Heyde, H.C. van der Participation of Lymphocyte Subpopulations in the Pathogenesis of Experimental Murine Cerebral Malaria. J. Immunol. 1996, 157, 1620–1624.
- Villegas-Mendez, A.; Greig, R.; Shaw, T.N.; de Souza, J.B.; Gwyer Findlay, E.; Stumhofer, J.S.; Hafalla, J.C.R.; Blount, D.G.; Hunter, C.A.; Riley, E.M.; et al. IFN-γ-Producing CD4+ T Cells Promote Experimental Cerebral Malaria by Modulating CD8+ T Cell Accumulation within the Brain. J. Immunol. 2012, 189, 968–979.
- Grau, G.E.; Heremans, H.; Piguet, P.F.; Pointaire, P.; Lambert, P.H.; Billiau, A.; Vassalli, P. Monoclonal Antibody against Interferon Gamma Can Prevent Experimental Cerebral Malaria and Its Associated Overproduction of Tumor Necrosis Factor. Proc. Natl. Acad. Sci. USA 1989, 86, 5572–5574.
- Zander, R.A.; Obeng-Adjei, N.; Guthmiller, J.J.; Kulu, D.I.; Li, J.; Ongoiba, A.; Traore, B.; Crompton, P.D.; Butler, N.S. PD-1 Co-Inhibitory and OX40 Co-Stimulatory Crosstalk Regulates Helper T Cell Differentiation and Anti-Plasmodium Humoral Immunity. Cell Host Microbe 2015, 17, 628–641.
- Ryg-Cornejo, V.; Ioannidis, L.J.; Ly, A.; Chiu, C.Y.; Tellier, J.; Hill, D.L.; Preston, S.P.; Pellegrini, M.; Yu, D.; Nutt, S.L.; et al. Severe Malaria Infections Impair Germinal Center Responses by Inhibiting T Follicular Helper Cell Differentiation. Cell Rep. 2016, 14, 68–81.
- Obeng-Adjei, N.; Portugal, S.; Tran, T.M.; Yazew, T.B.; Skinner, J.; Li, S.; Jain, A.; Felgner, P.L.; Doumbo, O.K.; Kayentao, K.; et al. Circulating Th1-Cell-Type Tfh Cells That Exhibit Impaired B Cell Help Are Preferentially Activated during Acute Malaria in Children. Cell Rep. 2015, 13, 425–439.
- Guthmiller, J.J.; Graham, A.C.; Zander, R.A.; Pope, R.L.; Butler, N.S. Cutting Edge: IL-10 Is Essential for the Generation of Germinal Center B Cell Responses and Anti-Plasmodium Humoral Immunity. J. Immunol. 2017, 198, 617–622.
- Rivera-Correa, J.; Guthmiller, J.J.; Vijay, R.; Fernandez-Arias, C.; Pardo-Ruge, M.A.; Gonzalez, S.; Butler, N.S.; Rodriguez, A. Plasmodium DNA-Mediated TLR9 Activation of T-Bet+ B Cells Contributes to Autoimmune Anaemia during Malaria. Nat. Commun. 2017, 8, 1282.
- Shi, M.; Pan, W.; Tabel, H. Experimental African Trypanosomiasis: IFN-γ Mediates Early Mortality. Eur. J. Immunol. 2003, 33, 108–118.
- Barkhuizen, M.; Magez, S.; Ryffel, B.; Brombacher, F. Interleukin-12p70 Deficiency Increases Survival and Diminishes Pathology in Trypanosoma Congolense Infection. J. Infect. Dis. 2008, 198, 1284–1291.
- Uzonna, J.E.; Kaushik, R.S.; Gordon, J.R.; Tabel, H. Experimental Murine Trypanosoma Congolense Infections. I. Administration of Anti-IFN-γ Antibodies Alters Trypanosome-Susceptible Mice to a Resistant-Like Phenotype. J. Immunol. 1998, 161, 5507–5515.
