Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Bryan Mathis | -- | 1794 | 2022-12-12 23:33:08 | | | |
| 2 | Dean Liu | Meta information modification | 1794 | 2022-12-15 08:30:30 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Mathis, B.J.; Kato, H.; Hiramatsu, Y. Nrf2 in the Heart versus Other Systems. Encyclopedia. Available online: https://encyclopedia.pub/entry/38619 (accessed on 24 July 2026).
Mathis BJ, Kato H, Hiramatsu Y. Nrf2 in the Heart versus Other Systems. Encyclopedia. Available at: https://encyclopedia.pub/entry/38619. Accessed July 24, 2026.
Mathis, Bryan J., Hideyuki Kato, Yuji Hiramatsu. "Nrf2 in the Heart versus Other Systems" Encyclopedia, https://encyclopedia.pub/entry/38619 (accessed July 24, 2026).
Mathis, B.J., Kato, H., & Hiramatsu, Y. (2022, December 12). Nrf2 in the Heart versus Other Systems. In Encyclopedia. https://encyclopedia.pub/entry/38619
Mathis, Bryan J., et al. "Nrf2 in the Heart versus Other Systems." Encyclopedia. Web. 12 December, 2022.
Copy Citation
Nuclear factor erythroid 2-related factor 2 (Nrf2) is a master regulator of the endogenous antioxidant response to reactive oxygen species as well as a controller of Phase II detoxification in response to xenobiotics. However, more is not necessarily better as the heart's unique biochemical and molecular composition make it susceptible to damage if the Nrf2 response is uncontrolled.
Nrf2
Keap1
cardiomyopathy
bardoloxone
1. Introduction
Reactive oxygen species (ROS) are both a normal byproduct of mitochondrial metabolism and an endproduct of oxidative biochemical reactions in the cell. Balanced levels of subcellular compartmental ROS are important for normal cellular functions, whereas dysregulated ROS, usually caused by relative insufficiency or impairment of the endogenous antioxidant defense system, attack cellular components leading to cellular damage and death, a state referred as to oxidative stress. To maintain cellular redox homeostasis and neutralize uncontrolled ROS, conserved antioxidant defense enzymes are placed under the control of the nuclear factor erythroid 2-related factor 2 (Nrf2) gene which is constitutively expressed in all higher-order animals. This gene, as a master antioxidant transcription factor, is responsible for global antioxidant activity in response to internally and externally sourced ROS threats but also modulates such species to maintain important intracellular second messenger capability. As the inhibitor of Nrf2, Kelch-like erythroid cell-derived protein with CNC homology-associated protein 1 (Keap1), is amenable to attack by exogenous compounds, research has focused its efforts to increase Nrf2 activity via direct interaction with Keap1. These compounds, such as oleanic acid derivative 2-cyano-3,12-dioxoolean-1,9- dien-28-oic acid (CDDO, bardoloxone), have been intensely studied in animal models and human trials as potential defense agents oxidative stress-associated diseases, such as cancer, chronic kidney disease, fatty liver, and endometriosis. Unfortunately, human trials employing CDDO and similar compounds for amelioration of these maladies have met with ambiguous and often disappointing results. Furthermore, multiple trials throughout 2007–2014 were terminated as unforeseen myocardial pathologies resulted. What follows is a survey of Nrf2, its endogenous regulation, action, and potential for exogenous modulation. Additionally, current clinical trial results are presented and analyzed for characteristics of Nrf2 upregulation that result in possible cardiac issues. Finally, comparisons between these pharmaceutical regulators and exercise are made from mechanistic and physiological viewpoints to elucidate the differences between endogenous and exogenous regulatory effects on Nrf2 and cardiac health.
