Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Mahesh Rachamalla | -- | 3399 | 2022-12-12 17:36:47 | | | |
| 2 | Conner Chen | Meta information modification | 3399 | 2022-12-13 10:08:05 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Rachamalla, M. The Arsenic on Male Reproductive System in Animals. Encyclopedia. Available online: https://encyclopedia.pub/entry/38612 (accessed on 25 June 2026).
Rachamalla M. The Arsenic on Male Reproductive System in Animals. Encyclopedia. Available at: https://encyclopedia.pub/entry/38612. Accessed June 25, 2026.
Rachamalla, Mahesh. "The Arsenic on Male Reproductive System in Animals" Encyclopedia, https://encyclopedia.pub/entry/38612 (accessed June 25, 2026).
Rachamalla, M. (2022, December 12). The Arsenic on Male Reproductive System in Animals. In Encyclopedia. https://encyclopedia.pub/entry/38612
Rachamalla, Mahesh. "The Arsenic on Male Reproductive System in Animals." Encyclopedia. Web. 12 December, 2022.
Copy Citation
Epidemiological studies have suggested that arsenic is one of the most hazardous reproductive toxicants present in the environment, which is significantly accumulated in the reproductive tissues like the testes, epididymis, seminal vesicle, and prostate gland. Arsenic exposure exerted cellular and molecular perturbations such as oxidative stress, inflammation, induction of autophagy, and apoptosis, which obstructed male gonadal development and led to reproductive dysfunction in humans and animals.
arsenic
metalloid
phytonutrients
reproductive toxicity
molecular mechanisms
1. Introduction
Arsenic (As) is a naturally occurring toxic metalloid with odourless, colourless, and tasteless properties. The most commonly available forms of arsenic include inorganic As, organic As, and arsine gas [1]. Arsenic is ranked as the 20th most abundant element in the terrestrial, 14th in the marine, and 12th in the human ecosystem. It possesses significant health concerns due to its ubiquitous existence, discharged into the environment from volcanic and industrial activities [2][3]. Charles Dickens used arsenic as a tonic, called Fowler’s solution (potassium arsenate in water). It was also employed to treat leukaemia, psoriasis, and chronic bronchial asthma [4][5]. As is a well-known poison whose use is restricted to the production of pesticides, herbicides, cotton desiccants, and exfoliants in agriculture, applied as doping material in the semiconductor industry, bronzing, pyrotechnics, as well as the manufacturing of special kinds of glasses and preservation of wood [6][7]. As is found naturally in both trivalent and pentavalent forms, but the pentavalent form is much more common and is the best form for eliminating arsenic from biological systems [8][9]. The commonly available pentavalent forms of arsenic are arsenic pentoxide, arsenic acid, sodium arsenate, lead arsenate, and calcium arsenate [10][11]. More than 200 million people are adversely affected by toxic responses to As, which acknowledges research and mitigation methods to control human arsenic exposure [12]. Since it is ubiquitous, humans are exposed via groundwater, food, and industrial and anthropogenic sources [13]. The largest source of As exposure for people is drinking water; the World Health Organization (WHO) and Environmental Protection Agency (EPA) recommend a maximum concentration of 10 µg/L for As in drinking water [14]. As exposure through the air is negligible, effects are observed when the air comprises a mixture of arsenite and arsenate [14]. As mobilisation in drinking water results from microbial reactions such as oxidizing arsenite or reducing arsenate [15]. Furthermore, arsine gas, gallium arsenide, glass manufacturing facilities, and coal-fired power plants are some of the occupational sources of arsenic [16]. Furthermore, elevated levels of arsenic were found in commonly consumed foods such as grains, vegetables, and rice, as well as significant concentrations in meat products such as beef, poultry, and shellfish [17]. Several countries around the world are highly exposed and vulnerable to arsenic toxicity, including Cambodia, China, India, Mexico, Pakistan, the United States, Vietnam, and East Croatia [18][19][20]. Chronic exposure in Bangladesh and Taiwan, as well as low-level exposure in the United States, has resulted in the onset of type II diabetes [21][22]. Pregnant women in the Chilean region who were exposed to As in their drinking water for an extended period of time experienced a decrease in baby birth weight [23]. Cancer, genetic changes, and dermatological diseases have been reported in Argentina when As concentrations reach 100 µg/L [24]. In Taiwan, As contamination in drinking water increases the risk of lung, kidney, and bladder cancer [25]. As levels in the aquifer have the greatest impact on a few provinces in India (Uttar Pradesh, Bihar, and West Bengal) and Bangladesh [26][27][28][29].
As is designated by the International Agency for Research on Cancer (IARC) as a group-I carcinogen that also raises the risk of bladder, lung, kidney, and liver cancer [25][30][31][32].
As induces carcinogenesis by epigenetic modifications in miRNA expression [33][34], DNA methylation, and histone modifications [35]. Low doses of As in the form of orpiment (AS2S3), realgar (AS4S4), and especially arsenolite (contains arsenic trioxide, AS2O3) have been used as a therapeutic agent in Iranian traditional medicine since Avicenna (1023 A.D.) [36][37]. Interestingly, As was therapeutically used to treat chronic myelogenous leukaemia (CML) until radiation and chemotherapy took over [9]. As has also demonstrated significant success in the treatment of newly diagnosed and relapsed individuals with acute promyelocytic leukaemia (APL) [38]. Acute exposure to As causes nausea, vomiting, abdominal pain, and severe diarrhoea, whereas prolonged exposure causes damage to multiple organs [11][39]. Interestingly, Calabrese and Baldwin (2009) found hormetic dose–response relationships with inorganic compounds including arsenic showed that mechanisms associated with arsenic may decipher the new dimensions. Recently, Ommati et al., found that in mature F1 male mice, the spermatogenic index was higher in low-dose (0.2 ppm) animals, demonstrating another possible hormesis effect, but a dose-dependent decrease in the spermatogenic index was found at higher doses (2 and 20 ppm) of As2O3. It is interesting to note that Calabrese and Baldwin (2009) discovered hormetic dose–response relationships with inorganic compounds, one of which was arsenic. These relationships revealed that mechanisms associated with arsenic may be able to decipher new dimensions [40]. Ommati et al. (2019) recently found that in mature F1 male mice, the spermatogenic index was higher in low-dose (0.2 ppm) animals, demonstrating another possible hormesis effect, whereas a dose-dependent decrease in the spermatogenic index was found at higher doses (2 and 20 ppm) of As2O3 [41]. In addition, a hormesis effect was seen in female mice, as indicated by a decrement in thiobarbituric acid reactive substances (TBARS) content and an increase in ovarian mammalian target of rapamycin (mTOR) gene expression level, both of which occurred at lower doses of As2O3 [42]. They also observed the hormesis effect in the hypothalamic–pituitary–gonadal (HPG axis) of pubertal male offspring when exposed to low levels of As2O3 (0.2 ppm) [37].
2. Exposure to Arsenic in the Environment
Arsenic exposure in humans is constantly rising in drinking water, food, and industrial sources. Higher arsenic levels were reported in drinking water in most countries and were higher than the specified limits of WHO and EPA. Cultivation of rice with As-contaminated soil and the use of As-contaminated water to cook rice are the major contributors to As exposure in cooked rice [43]. Finfish, shellfish, and seaweed are the most common sources of As exposure in humans through seafood [44]. As depicted in Table 1, urine, hair, and drinking water samples collected from children and adults from As-contaminated regions in Argentina, Uruguay, India, Pakistan, and Spain showed higher levels of arsenic. Accumulation levels of arsenic in various foods like rice, meat, cereals, and vegetables in different regions of the world were presented in Table 2. Overall, there is a significant risk of cancer in children and adults due to the build-up of arsenic in the diet.