- Siddiqui, A.J.; Bhardwaj, J.; Goyal, M.; Prakash, K.; Adnan, M.; Alreshidi, M.M.; Patel, M.; Soni, A.; Redman, W. Immune Responses in Liver and Spleen against Plasmodium Yoelii Pre-Erythrocytic Stages in Swiss Mice Model. J. Adv. Res. 2020, 24, 29–41.
- Clark, I.A.; Hunt, N.H.; Butcher, G.A.; Cowden, W.B. Inhibition of Murine Malaria (Plasmodium Chabaudi) in Vivo by Recombinant Interferon-Gamma or Tumor Necrosis Factor, and Its Enhancement by Butylated Hydroxyanisole. J. Immunol. 1987, 139, 3493–3496.
- De Souza, J.B.; Williamson, K.H.; Otani, T.; Playfair, J.H. Early Gamma Interferon Responses in Lethal and Nonlethal Murine Blood-Stage Malaria. Infect. Immun. 1997, 65, 1593–1598.
- Kester, K.E.; Cummings, J.F.; Ofori-Anyinam, O.; Ockenhouse, C.F.; Krzych, U.; Moris, P.; Schwenk, R.; Nielsen, R.A.; Debebe, Z.; Pinelis, E.; et al. Randomized, Double-Blind, Phase 2a Trial of Falciparum Malaria Vaccines RTS,S/AS01B and RTS,S/AS02A in Malaria-Naive Adults: Safety, Efficacy, and Immunologic Associates of Protection. J. Infect. Dis. 2009, 200, 337–346.
- Chuang, I.; Sedegah, M.; Cicatelli, S.; Spring, M.; Polhemus, M.; Tamminga, C.; Patterson, N.; Guerrero, M.; Bennett, J.W.; McGrath, S.; et al. DNA Prime/Adenovirus Boost Malaria Vaccine Encoding P. Falciparum CSP and AMA1 Induces Sterile Protection Associated with Cell-Mediated Immunity. PLoS ONE 2013, 8, e55571.
- Seder, R.A.; Chang, L.-J.; Enama, M.E.; Zephir, K.L.; Sarwar, U.N.; Gordon, I.J.; Holman, L.A.; James, E.R.; Billingsley, P.F.; Gunasekera, A.; et al. Protection Against Malaria by Intravenous Immunization with a Nonreplicating Sporozoite Vaccine. Science 2013, 341, 1359–1365.
- Stanisic, D.I.; Fink, J.; Mayer, J.; Coghill, S.; Gore, L.; Liu, X.Q.; El-Deeb, I.; Rodriguez, I.B.; Powell, J.; Willemsen, N.M.; et al. Vaccination with Chemically Attenuated Plasmodium Falciparum Asexual Blood-Stage Parasites Induces Parasite-Specific Cellular Immune Responses in Malaria-Naïve Volunteers: A Pilot Study. BMC Med. 2018, 16, 184.
- Chawla, B.; Mahajan, B.; Oakley, M.; Majam, V.F.; Belmonte, A.; Sedegah, M.; Shimp, R.L.; Kaslow, D.C.; Kumar, S. Antibody-Dependent, Gamma Interferon-Independent Sterilizing Immunity Induced by a Subunit Malaria Vaccine. Infect. Immun. 2019, 87, e00236-19.
- Carneiro, M.B.H.; Lopes, M.E.D.M.; Vaz, L.G.; Sousa, L.M.A.; dos Santos, L.M.; de Souza, C.C.; Campos, A.C.D.A.; Gomes, D.A.; Gonçalves, R.; Tafuri, W.L.; et al. IFN-γ-Dependent Recruitment of CD4+ T Cells and Macrophages Contributes to Pathogenesis During Leishmania Amazonensis Infection. J. Interferon Cytokine Res. 2015, 35, 935–947.
- Wu, H.; Liu, G.; Shi, M. Interferon Gamma in African Trypanosome Infections: Friends or Foes? Front. Immunol. 2017, 8, 1105.
- MacMicking, J.D. Interferon-Inducible Effector Mechanisms in Cell-Autonomous Immunity. Nat. Rev. Immunol. 2012, 12, 367–382.