2. Nrf2 in the Myocardium: Not a Silver Bullet
The heart is obligately aerobic and relies on oxidative phosphorylation to generate the biochemical energy needed for a lifetime of pumping. The coronary arteries supply oxygenated blood to the heart during diastole and increases in oxygen demand by the myocardium are directly related to the heart rate (higher rate = higher oxygen demand and shorter diastole for coronary supply) and saturation of blood by oxygen (to prevent hypoxia). Even at rest, the myocardium consumes 8 to 13 mL of oxygen per 100 g of tissue per minute and ROS from mitochondrial respiration and pro-ROS proteins, such as Nox4, create a pro-oxidative state that requires constant rebalancing to maintain redox homeostasis [1][2]). Xenobiotics may also introduce ROS either by direct chemical action (e.g., nitrosamines from cigarette smoke, fermented foods, or cured meats) or immune response. However, since ROS function as a second-messenger system and have been implicated as crucial regulators of stem cell differentiation and apoptosis/necrosis, tight regulation of the Nrf2-mediated antioxidant response (e.g., via Keap1 direct and Bach1 competitive pathways) is required to maintain such basal messenger activity. Cardiomyocyte differentiation, in particular, is sensitive to ROS, requiring it for progression to maturity, and cardiac-resident stem cells in adults may be similarly affected by imbalanced redox homeostasis, driving them towards hypertrophic or synthetic phenotypes [3].
Of current controversy in cardiac research is the involvement of Nrf2 as a pro-hypertrophic, factor in progressive heart failure. On one side, numerous reports have linked Nrf2 deficiencies to ROS-mediated cardiac hypertrophy related to Angiotensin II, IL-6-mediated inflammation, aortic constriction (TGFβ1/SMAD2 signaling), and obesity-related stress [4][5][6]. Diverse other reports have detailed the role of Nrf2 in preventing cardiomyocyte necrosis, hypertrophy, and fibrosis of the myocardium due to ROS while antioxidant response proteins (e.g., NQO1, SOD1, GPX4) have been found at low expression levels under ischemic cardiomyopathy conditions [7][8]). However, recent evidence that Nrf2 induces progressively maladapted remodeling in the absence of functional autophagy casts doubt on the exploitation of Nrf2 in patients with metabolic disorders or heart disease (Figure 1) [9]. Reports from the Cui research group have indicated that Fyn-mediated nuclear export inhibition is to blame but other yet-discovered factors may also play crucial roles in pathogenesis [9]. Future studies on the effect of autophagy and other regulatory modalities (methylation, sumoylation, etc.) will delineate the thresholds beyond which Nrf2 enhancement becomes problematic for the heart.
Ostensibly, boosting Nrf2 will increase the total antioxidant capacity within the heart and neutralize ROS that perpetuate necrotic and fibrotic pathways, leading to the concept of “the more antioxidant capacity, the better”. In spite of this theory, results from well-controlled clinical trials of supplemental antioxidants (selenium, vitamin E, beta-carotene, etc.) have returned disappointing results where risk was either unchanged or even enhanced [10]. Results from previously reviewed meta-studies with 156,663 and 188,209 total participants found no significant effects of antioxidant/vitamin supplements on cardiovascular risk [10]). However, a recent meta-study of selenium and other antioxidants only found significant risk reduction for selenium across 43 studies (possibly because such minerals, similarly to zinc, are important constituents of antioxidant enzymes and not activators of Nrf2) [11]). Consequently, the Selenium and Vitamin E Cancer Prevention trial (N = 35,533) found that supplementation increased diabetes and prostate cancer risks, while a beta-carotene study did find inverse relationships with lower cardiovascular risk but could not completely rule out the effects of confounding variables (i.e., accidents and injuries) [11][12]. In general, antioxidants have proven to be poor substitutes for generally healthy lifestyle habits (e.g., no tobacco use, moderate diet, moderate exercise, stress reduction, good sleep habits) and excessive antioxidant use is associated with increased all-cause mortality (vitamin E), oxidative stress (ascorbic acid), and cancer risk (vitamin A) [13].