Table 1. Compilation of research studies showing the effect of arsenic exposures among diverse demographics in various parts of the world.
| S. No | Countries | Population (Subjects) | Sample Size | Sample | Detected Levels of Arsenic | Reference |
|---|---|---|---|---|---|---|
| 1 | Argentina | Children (3–15 years) | 101 | Hair, Urine | 110–1311 μg/kg | [45] |
| 2 | Montevideo, Uruguay | Children (5–8 years) | 328 | Drinking water | 9.9 μg/L | [46] |
| 0.45 μg/L | ||||||
| 3 | Shaanxi province, China | Adults | 96 | Drinking water | 4.52 µg/L | [47] |
| Indoor air | 0.03 mg/m3 | |||||
| Soil | 14930 mg/kg | |||||
| 4 | Inner Mongolia | Adults | 96 | Drinking water | 144.71 µg/L | |
| Soil | 10190 mg/kg | |||||
| 5 | Villages of Pakistan | Children (≤16 years) | 223 | Ground water | 15. 63 µg/kg/day (arsenate) 0.09 µg/kg/day (arsenite) | [48] |
| Adult | 15.07 µg/kg/day (arsenate) 0.26 µg/kg/day (arsenite) | |||||
| 6 | Indae metal mine area | Residents (mean age of 66.8 years) | 50 | Urine | Arsenite (1.45 µg/L), arsenate (0.74 µg/L), MMA (2.43 µg/L), DMA (27.63 µg/L), and arsenobetaine (88.62 µg/L) | [49] |
| 7 | Spain | Children (4–5 years) | 400 | Urine | 2.74–7.54 µg/L | [50] |
Table 2. Evidence from research findings indicates the presence of arsenic in grains, vegetables, dairy products, and meat which increases the risk of cancer.
| S. No | Country | Sample (s) | Arsenic Levels | Reference |
|---|---|---|---|---|
| 1 | Kolkata, India | Rice, grain, and vegetable | 76 µg/kg and 41.4 µg/kg | [51] |
| 2 | UK | Rice | 130 μg/kg | [52] |
| 3 | Kunming, China | Rice | 3520 µg/kg | [53] |
| 4 | Japan | Rice, hijiki | 19 µg/kg and 59 µg/kg | [54] |
| 5 | Pakistan | Raw rice | 92.5 ± 41.88 μg/kg | [55] |
| Cooked rice | 79.21 ± 76.42 μg/kg | |||
| Wheat | 116.38 ± 51.38 μg/kg | |||
| 6 | West Bengal, India | Boro and Aman rice | 194 μg/kg and 156 μg/kg | [51] |
| Arum and radish | 780 and 674 μg/kg | |||
| Urine | 154–276 µg/L | |||
| 7 | Ngari area, Western Tibet | Barley | 180 ± 210 µg/kg | [56] |
| Vegetable | 400 ± 570 µg/kg, | |||
| Meat | 210 ± 160 µg/kg | |||
| Dairy products | 180 ± 80 µg/kg | |||
| Daily intake | 3600 µg/kg | |||
| 8 | Kurdistan province, Iran | Sheep meat | 230 ± 140 µg/kg | [57] |
| Beef meat | 200 ± 90 µg/kg | |||
| Turkey meat | 880 ± 210 µg/kg | |||
| Ostrich meat | 850 ± 130 µg/kg |
3. Metabolism of Arsenic
In many organisms, along with humans, the inorganic form of As gets rapidly reduced from arsenate (pentavalent) to arsenite (trivalent) in the blood, facilitated by glutathione (GSH) [58]. The latter is 2–10 folds more toxic and rapidly taken up by the cells than the former [10]. Once absorbed, arsenite binds to globin of Hb- and -SH-containing proteins such as Glutathione and Cysteine and is distributed to the skin, hair, and mucosa owing to thiol-cystine amino acids. Skin, hair, epithelium of gastrointestinal tract (GIT), and epididymis had the highest As retention [59][60][61][62]. Aquaglyceroporins (AQPs) are involved in transporting As into cells [63], whereas efflux is carried out by major facilitator superfamily (MFS) transporters and ATP-binding cassette (ABC) transporters [64]. As shown in Figure 1, initially, biotransformation of arsenic involves oxidative methylation of arsenite to arsenate, mediated by the enzyme arsenite methyltransferase (As3MT), a primary methyl donor, followed by oxidation, arsenite to a pentavalent metabolite of arsenate, monomethylarsonic acid (MMA), and dimethylarsinic acid (DMA) [65][66]. MMA and DMA were taken up less effectively by organs and tissue than arsenite and rapidly excreted into urine [67]. In contrast, evidence suggests that metabolites of As tend to induce chromosomal mutations [68] and show genotoxicity [69]. Methylated forms of As were also found in human urine, natural water, and bird eggshells [70].
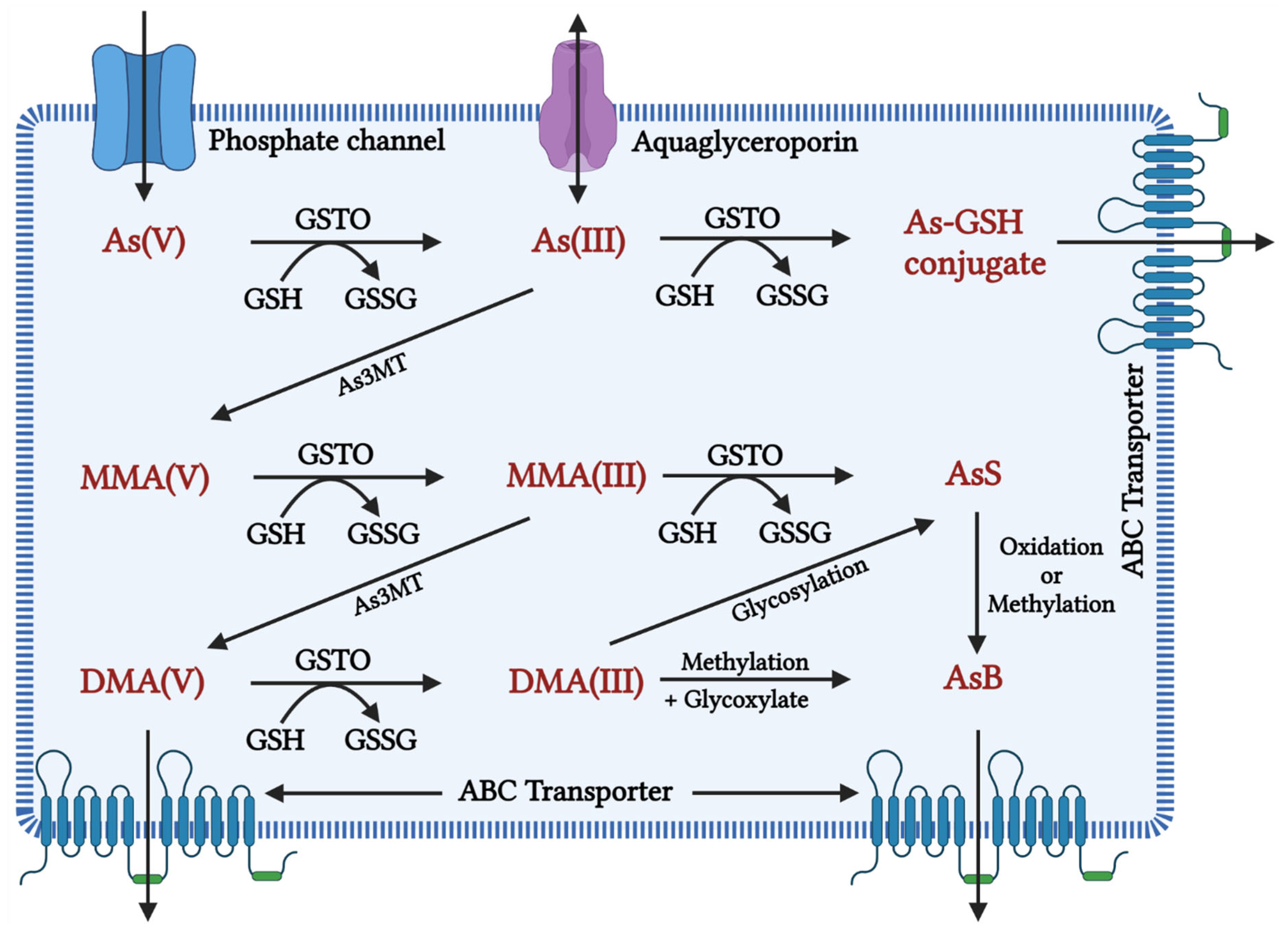
Figure 1. Inorganic arsenic conversion pathways into mono-, di-, and trimethylated products when the toxic form of As(III) is taken up by a cell, it is methylated to MMA (monomethylarsonic acid) and DMA (dimethylarsinic acid) with the help of the enzyme arsenic methyl transferase (AsMT) and can then be eliminated from the body through urine. However, in a pathological state, SAM depletion causes As(III) aggregation, affecting cellular homeostasis in a variety of ways, including oxidative balance, inflammation, and genetic and epigenetic processes. This disruption in homeostasis causes cellular damage and, eventually, cell death. As(V): inorganic pentavalent arsenic; As(III): inorganic trivalent arsenic; As3MT: arsenite methyltransferase, MMA(V): methyl arsonate; MMA(III): monomethylarsonous acid; DMA(V): dimethyl arsenate; DMA(III): dimethylarsinous acid; GSH: glutathione; GSTO: Glutathione S-transferase omega, GSSG: Glutathione disulfide As-GSH: Arsenic glutathione; AsS: Arsenic sulphide; AsB: Arsenobetaine; ABC transporter: ATP-binding cassette transporters; AS3MT: arsenite methyltransferase.