- Hölscher, C.; Köhler, G.; Müller, U.; Mossmann, H.; Schaub, G.A.; Brombacher, F. Defective Nitric Oxide Effector Functions Lead to Extreme Susceptibility of Trypanosoma Cruzi-Infected Mice Deficient in Gamma Interferon Receptor or Inducible Nitric Oxide Synthase. Infect. Immun. 1998, 66, 1208–1215.
- Mellouk, S.; Hoffman, S.L.; Liu, Z.Z.; de la Vega, P.; Billiar, T.R.; Nussler, A.K. Nitric Oxide-Mediated Antiplasmodial Activity in Human and Murine Hepatocytes Induced by Gamma Interferon and the Parasite Itself: Enhancement by Exogenous Tetrahydrobiopterin. Infect. Immun. 1994, 62, 4043–4046.
- Green, S.J.; Meltzer, M.S.; Hibbs, J.B.; Nacy, C.A. Activated Macrophages Destroy Intracellular Leishmania Major Amastigotes by an L-Arginine-Dependent Killing Mechanism. J. Immunol. 1990, 144, 278–283.
- Scharton-Kersten, T.M.; Yap, G.; Magram, J.; Sher, A. Inducible Nitric Oxide Is Essential for Host Control of Persistent but Not Acute Infection with the Intracellular Pathogen Toxoplasma Gondii. J. Exp. Med. 1997, 185, 1261–1274.
- Leitch, G.J.; He, Q. Reactive Nitrogen and Oxygen Species Ameliorate Experimental Cryptosporidiosis in the Neonatal BALB/c Mouse Model. Infect. Immun. 1999, 67, 5885–5891.
- Kolios, G.; Rooney, N.; Murphy, C.T.; Robertson, D.A.; Westwick, J. Expression of Inducible Nitric Oxide Synthase Activity in Human Colon Epithelial Cells: Modulation by T Lymphocyte Derived Cytokines. Gut 1998, 43, 56–63.
- Hayward, A.R.; Chmura, K.; Cosyns, M. Interferon-Gamma Is Required for Innate Immunity to Cryptosporidium Parvum in Mice. J. Infect. Dis. 2000, 182, 1001–1004.
- Hunn, J.P.; Koenen-Waisman, S.; Papic, N.; Schroeder, N.; Pawlowski, N.; Lange, R.; Kaiser, F.; Zerrahn, J.; Martens, S.; Howard, J.C. Regulatory Interactions between IRG Resistance GTPases in the Cellular Response to Toxoplasma Gondii. EMBO J. 2008, 27, 2495–2509.
- Zhao, Y.; Ferguson, D.J.P.; Wilson, D.C.; Howard, J.C.; Sibley, L.D.; Yap, G.S. Virulent Toxoplasma Gondii Evade Immunity-Related GTPase (IRG)-Mediated Parasite Vacuole Disruption Within Primed Macrophages. J. Immunol. 2009, 182, 3775–3781.
- Halonen, S.K.; Taylor, G.A.; Weiss, L.M. Gamma Interferon-Induced Inhibition of Toxoplasma Gondii in Astrocytes Is Mediated by IGTP. Infect. Immun. 2001, 69, 5573–5576.
- Martens, S.; Parvanova, I.; Zerrahn, J.; Griffiths, G.; Schell, G.; Reichmann, G.; Howard, J.C. Disruption of Toxoplasma Gondii Parasitophorous Vacuoles by the Mouse P47-Resistance GTPases. PLoS Pathog. 2005, 1, e24.
- Ling, Y.M.; Shaw, M.H.; Ayala, C.; Coppens, I.; Taylor, G.A.; Ferguson, D.J.P.; Yap, G.S. Vacuolar and Plasma Membrane Stripping and Autophagic Elimination of Toxoplasma Gondii in Primed Effector Macrophages. J. Exp. Med. 2006, 203, 2063–2071.
- Zhao, Y.; Wilson, D.; Matthews, S.; Yap, G.S. Rapid Elimination of Toxoplasma Gondii by Gamma Interferon-Primed Mouse Macrophages Is Independent of CD40 Signaling. Infect. Immun. 2007, 75, 4799–4803.