In similar fashion, Nrf2 exogenous enhancers have not shown promise in either preventing or treating cardiovascular diseases and several trials have ended early because of deleterious heart effects after treatment. For this reason, external and sustained enhancement of the antioxidant response out of context with other regulatory factors (e.g., autophagy) could counterintuitively damage the myocardium through pathways not yet fully elucidated (Figure 1). More antioxidant capacity is, in light of these studies, definitely not better.
3. Nrf2 in the Failing Heart: Autophagy as a Keystone Mechanism
Aging and failing hearts experience stiffening from fibrosis caused by immune responses to myocardial necrosis, increased ROS from aging and senescent mitochondria, lipofuscin accumulation from lysosomal degradation, deficiencies in calmodulin signaling/calcium flux (RYR2, SERCA2a) and increased maladaptive remodeling due to high blood pressure that stems from glucose dysregulation and hyperkalemia [14][15][16]. Additionally, autophagic capacity drops as suppression factors like mTOR are overexpressed by chronically high AKT levels while chronic IGF-1 expression, long touted as a youth-sustaining factor, paradoxically ages the heart rapidly as it has been shown to downregulate autophagy by suppression of autophagosome formation and increases in AKT/mTOR [17][18].
Hyperglycemia has been shown to modulate autophagy via AMPK and ROS induction of the ERK/JNK-p53 mechanism [19][20][21]. Additionally, fasting is a potent activator of autophagy even under increased peroxide generation by mitochondria in animals [20][22]. In type 2 diabetics, while initially protective, mitophagy (i.e., autophagy of damaged mitochondria) may eventually drive cells towards reduced energy as mitochondria are damaged by increased metabolic activity and are recycled faster than replacement [23]. However, the loss of autophagic capacity, especially in pancreatic β cells and diabetic hearts, may also be important in progression to end-stage disease [23][24]). Thus, patients who do not possess a fully intact autophagy capacity (e.g., heart failure or type 2 diabetics) may be harmed by artificial Nrf2 enhancement.
Wu et al. recently reported a putative mechanism for this effect in pressure-overloaded hearts that involves dysfunctional autophagy, restricting phosphorylated Fyn and ERK from translocating to the nucleus and downregulating Nrf2 activity that would otherwise restrict angiotensin expression [25]. In such cases, subsequent activation of angiotensin II (Ang-II) receptors by Ang-II production would increase blood pressure and eventual hypertrophy [25]. Additionally, interactions between autophagic control factor p62 and Keap1 mean that reduction in upstream p62/AKT/mTOR result in increased Nrf2 activation and further exacerbation of Ang-II-induced maladaptive remodeling (Figure 1) [26].
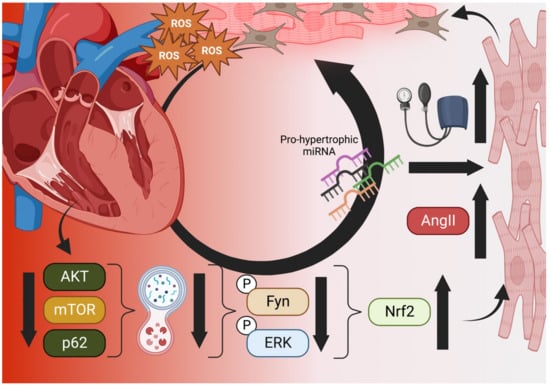
Figure 1. The Vicious Cycle of Nrf2 in Cardiac Hypertrophy. Aged and failing hearts have dysfunctional autophagy (bottom), which cannot downregulate Nrf2 transcription of Angiotensinogen and AngII, increasing blood pressure and mechanical induction of hypertrophy, pro-hypertrophic miRNA, necrosis, and fibroblast activation (right). Hypertrophic cells increase ROS output and decompensation within the heart occurs, increasing the ischemic microenvironment and generating even more ROS in a vicious cycle (top) [25][26]. Created in BioRender.com.