4. Arsenic-Induced Oxidative Stress and DNA Damage
At a biochemical level, inorganic arsenic in the pentavalent state replaces the phosphate in several reactions, the trivalent form of inorganic and organic (methylated) arsenic reacts with critical thiols and inhibits their activity [65]. As induces the formation of reactive oxygen species (ROS), enhances free radicals, and diminishes the activity of thiol group-rich antioxidants such as glutathione (GSH) [71]. Production of ROS will take place during the generation of intermediate arsine species [72]. Methylated arsenicals cause the release of iron from ferritin and initiate the production of hydroxy radicals and progress ferroptosis [73]. Arsenic shows mitochondrial toxicity by inhibiting succinic dehydrogenase activity and uncoupling oxidative phosphorylation with the production of O2, which gives rise to other forms of ROS [74]. Oxidative stress leading to the activation of ERK/AKT/NF-kβ pathway is one of the primary molecular mechanisms involved in arsenic-induced male reproductive toxicity [10]. Mitogen-activated protein kinases (MAPKs) are proteins that regulate the signalling of cells in response to environmental stimuli. The ERK signalling pathway is involved in a variety of male reproductive functions, such as spermatogenesis and Sertoli cell function. Activation of ERK1/2 inhibits the function of Sertoli cells and increases apoptosis in the testes. Protein kinase B (AKT) regulates oxidative stress in conjunction with the immune system by regulating cell growth, survival, proliferation, and inflammation. MAPK and AKT both directly phosphorylate nuclear factor kappa B (NF-B), which increases NF-ĸB binding to target DNAs and the expression/activity of NF-ĸB-controlled genes. Spermatogenesis and the function of Sertoli cells in the testes are governed by the transcription factor NF-ĸB. Activation of NF-ĸB inhibits spermatogenesis in both humans and mice. Arsenic activated the ERK/AKT/NF-B signalling pathways in numerous cell types. The expression of ERK1/2, IKK, PI3K, AKT, and NF-B, as well as the phosphorylation of ERK/AKT, increased in rats exposed to sodium arsenite (1, 5, or 25 mg/L) for 6 months. This causes reproductive toxicity via ERK/AKT/NF-kB signalling [75][76][77][78][79][80][81][82]. Figure 2 depicts As-induced oxidative stress mechanisms.
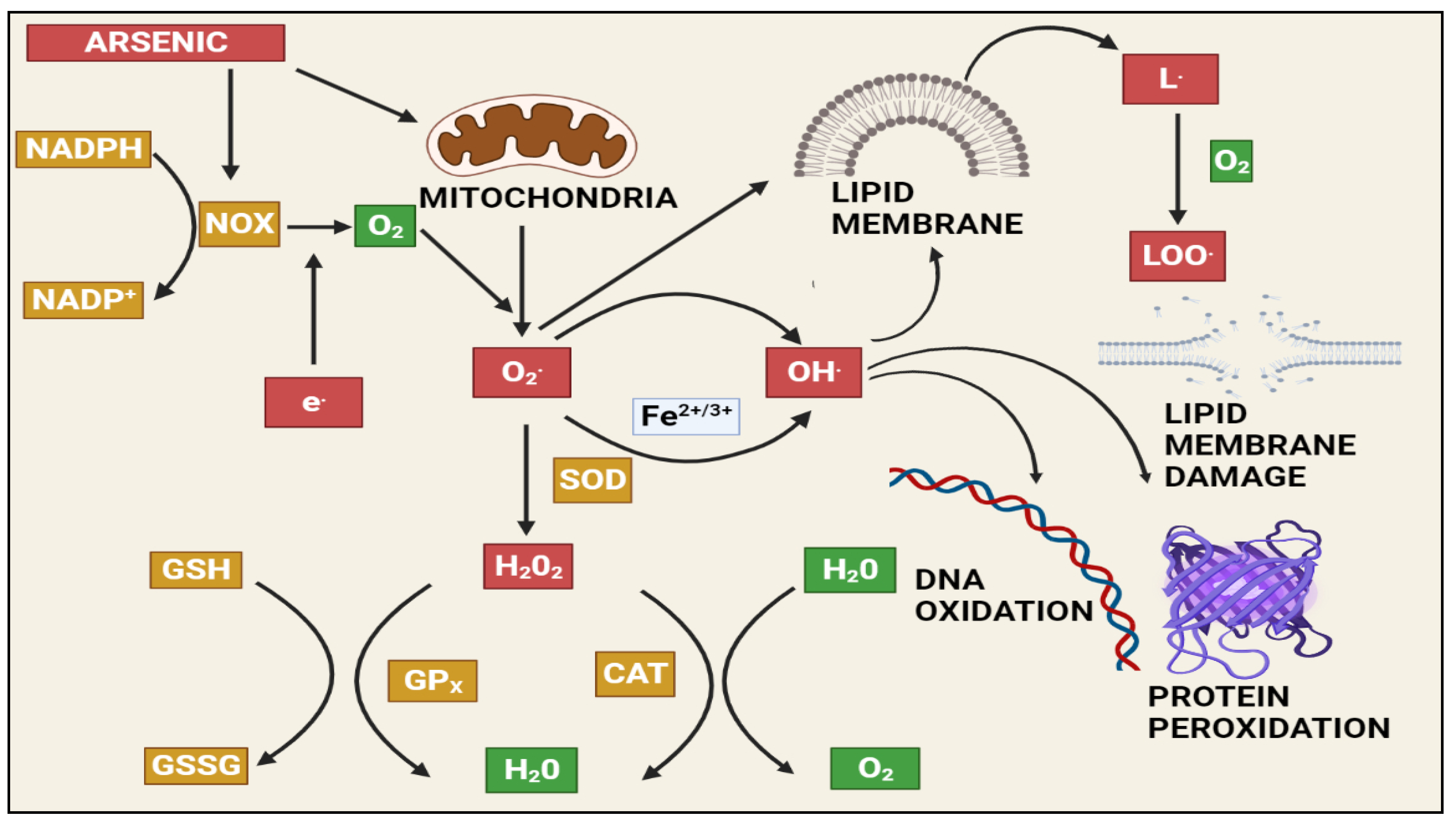
Figure 2. The effects of arsenic-induced mitochondrial reactive oxygen species generation. Arsenic generates a significant amount of ROS, primarily through complexes I and II of the electron transport chain (ETC). The ETC produces superoxide radicals, which react with other radicals in the cell to form stable and long-lived reactive species that damage macromolecules and induce apoptosis via various pathways. NAPDH: Nicotinamide adenine dinucleotide phosphate (NADP+), NADPH oxidase (NOX), GSH: Gluthathione, GSSG: Glutathione disulfide, GPx: Glutathione peroxidase, SOD: Superoxide dismutase, CAT: Catalase, H2O2: Hydrogen peroxide, LOO: lipid peroxy radical.