- Liesenfeld, O.; Parvanova, I.; Zerrahn, J.; Han, S.-J.; Heinrich, F.; Muñoz, M.; Kaiser, F.; Aebischer, T.; Buch, T.; Waisman, A.; et al. The IFN-γ-Inducible GTPase, Irga6, Protects Mice against Toxoplasma Gondii but Not against Plasmodium Berghei and Some Other Intracellular Pathogens. PLoS ONE 2011, 6, e20568.
- Santiago, H.C.; Feng, C.G.; Bafica, A.; Roffe, E.; Arantes, R.M.; Cheever, A.; Taylor, G.; Vierira, L.Q.; Aliberti, J.; Gazzinelli, R.T.; et al. Mice Deficient in LRG-47 Display Enhanced Susceptibility to Trypanosoma Cruzi Infection Associated with Defective Hemopoiesis and Intracellular Control of Parasite Growth. J. Immunol. 2005, 175, 8165–8172.
- Khaminets, A.; Hunn, J.P.; Könen-Waisman, S.; Zhao, Y.O.; Preukschat, D.; Coers, J.; Boyle, J.P.; Ong, Y.-C.; Boothroyd, J.C.; Reichmann, G.; et al. Coordinated Loading of IRG Resistance GTPases on to the Toxoplasma Gondii Parasitophorous Vacuole. Cell. Microbiol. 2010, 12, 939–961.
- Kim, B.-H.; Shenoy, A.R.; Kumar, P.; Bradfield, C.J.; MacMicking, J.D. IFN-Inducible GTPases in Host Defense. Cell Host Microbe 2012, 12, 432–444.
- Yamamoto, M.; Okuyama, M.; Ma, J.S.; Kimura, T.; Kamiyama, N.; Saiga, H.; Ohshima, J.; Sasai, M.; Kayama, H.; Okamoto, T.; et al. A Cluster of Interferon-γ-Inducible P65 GTPases Plays a Critical Role in Host Defense against Toxoplasma Gondii. Immunity 2012, 37, 302–313.
- Degrandi, D.; Kravets, E.; Konermann, C.; Beuter-Gunia, C.; Klümpers, V.; Lahme, S.; Rasch, E.; Mausberg, A.K.; Beer-Hammer, S.; Pfeffer, K. Murine Guanylate Binding Protein 2 (MGBP2) Controls Toxoplasma Gondii Replication. Proc. Natl. Acad. Sci. USA 2013, 110, 294–299.
- Selleck, E.M.; Fentress, S.J.; Beatty, W.L.; Degrandi, D.; Pfeffer, K.; Virgin, H.W.; MacMicking, J.D.; Sibley, L.D. Guanylate-Binding Protein 1 (Gbp1) Contributes to Cell-Autonomous Immunity against Toxoplasma Gondii. PLoS Pathog. 2013, 9, e1003320.
- Winter, S.V.; Niedelman, W.; Jensen, K.D.; Rosowski, E.E.; Julien, L.; Spooner, E.; Caradonna, K.; Burleigh, B.A.; Saeij, J.P.J.; Ploegh, H.L.; et al. Determinants of GBP Recruitment to Toxoplasma Gondii Vacuoles and the Parasitic Factors That Control It. PLoS ONE 2011, 6, e24434.
- Selleck, E.M.; Orchard, R.C.; Lassen, K.G.; Beatty, W.L.; Xavier, R.J.; Levine, B.; Virgin, H.W.; Sibley, L.D. A Noncanonical Autophagy Pathway Restricts Toxoplasma Gondii Growth in a Strain-Specific Manner in IFN-γ-Activated Human Cells. mBio 2015, 6, e01157-15.
- Lee, Y.; Sasai, M.; Ma, J.S.; Sakaguchi, N.; Ohshima, J.; Bando, H.; Saitoh, T.; Akira, S.; Yamamoto, M. P62 Plays a Specific Role in Interferon-γ-Induced Presentation of a Toxoplasma Vacuolar Antigen. Cell Rep. 2015, 13, 223–233.