4. Nrf2 in the Kidneys
Because of their role in blood filtering and dependence on over ¼ of cardiac output to function, the kidneys are inextricably linked to the heart [27]. As in the heart, Nrf2 plays an important role in defense against bloodborne sources of ROS (e.g., hyperglycemia, nitrosamines, xenobiotics) and the dense, fine capillary network within kidneys is easily damaged. The primary basal ROS within the kidney are produced by epithelial cells that use mitochondrial respiration for ATP generation that drives glomerular filtration [28]. Such ROS are also important messengers in secondary pathways, including hormone secretion and vascular reactivity [27]. Transient ischemia from heart failure, atherosclerosis, or chronic kidney diseases happens from occluded blood flow and creates excessive ROS from reperfusion injury that can easily damage delicate epithelial cells within the glomerular network and release inflammatory factors that locally propagate ROS production [27]. To compensate, Nrf2, in addition to its suite of ARE-mediated antioxidant enzymes, also produces pentose phosphate that generates NADPH which serves as a local and direct antioxidant [27]. Nrf2 is also protective against heavy metal insult from cadmium or arsenic and glutathione production by Nrf2 may attenuate damage from hyperglycemia in addition to reduction in inflammation through cytokine and NLRP3 inflammasome suppression [29]). Once transcription has been activated, Nrf2 can then be degraded in its canonical manner (proteasome via β-TrCP or Hrd1) and is thus prevented from overaccumulation [29][30]).
Unfortunately, as in the heart, Nrf2 has the potential to inflict harm as a report by Rush et al. (as reviewed by Nezu and Suzuki) revealed that sustained increases of Nrf2 in injured kidneys from treatment with bardoloxone-methyl (CDDO-Me) results in proteinuria and malformed podocyte feet [30][31]. This was thought to be due to inactivation of Keap1 suppression of Nrf2 by electrophilic effect [30]. Thus, in light of the links between deficient autophagy, Nrf2, and myocardial maladaptation, similarly suppressed autophagy in aged or damaged kidneys may also modulate chronic status and permanent damage. However, as sustained autophagy after acute kidney injury has been shown to promote fibrosis, further kidney-specific studies on the relationship between autophagy, Nrf2, and kidney fibrosis with regard to acute and chronic kidney injury microenvironments are required [32].
References
- Hoffman, J.I.E.; Buckberg, G.D. The Myocardial Oxygen Supply:Demand Index Revisited. J. Am. Heart Assoc. 2014, 3, e000285.
- Moris, D.; Spartalis, M.; Tzatzaki, E.; Spartalis, E.; Karachaliou, G.-S.; Triantafyllis, A.S.; Karaolanis, G.I.; Tsilimigras, D.I.; Theocharis, S. The role of reactive oxygen species in myocardial redox signaling and regulation. Ann. Transl. Med. 2017, 5, 324.
- Noseda, M.; Abreu-Paiva, M.; Schneider, M.D. The Quest for the Adult Cardiac Stem Cell. Circ. J. 2015, 79, 1422–1430.
- Chen, D.; Li, Z.; Bao, P.; Chen, M.; Zhang, M.; Yan, F.; Xu, Y.; Ji, C.; Hu, X.; Sanchis, D. Nrf2 deficiency aggravates Angiotensin II-induced cardiac injury by in-creasing hypertrophy and enhancing IL-6/STAT3-dependent inflammation. Biochim. Biophys. Acta 2019, 1865, 1253–1264.
- Syed, A.M.; Kundu, S.; Ram, C.; Kulhari, U.; Kumar, A.; Mugale, M.N.; Mohapatra, P.; Murty, U.S.; Sahu, B.D. Up-regulation of Nrf2/HO-1 and inhibition of TGF-β1/Smad2/3 signaling axis by daphnetin alleviates transverse aortic constriction-induced cardiac remodeling in mice. Free Radic. Biol. Med. 2022, 186, 17–30.