5. Effects of Arsenic on the Male Reproductive System in Animals
Epidemiological studies have suggested that arsenic is one of the most hazardous reproductive toxicants present in the environment, which is significantly accumulated in the reproductive tissues like the testes, epididymis, seminal vesicle, and prostate gland [83][84]. The inhibition of testosterone biosynthesis by arsenic is depicted in Figure 3. Arsenic exposure exerted cellular and molecular perturbations such as oxidative stress, inflammation, induction of autophagy, and apoptosis, which obstructed male gonadal development and led to reproductive dysfunction in humans and animals [85][86]. Arsenic has been shown to damage the histology of various tissues, such as the liver, brain, and kidney [87][88]. Notably, arsenic exposure during development significantly altered tight junctions’ proteins, leading to an increase in blood–brain barrier permeability [89]. As a result, the age-dependent inhibition of the PI3K/Akt/mTOR signalling pathway may contribute to the induction of autophagy and the facilitation of arsenic transfer through the cerebellum’s cerebral cortex and hippocampus leaky blood–brain barrier [90]. Recent studies from the Ommati research group also shed some light on hypothalamic–pituitary–gonadal (HPG) axis disruption and subsequent toxicity in mice and its continued impact on offspring as well (refer to Figure 3) [37][41][91][92][93]. Similarly, the blood–testis barrier (BTB) is one of the most impermeable blood–tissue barriers in mammals. It divides the seminiferous epithelium into basal (intraluminal) and apical (intraluminal) compartments. Meiosis I and II, spermiogenesis, and spermiation all occur in a specialised microenvironment behind the BTB in the apical compartment, whereas spermatogonial renewal and differentiation, as well as cell cycle progression up to the preleptotene spermatocyte stage, occurs outside the BTB in the basal compartment of the epithelium. However, the BTB is not a static ultrastructure. Instead, it undergoes extensive remodelling during stage VIII of the seminiferous epithelial cycle of spermatogenesis to allow preleptotene spermatocytes to pass through the BTB. However, the BTB’s immunological barrier cannot be compromised, even temporarily, during the epithelial cycle in order to prevent the production of antibodies against meiotic and postmeiotic germ cells. Adhesion protein complexes (e.g., occludin-ZO-1, N-cadherin—catenin, claudin-5-ZO-1), steroids (e.g., testosterone, estradiol-17), nonreceptor protein kinases (e.g., focal adhesion kinase, c-Src, c-Yes), polarity proteins induce testicular damage via their initial actions at the BTB, resulting in germ-cell loss, reduced sperm count, and male infertility or subfertility. Metallothioneins [cysteine-rich low molecular weight metal-binding proteins localised to the membrane of the Golgi apparatus that protect cells from cytotoxicity of essential heavy metals (such as zinc, selenium, and copper) and non-essential heavy metals (such as arsenic, mercury, silver, and cadmium) by binding to these metals via the thiol groups] are largely responsible for heavy metal accumulation in the body. As a result, significant and harmful amounts of heavy metal accumulation can accumulate in a person over time, exceeding the capacity of metallothioneins in the process [94][95][96][97][98][99].
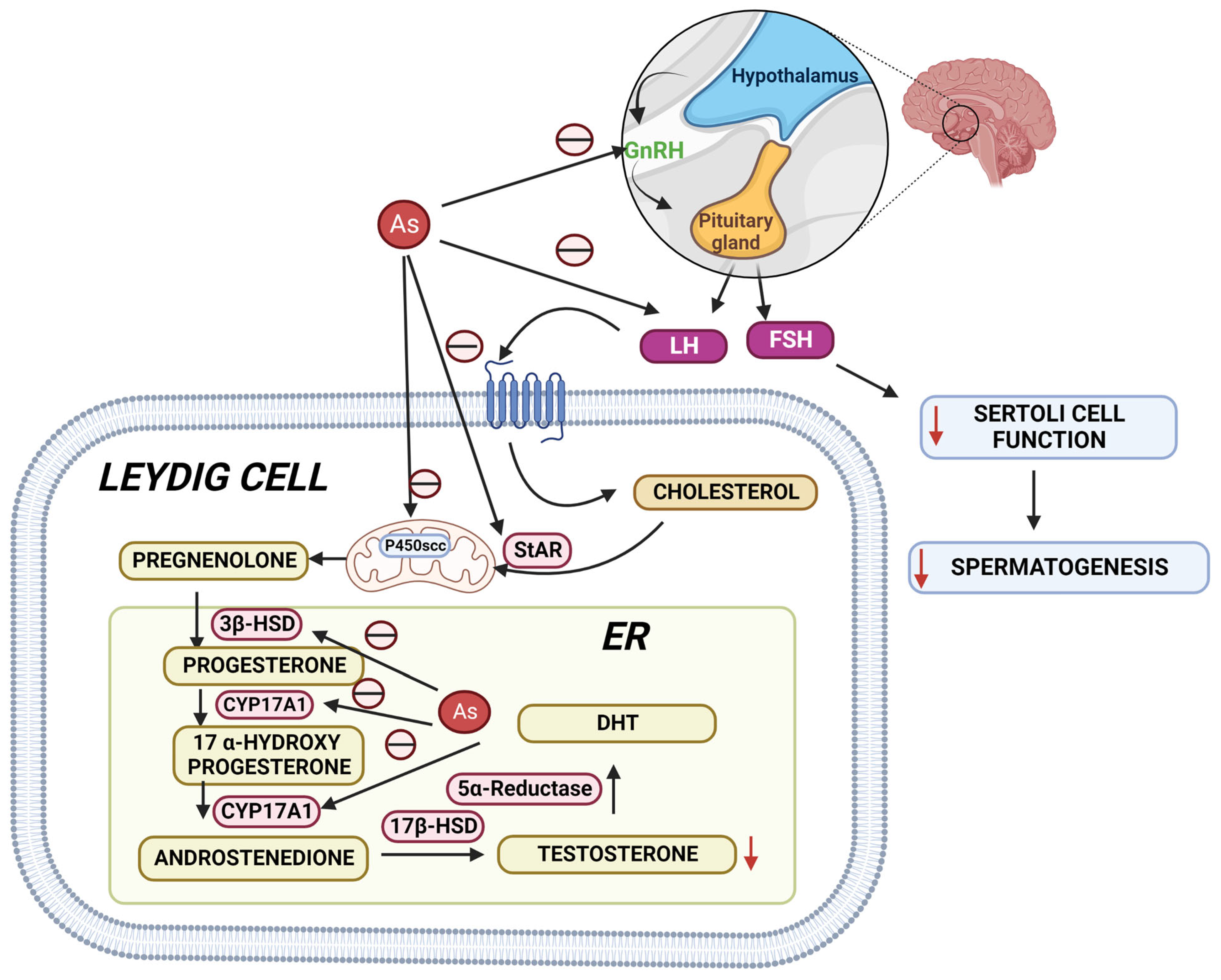
Figure 3. Arsenic’s mechanisms show that it alters the hypothalamic–pituitary–gonadal (HPG) axis, resulting in a decrease in testosterone biosynthesis, Sertoli cell activity, and spermatogenesis. StAR: steroidogenic acute regulatory protein; 3HSD: 3-beta (β)-hydroxysteroid dehydrogenase; CYP17A1: Cytochrome P450 17A1; 17HSD: 17-beta (β)-hydroxysteroid dehydrogenase; DHT: Dihydrotestosterone.