- Haldar, A.K.; Foltz, C.; Finethy, R.; Piro, A.S.; Feeley, E.M.; Pilla-Moffett, D.M.; Komatsu, M.; Frickel, E.-M.; Coers, J. Ubiquitin Systems Mark Pathogen-Containing Vacuoles as Targets for Host Defense by Guanylate Binding Proteins. Proc. Natl. Acad. Sci. USA 2015, 112, E5628–E5637.
- Foltz, C.; Napolitano, A.; Khan, R.; Clough, B.; Hirst, E.M.; Frickel, E.-M. TRIM21 Is Critical for Survival of Toxoplasma Gondii Infection and Localises to GBP-Positive Parasite Vacuoles. Sci. Rep. 2017, 7, 5209.
- Choi, J.; Park, S.; Biering, S.B.; Selleck, E.; Liu, C.Y.; Zhang, X.; Fujita, N.; Saitoh, T.; Akira, S.; Yoshimori, T.; et al. The Parasitophorous Vacuole Membrane of Toxoplasma Gondii Is Targeted for Disruption by Ubiquitin-like Conjugation Systems of Autophagy. Immunity 2014, 40, 924–935.
- Sasai, M.; Sakaguchi, N.; Ma, J.S.; Nakamura, S.; Kawabata, T.; Bando, H.; Lee, Y.; Saitoh, T.; Akira, S.; Iwasaki, A.; et al. Essential Role for GABARAP Autophagy Proteins in Interferon-Inducible GTPase-Mediated Host Defense. Nat. Immunol. 2017, 18, 899–910.
- Jabado, N.; Jankowski, A.; Dougaparsad, S.; Picard, V.; Grinstein, S.; Gros, P. Natural Resistance to Intracellular Infections. J. Exp. Med. 2000, 192, 1237–1248.
- Kontoghiorghes, G.J.; Weinberg, E.D. Iron: Mammalian Defense Systems, Mechanisms of Disease, and Chelation Therapy Approaches. Blood Rev. 1995, 9, 33–45.
- Vidal, S.; Tremblay, M.L.; Govoni, G.; Gauthier, S.; Sebastiani, G.; Malo, D.; Skamene, E.; Olivier, M.; Jothy, S.; Gros, P. The Ity/Lsh/Bcg Locus: Natural Resistance to Infection with Intracellular Parasites Is Abrogated by Disruption of the Nramp1 Gene. J. Exp. Med. 1995, 182, 655–666.
- Savitz, J. The Kynurenine Pathway: A Finger in Every Pie. Mol. Psychiatry 2020, 25, 131–147.
- Pfefferkorn, E.R. Interferon Gamma Blocks the Growth of Toxoplasma Gondii in Human Fibroblasts by Inducing the Host Cells to Degrade Tryptophan. Proc. Natl. Acad. Sci. USA 1984, 81, 908–912.
- Däubener, W.; Spors, B.; Hucke, C.; Adam, R.; Stins, M.; Kim, K.S.; Schroten, H. Restriction of Toxoplasma Gondii Growth in Human Brain Microvascular Endothelial Cells by Activation of Indoleamine 2,3-Dioxygenase. Infect. Immun. 2001, 69, 6527–6531.
- Nagineni, C.N.; Pardhasaradhi, K.; Martins, M.C.; Detrick, B.; Hooks, J.J. Mechanisms of Interferon-Induced Inhibition of Toxoplasma Gondii Replication in Human Retinal Pigment Epithelial Cells. Infect. Immun. 1996, 64, 4188–4196.
- Murray, H.W.; Szuro-Sudol, A.; Wellner, D.; Oca, M.J.; Granger, A.M.; Libby, D.M.; Rothermel, C.D.; Rubin, B.Y. Role of Tryptophan Degradation in Respiratory Burst-Independent Antimicrobial Activity of Gamma Interferon-Stimulated Human Macrophages. Infect. Immun. 1989, 57, 845–849.
- Däubener, W.; Remscheid, C.; Nockemann, S.; Pilz, K.; Seghrouchni, S.; Mackenzie, C.; Hadding, U. Anti-Parasitic Effector Mechanisms in Human Brain Tumor Cells: Role of Interferon-Gamma and Tumor Necrosis Factor-Alpha. Eur. J. Immunol. 1996, 26, 487–492.