- Gutiérrez-Cuevas, J.; Galicia-Moreno, M.; Monroy-Ramírez, H.C.; Sandoval-Rodriguez, A.; García-Bañuelos, J.; Santos, A.; Armendariz-Borunda, J. The Role of NRF2 in Obesity-Associated Cardiovascular Risk Factors. Antioxidants 2022, 11, 235.
- Zhou, S.; Sun, W.; Zhang, Z.; Zheng, Y. The Role of Nrf2-Mediated Pathway in Cardiac Remodeling and Heart Failure. Oxidative Med. Cell. Longev. 2014, 2014, 260429.
- Lu, Y.; An, L.; Taylor, M.R.G.; Chen, Q.M. Nrf2 signaling in heart failure: Expression of Nrf2, Keap1, antioxidant, and detoxification genes in dilated or ischemic cardiomyopathy. Physiol. Genom. 2022, 54, 115–127.
- Qin, Q.; Qu, C.; Niu, T.; Zang, H.; Qi, L.; Lyu, L.; Wang, X.; Nagarkatti, M.; Nagarkatti, P.; Janicki, J.S.; et al. Nrf2-Mediated Cardiac Maladaptive Remodeling and Dysfunction in a Setting of Autophagy Insufficiency. Hypertension 2016, 67, 107–117.
- Leopold, J.A. Antioxidants and coronary artery disease: From pathophysiology to preventive therapy. Coron. Artery Dis. 2015, 26, 176–183.
- Jenkins, A.D.J.; Kitts, D.; Giovannucci, E.L.; Sahye-Pudaruth, S.; Paquette, M.; Mejia, S.B.; Patel, D.; Kavanagh, M.; Tsirakis, T.; Kendall, C.W.C.; et al. Selenium, antioxidants, cardiovascular disease, and all-cause mortality: A systematic review and meta-analysis of randomized controlled trials. Am. J. Clin. Nutr. 2020, 112, 1642–1652.
- Huang, J.; Weinstein, S.J.; Yu, K.; Männistö, S.; Albanes, D. Serum Beta Carotene and Overall and Cause-Specific Mortality. Circ. Res. 2018, 123, 1339–1349.
- Henkel, R.; Sandhu, I.S.; Agarwal, A. The excessive use of antioxidant therapy: A possible cause of male infertility? Andrologia 2019, 51, e13162.
- Ferreira, J.P.; Butler, J.; Rossignol, P.; Pitt, B.; Anker, S.D.; Kosiborod, M.; Lund, L.H.; Bakris, G.L.; Weir, M.R.; Zannad, F. Abnormalities of Potassium in Heart Failure: JACC State-of-the-Art Review. J. Am. Coll. Cardiol. 2020, 75, 2836–2850.
- Abdellatif, M.; Sedej, S.; Carmona-Gutierrez, D.; Madeo, F.; Kroemer, G. Autophagy in Cardiovascular Aging. Circ. Res. 2018, 123, 803–824.
- Marks, A.R. Calcium cycling proteins and heart failure: Mechanisms and therapeutics. J. Clin. Investig. 2013, 123, 46–52.
- Bitto, A.; Lerner, C.; Torres, C.; Roell, M.; Malaguti, M.; Perez, V.; Lorenzini, A.; Hrelia, S.; Ikeno, Y.; Matzko, M.E.; et al. Long-Term IGF-I Exposure Decreases Autophagy and Cell Viability. PLoS ONE 2010, 5, e12592.
- Yang, M.; Wen, T.; Chen, H.; Deng, J.; Yang, C.; Zhang, Z. Knockdown of insulin-like growth factor 1 exerts a protective effect on hypoxic injury of aged BM-MSCs: Role of autophagy. Stem Cell Res. Ther. 2018, 9, 284.