Testicular histopathological investigations showed that sodium arsenite decreased seminiferous tubule diameter in Wistar rats [16] and outbred Institute of Cancer Research (ICR) mice led to a significant decrease in the lumen in the arsenic-treated group compared to the control group [100]. Vacuolisation, acidophilic cells, and epithelial degeneration were associated with increased inflammatory cytokines in male rats exposed to sodium arsenite through drinking water [101]. Moreover, sodium arsenite exhibits severe damage to the testicular structure and elevated cleaved caspase 3 (CC3) which is an apoptotic marker in the cluster differentiation 1 (CD1) mouse testes cell line [102]. Increased expression of CC3 also indicates increased apoptosis. Together, sodium arsenite and arsenate at a concentration of 0.01 and 10 mg/L in drinking water showed a decrease in catalase activity and vacuolisation of seminiferous tubules in rats [103]. Arsenic exposure inhibited the spermatogenesis process and decreased the mobility and viability of sperm [104]. An increase in apoptotic spermatozoa has been observed with arsenic exposure, the central mechanism involved in arsenic-induced decreased sperm count [105]. Sodium arsenite exposure at a concentration of 10 mg/L through drinking water for eight weeks exhibited a decrease in sperm counts and enhanced sperm head abnormalities, which led to an increase in the infertility risk and pre-implantation loss in Wistar rats [106]. Moreover, in mice, arsenic trioxide at doses of 0.3 and 3 mg/kg s.c. for 35 days reduced the number of spermatozoa, increased epithelial aberration and exfoliation of germ cells in the tubule lumen, and altered the nucleus/cytoplasm ratio of Leydig cells [107]. In mouse testes, arsenic trioxide at concentrations of 0.2, 2, and 20 ppm in drinking water for six months impaired sperm motility and affected the ultra-structure of the acrosome structure and sperm tail by downregulating the protein expression of DPY19L2, AKAP3, CFAP44, and SPAG16 [108]. Similarly, arsenic trioxide inhibited the expression of the DDX25 and CRM1 mRNAs, as well as the downstream proteins HMG2, PGK2, and H4 necessary for spermatogenesis in mice [109]. Oral exposure to arsenic trioxide at doses of 0.3 and 30 µg/kg for 15 days resulted in a dose-dependent frequency of sperm production in mice with aberrant head morphology [110]. Diabetic rats exposed to sodium arsenite at a concentration of 10 mg/L in drinking water for 40 days caused a drop in serum testosterone, sperm counts, motility, morphology, and acrosomal and plasma membrane structure [111]. Decreases in sperm count and viability with increased arsenic accumulation, lipid peroxidation, and protein carbonylation in the testes have been observed by administration of sodium arsenate at a concentration of 10, 25, 50, 100, and 200 ppm for 40 days through drinking water in mice [112]. Rats exposed to sodium arsenite at doses of 1, 5, and 25 mg/L through drinking water for 6 months showed compromised sperm counts and motility, and testosterone and altered 19 proteins related to reproduction such as Vdac3, Prkaca, Hspa41, Spaca1, Ma1b, Gpx4, Safb1, Trim28, Rbp1, Hsd11b1, Mapk3, Gpd2, Ace, Hspa11, Dnaja1, Ybx3, Smcp, Nasp, and Cabs1 were altered [79].
6. Pre-Natal Arsenic Exposure Induced Male Reproductive Toxicity in Animals
The growing evidence suggests that parental and/or pre-natal arsenic exposure to animals resulted in post-natal developmental toxicity. In-utero exposure to sodium arsenite from embryonic day-10 to day-18 at a concentration of 10 ppb induced an increase in leptin levels, and at 42.5 ppm reduced the litter size compared to control mice [113]. Chronic As trioxide exposure to parental male at a dose of 1 mg/L showed genotoxic damage in F0-F3, altered methylation patterns, changes in reproductive parameters, morphological damage in the ovaries (F0 and F1) and testicles (F1–F3), and compromised sperm quality (F0-F3, except F2) [114]. Exposure of sodium arsenite at the dose of 10 mg/L in drinking water to pregnant females from GD1-21 affected body weight and initial sexual development in male pups and relative anogenital distance also showed changes in the expression of SOD1, SOD2, CAT, and GSTK1 gene in male pup rat [115]. Administration of sodium arsenite at a concentration of 10 mg/L showed lower sperm production, sperm count, motility, and quality in the epididymis of rats [116]. Daily sperm production and the number of spermatids in the rat epididymis were reduced after oral treatment of sodium arsenite at doses of 0.01 and 10 mg/L for 56 days [117]. Lead at the dose of 819 mg/L was exposed to pregnant rats through drinking water until weaning, followed by the exposure of arsenic to male offspring at a dose of 2.3 mg/L, which showed a decrease in daily sperm production, relative weights of testes, epididymis, seminiferous tubules, and prostate, and decreased the activity of 3β-HSD and 17β-HSD in male offspring [118]. Sodium fluoride and sodium arsenite at 100 mg/L and 50 mg/L, respectively, via drinking water during the pre-pregnancy period, decreased the testicular weights, serum FSH, LH, and testosterone levels, and increased Beclin1 and LC3 expressions in Sprague Dawley (SD) rats’ testes [119].
References
- Rossy, K.M.; Janusz, C.A.; Schwartz, R.A. Cryptic Exposure to Arsenic. Indian J. Dermatol. Venereol. Leprol. 2005, 71, 230–235.
- Verma, N.; Rachamalla, M.; Sravan Kumar, P.; Dua, K. Assessment and Impact of Metal Toxicity on Wildlife and Human Health; Woodhead Publishing: Sawston, UK, 2023; pp. 93–110.
- Mandal, B.K.; Suzuki, K.T. Arsenic Round the World: A Review. Talanta 2002, 58, 201–235.
- Antman, K.H. Introduction: The History of Arsenic Trioxide in Cancer Therapy. Oncologist 2001, 6, 1–2.
- Zhu, J.; Chen, Z.; Lallemand-Breitenbach, V.; de Thé, H. Timeline: How Acute Promyelocytic Leukaemia Revived Arsenic. Nat. Rev. Cancer 2002, 2, 705–714.
- Tamaki, S.; Frankenberger, W.T. Environmental Biochemistry of Arsenic. Rev. Environ. Contam. Toxicol. 1992, 124, 79–110.
- Sears, M.E.; Kerr, K.J.; Bray, R.I. Arsenic, Cadmium, Lead, and Mercury in Sweat: A Systematic Review. J. Environ. Public Health 2012, 2012, 1–10.
- Leist, M.; Casey, R.; Caridi, D. The management of arsenic wastes: Problems and prospects. J. Hazard. Mater. 2000, 76, 125–138.
- Waxman, S.; Anderson, K.C. History of the Development of Arsenic Derivatives in Cancer Therapy. Oncologist 2001, 6, 3–10.
- Renu, K.; Madhyastha, H.; Madhyastha, R.; Maruyama, M.; Vinayagam, S.; Gopalakrishnan, A.V. Review on molecular and biochemical insights of arsenic-mediated male reproductive toxicity. Life Sci. 2018, 212, 37–58.
- Kim, Y.-J.; Kim, J.-M. Arsenic Toxicity in Male Reproduction and Development. Dev. Reprod. 2015, 19, 167–180.
- Chen, Q.Y.; Costa, M. Arsenic: A Global Environmental Challenge. Annu. Rev. Pharmacol. Toxicol. 2021, 61, 47–63.
- Abdul, K.S.M.; Jayasinghe, S.S.; Chandana, E.P.S.; Jayasumana, C.; De Silva, P.M.C.S. Arsenic and human health effects: A review. Environ. Toxicol. Pharmacol. 2015, 40, 828–846.
- Chung, J.-Y.; Yu, S.-D.; Hong, Y.-S. Environmental Source of Arsenic Exposure. J. Prev. Med. Public Health 2014, 47, 253–257.
- Oremland, R.S.; Stolz, J.F. The Ecology of Arsenic. Science 2003, 300, 939–944.
- Ramos, A.T.D.A.; Diamante, M.A.S.; Lamas, C.D.A.; Dolder, H.; Predes, F.D.S. Morphological and morphometrical changes on adult Wistar rat testis caused by chronic sodium arsenite exposure. Environ. Sci. Pollut. Res. 2017, 24, 27905–27912.
- Xu, W.; Bao, H.; Liu, F.; Liu, L.; Zhu, Y.-G.; She, J.; Dong, S.; Cai, M.; Li, L.; Li, C.; et al. Environmental exposure to arsenic may reduce human semen quality: Associations derived from a Chinese cross-sectional study. Environ. Health 2012, 11, 46.
- Quansah, R.; Armah, F.; Essumang, D.K.; Luginaah, I.; Clarke, E.; Marfoh, K.; Cobbina, S.J.; Nketiah-Amponsah, E.; Namujju, P.B.; Obiri, S.; et al. Association of Arsenic with Adverse Pregnancy Outcomes/Infant Mortality: A Systematic Review and Meta-Analysis. Environ. Health Perspect. 2015, 123, 412–421.
- Podgorski, J.; Berg, M. Global threat of arsenic in groundwater. Science 2020, 368, 845–850.