- Gupta, S.L.; Carlin, J.M.; Pyati, P.; Dai, W.; Pfefferkorn, E.R.; Murphy, M.J. Antiparasitic and Antiproliferative Effects of Indoleamine 2,3-Dioxygenase Enzyme Expression in Human Fibroblasts. Infect. Immun. 1994, 62, 2277–2284.
- Heseler, K.; Spekker, K.; Schmidt, S.K.; MacKenzie, C.R.; Däubener, W. Antimicrobial and Immunoregulatory Effects Mediated by Human Lung Cells: Role of IFN-Gamma-Induced Tryptophan Degradation. FEMS Immunol. Med. Microbiol. 2008, 52, 273–281.
- Divanovic, S.; Sawtell, N.M.; Trompette, A.; Warning, J.I.; Dias, A.; Cooper, A.M.; Yap, G.S.; Arditi, M.; Shimada, K.; DuHadaway, J.B.; et al. Opposing Biological Functions of Tryptophan Catabolizing Enzymes During Intracellular Infection. J. Infect. Dis. 2012, 205, 152–161.
- Knubel, C.P.; Martínez, F.F.; Fretes, R.E.; Lujan, C.D.; Theumer, M.G.; Cervi, L.; Motrán, C.C. Indoleamine 2,3-dioxigenase (IDO) Is Critical for Host Resistance against Trypanosoma Cruzi. FASEB J. 2010, 24, 2689–2701.
- Doolan, D.L.; Sedegah, M.; Hedstrom, R.C.; Hobart, P.; Yupin, C. Hoffman Circumventing Genetic Restriction of Protection against Malaria with Multigene DNA Immunization: CD8+ Cell-, Interferon Gamma-, and Nitric Oxide-Dependent Immunity. J. Exp. Med. 1996, 183, 1739–1746.
- Suzuki, Y.; Sa, Q.; Gehman, M.; Ochiai, E. Interferon-Gamma- and Perforin-Mediated Immune Responses for Resistance against Toxoplasma Gondii in the Brain. Expert Rev. Mol. Med. 2011, 13, e31.
- Su, Z.; Stevenson, M.M. Central Role of Endogenous Gamma Interferon in Protective Immunity against Blood-Stage Plasmodium Chabaudi AS Infection. Infect. Immun. 2000, 68, 4399–4406.
- Wassmer, S.C.; Combes, V.; Candal, F.J.; Juhan-Vague, I.; Grau, G.E. Platelets Potentiate Brain Endothelial Alterations Induced by Plasmodium Falciparum. Infect. Immun. 2006, 74, 645–653.
- Wassmer, S.C.; Lépolard, C.; Traoré, B.; Pouvelle, B.; Gysin, J.; Grau, G.E. Platelets Reorient Plasmodium Falciparum-Infected Erythrocyte Cytoadhesion to Activated Endothelial Cells. J. Infect. Dis. 2004, 189, 180–189.
- Uzonna, J.E.; Kaushik, R.S.; Gordon, J.R.; Tabel, H. Cytokines and Antibody Responses during Trypanosoma Congolense Infections in Two Inbred Mouse Strains That Differ in Resistance. Parasite Immunol. 1999, 21, 57–71.
- Tabel, H.; Kaushik, R.S.; Uzonna, J.E. Susceptibility and Resistance to Trypanosoma Congolense Infections. Microbes Infect. 2000, 2, 1619–1629.
- Tabel, H.; Wei, G.; Shi, M. T Cells and Immunopathogenesis of Experimental African Trypanosomiasis. Immunol. Rev. 2008, 225, 128–139.
- Stijlemans, B.; Beschin, A.; Magez, S.; Van Ginderachter, J.A.; De Baetselier, P. Iron Homeostasis and Trypanosoma Brucei Associated Immunopathogenicity Development: A Battle/Quest for Iron. Biomed Res. Int. 2015, 2015, 819389.
More
Information
Subjects:
Pathology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.4K
Revisions:
2 times
(View History)
Update Date:
24 Feb 2023
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No