- Ben Wang, B.; Shi, Y.; Chen, J.; Shao, Z.; Ni, L.; Lin, Y.; Wu, Y.; Tian, N.; Zhou, Y.; Sun, L.; et al. High glucose suppresses autophagy through the AMPK pathway while it induces autophagy via oxidative stress in chondrocytes. Cell Death Dis. 2021, 12, 506.
- Bagherniya, M.; Butler, A.E.; Barreto, G.E.; Sahebkar, A. The effect of fasting or calorie restriction on autophagy induction: A review of the literature. Ageing Res. Rev. 2018, 47, 183–197.
- Tang, Y.; Long, J.; Liu, J. Autophagy: Cancer, Other Pathologies, Inflammation, Immunity, Infection, and Aging; Elsevier: Amsterdam, The Netherlands, 2014; pp. 105–115.
- Salin, K.; Villasevil, E.M.; Anderson, G.J.; Auer, S.K.; Selman, C.; Hartley, R.C.; Mullen, W.; Chinopoulos, C.; Metcalfe, N.B. Decreased mitochondrial metabolic requirements in fasting animals carry an oxidative cost. Funct. Ecol. 2018, 32, 2149–2157.
- Quan, W.; Lim, Y.-M.; Lee, M.-S. Role of autophagy in diabetes and endoplasmic reticulum stress of pancreatic β-cells. Exp. Mol. Med. 2012, 44, 81–88.
- Kanamori, H.; Takemura, G.; Goto, K.; Tsujimoto, A.; Mikami, A.; Ogino, A.; Watanabe, T.; Morishita, K.; Okada, H.; Kawasaki, M.; et al. Autophagic adaptations in diabetic cardiomyopathy differ between type 1 and type 2 diabetes. Autophagy 2015, 11, 1146–1160.
- Wu, W.; Qin, Q.; Ding, Y.; Zang, H.; Li, D.-S.; Nagarkatti, M.; Nagarkatti, P.; Wang, W.; Wang, X.; Cui, T. Autophagy Controls Nrf2-Mediated Dichotomy in Pressure Over-loaded Hearts. Front. Physiol. 2021, 13, 12.
- Jiang, T.; Harder, B.; Rojo de la Vega, M.; Wong, P.K.; Chapman, E.; Zhang, D.D. p62 links autophagy and Nrf2 signaling. Free Radic. Biol. Med. 2015, 88 Pt B, 199–204.
- Nezu, M.; Suzuki, N. Roles of Nrf2 in Protecting the Kidney from Oxidative Damage. Int. J. Mol. Sci. 2020, 21, 2951.
- Kriz, W. Adenosine and ATP: Traffic regulators in the kidney. J. Clin. Investig. 2004, 114, 611–613.
- Guerrero-Hue, M.; Rayego-Mateos, S.; Vázquez-Carballo, C.; Palomino-Antolín, A.; García-Caballero, C.; Opazo-Rios, L.; Morgado-Pascual, J.L.; Herencia, C.; Mas, S.; Ortiz, A. Protective Role of Nrf2 in Renal Disease. Antioxidants 2020, 10, 39.
- Nezu, M.; Suzuki, N. Nrf2 activation for kidney disease treatment—A mixed blessing? Kidney Int. 2021, 99, 20–22.
- Rush, B.M.; Bondi, C.D.; Stocker, S.D.; Barry, K.M.; Small, S.A.; Ong, J.; Jobbagy, S.; Stolz, D.B.; Bastacky, S.I.; Chartoumpekis, D.V. Genetic or pharmacologic Nrf2 activation increases proteinuria in chronic kidney disease in mice. Kidney Int. 2021, 99, 102–116.
- Tang, C.; Livingston, M.J.; Liu, Z.; Dong, Z. Autophagy in kidney homeostasis and disease. Nat. Rev. Nephrol. 2020, 16, 489–508.
More
Information
Subjects:
Physiology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
713
Revisions:
2 times
(View History)
Update Date:
15 Dec 2022
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No