- Vidosavljevic, M.; Puntaric, D.; Gvozdic, V.; Vidosavljevic, D.; Juric, D.; Begovic, L. Assessment of Arsenic in Hair of the Inhabitants of East Croatia—Relationship to Arsenic Concentrations in Drinking Water. Water 2022, 14, 1558.
- Rahman, M.; Tondel, M.; Ahmad, S.A.; Axelson, O. Diabetes Mellitus Associated with Arsenic Exposure in Bangladesh. Am. J. Epidemiol. 1998, 148, 198–203.
- Kile, M.L.; Christiani, D.C. Environmental Arsenic Exposure and Diabetes. JAMA-J. Am. Med. Assoc. 2008, 300, 845–846.
- Hopenhayn, C.; Ferreccio, C.; Browning, S.R.; Huang, B.; Peralta, C.; Gibb, H.; Hertz-Picciotto, I. Arsenic Exposure from Drinking Water and Birth Weight. Epidemiology 2013, 14, 593–602.
- Bardach, A.E.; Ciapponi, A.; Soto, N.; Chaparro, M.R.; Calderon, M.; Briatore, A.; Cadoppi, N.; Tassara, R.; Litter, M.I. Epidemiology of Chronic Disease Related to Arsenic in Argentina: A Systematic Review. Sci. Total Environ. 2015, 538, 802–816.
- Smith, A.H.; Lopipero, P.A.; Bates, M.N.; Steinmaus, C.M. Arsenic Epidemiology and Drinking Water Standards. Science 2002, 296, 2145–2146.
- Rahman, M.M.; Chowdhury, U.K.; Mukherjee, S.C.; Mondal, B.K.; Paul, K.; Lodh, D.; Biswas, B.K.; Chanda, C.R.; Basu, G.K.; Saha, K.C.; et al. Chronic Arsenic Toxicity in Bangladesh and West Bengal, India—A Review and Commentary. J. Toxicol. Clin. Toxicol. 2001, 39, 683–700.
- Mazumder, D.N.G. Chronic Arsenic Toxicity: Clinical Features, Epidemiology, and Treatment: Experience in West Bengal. J. Environ. Sci. Health Part A Tox Hazard. Subst. Environ. Eng. 2003, 38, 141–163.
- Singh, A.; Ghosh, A.K. Groundwater Arsenic Contamination and Its Implications: A Case Study of Shahpur Block of Bhojpur District, Bihar. Int. J. Mod. Eng. Res. 2014, 4, 8.
- Chakraborti, D.; Singh, S.K.; Rahman, M.M.; Dutta, R.N.; Mukherjee, S.C.; Pati, S.; Kar, P.B. Groundwater Arsenic Contamination in the Ganga River Basin: A Future Health Danger. Int. J. Environ. Res. Public Heal. 2018, 15, 180.
- Yang, H.-C.; Rosen, B.P. New mechanisms of bacterial arsenic resistance. Biomed. J. 2016, 39, 5–13.
- Palma-Lara, I.; Martínez-Castillo, M.; Quintana-Pérez, J.; Arellano-Mendoza, M.; Tamay-Cach, F.; Valenzuela-Limón, O.; García-Montalvo, E.; Hernández-Zavala, A. Arsenic exposure: A public health problem leading to several cancers. Regul. Toxicol. Pharmacol. 2020, 110, 104539.
- Wang, W.; Cheng, S.; Zhang, D. Association of inorganic arsenic exposure with liver cancer mortality: A meta-analysis. Environ. Res. 2014, 135, 120–125.
- Cardoso, A.P.F.; Udoh, K.T.; States, J.C. Arsenic-induced changes in miRNA expression in cancer and other diseases. Toxicol. Appl. Pharmacol. 2020, 409, 115306.
- Cohen, S.M.; Chowdhury, A.; Arnold, L.L. Inorganic arsenic: A non-genotoxic carcinogen. J. Environ. Sci. 2016, 49, 28–37.
- Martinez, V.D.; Vucic, E.A.; Becker-Santos, D.D.; Gil, L.; Lam, W.L. Arsenic Exposure and the Induction of Human Cancers. J. Toxicol. 2011, 2011, 1–13.
- Darbandi, M.P.; Taheri, J. Using Sulfur-Containing minerals in medicine: Iranian traditional documents and modern pharmaceutical terminology. Earth Sci. Hist. 2018, 37, 25–33.
- Ommati, M.; Heidari, R.; Manthari, R.K.; Chiranjeevi, S.T.; Niu, R.; Sun, Z.; Sabouri, S.; Zamiri, M.; Zaker, L.; Yuan, J.; et al. Paternal exposure to arsenic resulted in oxidative stress, autophagy, and mitochondrial impairments in the HPG axis of pubertal male offspring. Chemosphere 2019, 236, 124325.
- Leu, L.; Mohassel, L. Arsenic Trioxide as First-Line Treatment for Acute Promyelocytic Leukemia. Am. J. Health-Syst. Pharm. 2009, 66, 1913–1918.
- Ratnaike, R.N. Acute and chronic arsenic toxicity. Postgrad. Med. J. 2003, 79, 391–396.
- Calabrese, E.J.; Baldwin, L.A. Inorganics and Hormesis. Crit. Rev. Toxicol. 2003, 33, 215–304.
- Ommati, M.M.; Manthari, R.K.; Tikka, C.; Niu, R.; Sun, Z.; Sabouri, S.; Zamiri, M.J.; Ahmadi, H.N.; Ghaffari, H.; Heidari, R.; et al. Arsenic-induced autophagic alterations and mitochondrial impairments in HPG-S axis of mature male mice offspring (F1-generation): A persistent toxicity study. Toxicol. Lett. 2020, 326, 83–98.
- Ommati, M.M.; Shi, X.; Li, H.; Zamiri, M.J.; Farshad, O.; Jamshidzadeh, A.; Heidari, R.; Ghaffari, H.; Zaker, L.; Sabouri, S.; et al. The mechanisms of arsenic-induced ovotoxicity, ultrastructural alterations, and autophagic related paths: An enduring developmental study in folliculogenesis of mice. Ecotoxicol. Environ. Saf. 2020, 204, 110973.
- Kumarathilaka, P.; Seneweera, S.; Ok, Y.S.; Meharg, A.; Bundschuh, J. Arsenic in cooked rice foods: Assessing health risks and mitigation options. Environ. Int. 2019, 127, 584–591.
- Taylor, V.; Goodale, B.; Raab, A.; Schwerdtle, T.; Reimer, K.; Conklin, S.; Karagas, M.R.; Francesconi, K.A. Human exposure to organic arsenic species from seafood. Sci. Total. Environ. 2017, 580, 266–282.
- Calatayud, M.; Farias, S.S.; de Paredes, G.S.; Olivera, M.; Carreras, N..; Giménez, M.C.; Devesa, V.; Vélez, D. Arsenic exposure of child populations in Northern Argentina. Sci. Total Environ. 2019, 669, 1–6.
- Kordas, K.; Queirolo, E.I.; Mañay, N.; Peregalli, F.; Hsiao, P.Y.; Lu, Y.; Vahter, M. Low-level arsenic exposure: Nutritional and dietary predictors in first-grade Uruguayan children. Environ. Res. 2016, 147, 16–23.
- Wei, B.; Yu, J.; Kong, C.; Li, H.; Yang, L.; Guo, Z.; Cui, N.; Xia, Y.; Wu, K. An investigation of the health effects caused by exposure to arsenic from drinking water and coal combustion: Arsenic exposure and metabolism. Environ. Sci. Pollut. Res. 2017, 24, 25947–25954.
- Rasheed, H.; Kay, P.; Slack, R.; Gong, Y.Y.; Carter, A. Human exposure assessment of different arsenic species in household water sources in a high risk arsenic area. Sci. Total. Environ. 2017, 584-585, 631–641.
- Chang, J.Y.; Ahn, S.C.; Lee, J.S.; Kim, J.-Y.; Jung, A.-R.; Park, J.; Choi, J.-W.; Yu, S.D. Exposure assessment for the abandoned metal mine area contaminated by arsenic. Environ. Geochem. Health 2019, 41, 2443–2458.
- Signes-Pastor, A.J.; Vioque, J.; Navarrete-Muñoz, E.M.; Carey, M.; García-Villarino, M.; Fernández-Somoano, A.; Tardón, A.; Santa-Marina, L.; Irizar, A.; Casas, M.; et al. Inorganic arsenic exposure and neuropsychological development of children of 4–5 years of age living in Spain. Environ. Res. 2019, 174, 135–142.
- Samal, A.C.; Kar, S.; Bhattacharya, P.; Santra, S. Human exposure to arsenic through foodstuffs cultivated using arsenic contaminated groundwater in areas of West Bengal, India. J. Environ. Sci. Health Part A Tox Hazard. Subst. Environ. Eng. 2011, 46, 1259–1265.
- Menon, M.; Sarkar, B.; Hufton, J.; Reynolds, C.; Reina, S.V.; Young, S. Do arsenic levels in rice pose a health risk to the UK population? Ecotoxicol. Environ. Saf. 2020, 197, 110601.
- Liao, N.; Seto, E.; Eskenazi, B.; Wang, M.; Li, Y.; Hua, J. A Comprehensive Review of Arsenic Exposure and Risk from Rice and a Risk Assessment among a Cohort of Adolescents in Kunming, China. Int. J. Environ. Res. Public Health 2018, 15, 2191.
- Oguri, T.; Yoshinaga, J. Daily Inorganic Arsenic Intake of the Japanese Estimated by a Probabilistic Approach. Nippon Eiseigaku Zasshi Jpn. J. Hyg. 2014, 69, 177–186.
- Rasheed, H.; Kay, P.; Slack, R.; Gong, Y.Y. Arsenic species in wheat, raw and cooked rice: Exposure and associated health implications. Sci. Total. Environ. 2018, 634, 366–373.
- Xue, L.; Zhao, Z.; Zhang, Y.; Liao, J.; Wu, M.; Wang, M.; Sun, J.; Gong, H.; Guo, M.; Li, S.; et al. Dietary exposure to arsenic and human health risks in western Tibet. Sci. Total Environ. 2020, 731, 138840.
- Raeeszadeh, M.; Gravandi, H.; Akbari, A. Determination of some heavy metals levels in the meat of animal species (sheep, beef, turkey, and ostrich) and carcinogenic health risk assessment in Kurdistan province in the west of Iran. Environ. Sci. Pollut. Res. 2022, 1–11.
- Vahter, M. Mechanisms of arsenic biotransformation. Toxicology 2002, 181–182, 211–217.
- National Research Council (US) Subcommittee on Arsenic in Drinking Water. Disposition of Inorganic Arsenic; National Academies Press (US): Washington, DC, USA, 1999.
- Lindgren, A.; Vahter, M.; Dencker, L. Autoradiographic Studies on the Distribution of Arsenic in Mice and Hamsters Administered 74As-Arsenite or -Arsenate. Acta Pharmacol. Toxicol. 1982, 51, 253–265.
- Lindgren, A.; Danielsson, B.R.G.; Dencker, L.; Vahter, M. Embryotoxicity of Arsenite and Arsenate: Distribution in Pregnant Mice and Monkeys and Effects on Embryonic Cells in Vitro. Acta Pharmacol. Toxicol. 1984, 54, 311–320.
- Vahter, M.; Marafante, E.; Lindgren, A.; Dencker, L. Tissue distribution and subcellular binding of arsenic in Marmoset monkeys after injection of 74As-Arsenite. Arch. Toxicol. 1982, 51, 65–77.
- Garbinski, L.D.; Rosen, B.P.; Chen, J. Pathways of arsenic uptake and efflux. Environ. Int. 2019, 126, 585–597.
- Roggenbeck, B.A.; Banerjee, M.; Leslie, E.M. Cellular arsenic transport pathways in mammals. J. Environ. Sci. 2016, 49, 38–58.
- Hughes, M.F. Arsenic toxicity and potential mechanisms of action. Toxicol. Lett. 2002, 133, 1–16.
- Rehman, K.; Naranmandura, H. Arsenic metabolism and thioarsenicals. Metallomics 2012, 4, 881–892.
- Suzuki, K.; Ogra, Y. Distributions and chemical forms of arsenic after intravenous administration of dimethylarsinic and monomethylarsonic acids to rats. Toxicol. Appl. Pharmacol. 2004, 198, 336–344.
- Kligerman, A.D.; Doerr, C.L.; Tennant, A.H.; Harrington-Brock, K.; Allen, J.W.; Winkfield, E.; Poorman-Allen, P.; Kundu, B.; Funasaka, K.; Roop, B.C.; et al. Methylated trivalent arsenicals as candidate ultimate genotoxic forms of arsenic: Induction of chromosomal mutations but not gene mutations. Environ. Mol. Mutagen. 2003, 42, 192–205.
- Mass, M.J.; Tennant, A.; Roop, B.C.; Cullen, W.R.; Styblo, M.; Thomas, D.J.; Kligerman, A.D. Methylated Trivalent Arsenic Species Are Genotoxic. Chem. Res. Toxicol. 2001, 14, 355–361.
- Braman, R.S.; Foreback, C.C. Methylated Forms of Arsenic in the Environment. Science 1973, 182, 1247–1249.
- Chouchane, S.; Snow, E.T. In Vitro Effect of Arsenical Compounds on Glutathione-Related Enzymes. Chem. Res. Toxicol. 2001, 14, 517–522.
- Yamanaka, K.; Hayashi, H.; Tachikawa, M.; Kato, K.; Hasegawa, A.; Oku, N.; Okada, S. Metabolic methylation is a possible genotoxicity-enhancing process of inorganic arsenics. Mutat. Res. Toxicol. Environ. Mutagen. 1997, 394, 95–101.
- Ahmad, S.; Kitchin, K.T.; Cullen, W.R. Arsenic Species That Cause Release of Iron from Ferritin and Generation of Activated Oxygen. Arch. Biochem. Biophys. 2000, 382, 195–202.
- Corsini, E.; Asti, L.; Viviani, B.; Marinovich, M.; Galli, C.L. Sodium Arsenate Induces Overproduction of Interleukin-1α in Murine Keratinocytes: Role of Mitochondria. J. Investig. Dermatol. 1999, 113, 760–765.
- Zargari, F.; Rahaman, S.; KazemPour, R.; Hajirostamlou, M. Arsenic, Oxidative Stress and Reproductive System. J. Xenobiotics 2022, 12, 214–222.
- Adewoyin, M.; Ibrahim, M.; Roszaman, R.; Isa, M.L.M.; Alewi, N.A.M.; Rafa, A.A.A.; Anuar, M.N.N. Male Infertility: The Effect of Natural Antioxidants and Phytocompounds on Seminal Oxidative Stress. Diseases 2017, 5, 9.
- Khair, A.; Awal, M.A.; Hoque, M.N.; Talukder, A.K.; Das, Z.C.; Rao, D.R.; Shamsuddin, M. Spirulina ameliorates arsenic induced reproductive toxicity in male rats. Anim. Reprod. 2021, 18.
- Tsai, C.-H.; Yang, M.-H.; Hung, A.C.; Wu, S.-C.; Chiu, W.-C.; Hou, M.-F.; Tyan, Y.-C.; Wang, Y.-M.; Yuan, S.-S.F. Identification of Id1 as a downstream effector for arsenic-promoted angiogenesis via PI3K/Akt, NF-κB and NOS signaling. Toxicol. Res. 2015, 5, 151–159.
- Huang, Q.; Luo, L.; Alamdar, A.; Zhang, J.; Liu, L.; Tian, M.; Eqani, S.A.M.A.S.; Shen, H. Integrated proteomics and metabolomics analysis of rat testis: Mechanism of arsenic-induced male reproductive toxicity. Sci. Rep. 2016, 6, 32518.
- Huang, Y.-C.; Yu, H.-S.; Chai, C.-Y. Proteins in the ERK pathway are affected by arsenic-treated cells. Toxicol. Res. 2015, 4, 1545–1554.
- Chen, J.; Fok, K.L.; Chen, H.; Zhang, X.H.; Xu, W.M.; Chan, H.C. Cryptorchidism-induced CFTR down-regulation results in disruption of testicular tight junctions through up-regulation of NF- B/COX-2/PGE2. Hum. Reprod. 2012, 27, 2585–2597.
- Xia, Z.-P.; Zheng, X.-M.; Zheng, H.; Liu, X.-J.; Liu, G.-Y.; Wang, X. Downregulation of cold-inducible RNA-binding protein activates mitogen-activated protein kinases and impairs spermatogenic function in mouse testes. Asian J. Androl. 2012, 14, 884–889.
- Danielsson, B.R.G.; Dencker, L.; Lindgren, A.; Tjälve, H. Accumulation of Toxic Metals in Male Reproduction Organs. Arch. Toxicol. Suppl. 1984, 7, 177–180.
- Pant, N.; Kumar, R.; Murthy, R.C.; Srivastava, S.P. Male reproductive effect of arsenic in mice. BioMetals 2001, 14, 113–117.
- Zubair, M.; Ahmad, M.; Qureshi, Z.I. Review on Arsenic-Induced Toxicity in Male Reproductive System and Its Amelioration. Andrologia 2017, 49, e12791.
- Kumar, A.; Raj, V.; Srivastava, A.; Ali, M.; Ghosh, A.K.; Rachamalla, M.; Kumar, D. Autophagy in arsenic exposed population and cancer patients. In Autophagy and Metabolism; Academic Press: Cambridge, MA, USA, 2022; pp. 141–161.
- Roy, S.; Bhattacharya, S. Arsenic-induced histopathology and synthesis of stress proteins in liver and kidney of Channa punctatus. Ecotoxicol. Environ. Saf. 2006, 65, 218–229.
- Ahmed, K.; Mamun, H.A.; Parvin, E.; Akter, M.S.; Khan, M.S. Arsenic induced toxicity and histopathological changes in gill and liver tissue of freshwater fish, tilapia (Oreochromis mossambicus). Exp. Toxicol. Pathol. 2013, 65, 903–909.
- Manthari, R.K.; Tikka, C.; Ommati, M.M.; Niu, R.; Sun, Z.; Wang, J.; Zhang, J.; Wang, J. Arsenic-Induced Autophagy in the Developing Mouse Cerebellum: Involvement of the Blood-Brain Barrier’s Tight-Junction Proteins and the PI3K-Akt-MTOR Signaling Pathway. J. Agric. Food Chem. 2018, 66, 8602–8614.
- Manthari, R.K.; Tikka, C.; Ommati, M.M.; Niu, R.; Sun, Z.; Wang, J.; Zhang, J.; Wang, J. Arsenic Induces Autophagy in De-velopmental Mouse Cerebral Cortex and Hippocampus by Inhibiting PI3K/Akt/MTOR Signaling Pathway: Involvement of Blood–Brain Barrier’s Tight Junction Proteins. Arch. Toxicol. 2018, 92, 3255–3275.
- Dastgiri, S.; Mosaferi, M.; Fizi, M.A.H.; Olfati, N.; Zolali, S.; Pouladi, N.; Azarfam, P. Arsenic Exposure, Dermatological Lesions, Hypertension, and Chromosomal Abnormalities among People in a Rural Community of Northwest Iran. J. Health Popul. Nutr. 2010, 28, 14.
- Ommati, M.M.; Heidari, R.; Zamiri, M.J.; Sabouri, S.; Zaker, L.; Farshad, O.; Jamshidzadeh, A.; Mousapour, S. The Footprints of Oxidative Stress and Mitochondrial Impairment in Arsenic Trioxide-Induced Testosterone Release Suppression in Pubertal and Mature F1-Male Balb/c Mice via the Downregulation of 3β-HSD, 17β-HSD, and CYP11a Expression. Biol. Trace Elem. Res. 2020, 195, 125–134.
- Acevedo-Rodriguez, A.; Kauffman, A.S.; Cherrington, B.D.; Borges, C.S.; Roepke, T.A.; Laconi, M. Emerging Insights into Hypothalamic-Pituitary-Gonadal (HPG) Axis Regulation and Interaction with Stress Signaling. J. Neuroendocrinol. 2018, 30, e12590.
- Chiaverini, N.; de Ley, M. Protective Effect of Metallothionein on Oxidative Stress-Induced DNA Damage. Free Radic. Res. 2010, 44, 605–613.
- Siu, E.; Mruk, D.D.; Porto, C.S.; Cheng, C.Y. Cadmium-induced testicular injury. Toxicol. Appl. Pharmacol. 2009, 238, 240–249.
- Dalton, T.; Fu, K.; Enders, G.C.; Palmiter, R.D.; Andrews, G.K. Analysis of the effects of overexpression of metallothionein-I in transgenic mice on the reproductive toxicology of cadmium. Environ. Health Perspect. 1996, 104, 68–76.
- Cheng, C.Y.; Mruk, D.D. The Blood-Testis Barrier and Its Implications for Male Contraception. Pharmacol. Rev. 2011, 64, 16–64.
- Sanghamitra, S.; Hazra, J.; Upadhyay, S.N.; Singh, R.K.; Amal, R.C. Arsenic induced toxicity on testicular tissue of mice. Indian J. Physiol. Pharmacol. 2008, 52, 84–90.
- Wang, L.; Hao, J.; Hu, J.; Pu, J.; Lü, Z.; Zhao, L.; Wang, Q.; Yu, Q.; Wang, Y.; Li, G. Protective Effects of Ginsenosides against Bisphenol A-Induced Cytotoxicity in 15P-1 Sertoli Cells via Extracellular Signal-Regulated Kinase 1/2 Signalling and Antioxidant Mechanisms. Basic Clin. Pharmacol. Toxicol. 2012, 111, 42–49.
- Momeni, H.R.; Oryan, S.; Eskandari, N. Effect of vitamin E on sperm number and testis histopathology of sodium arsenite-treated rats. Reprod. Biol. 2012, 12, 171–181.
- de Medeiros, P.; Samelo, R.R.; Silva, A.P.G.; Santiago, M.; Duarte, F.; Castro, .; Perobelli, J.E. Prepubertal exposure to low doses of sodium arsenite impairs spermatogenesis and epididymal histophysiology in rats. Environ. Toxicol. 2018, 34, 83–91.
- Anwar, N.; Qureshi, I.Z.; Spears, N.; Lopes, F. In vitro administration of sodium arsenite in mouse prepubertal testis induces germ cell loss and apoptosis. Toxicol. Vitro 2020, 67, 104924.
- Souza, A.C.F.; Marchesi, S.C.; Lima, G.D.D.A.; Ferraz, R.P.; Santos, F.C.; da Matta, S.L.P.; Machado-Neves, M. Effects of Sodium Arsenite and Arsenate in Testicular Histomorphometry and Antioxidants Enzymes Activities in Rats. Biol. Trace Element Res. 2015, 171, 354–362.
- Chiou, T.-J.; Chu, S.-T.; Tzeng, W.-F.; Huang, Y.-C.; Liao, C.-J. Arsenic Trioxide Impairs Spermatogenesis via Reducing Gene Expression Levels in Testosterone Synthesis Pathway. Chem. Res. Toxicol. 2008, 21, 1562–1569.
- Wang, Y.-X.; Wang, P.; Feng, W.; Liu, C.; Yang, P.; Chen, Y.-J.; Sun, L.; Sun, Y.; Yue, J.; Gu, L.-J.; et al. Relationships between seminal plasma metals/metalloids and semen quality, sperm apoptosis and DNA integrity. Environ. Pollut. 2017, 224, 224–234.
- Lima, G.D.D.A.; Sertorio, M.N.; Souza, A.C.F.; Menezes, T.P.; Mouro, V.G.S.; Gonçalves, N.M.; de Oliveira, J.M.; Henry, M.; Machado-Neves, M. Fertility in male rats: Disentangling adverse effects of arsenic compounds. Reprod. Toxicol. 2018, 78, 130–140.
- da Silva, R.F.; Borges, C.D.S.; Lamas, C.D.A.; Cagnon, V.H.A.; Kempinas, W.D.G. Arsenic trioxide exposure impairs testicular morphology in adult male mice and consequent fetus viability. J. Toxicol. Environ. Health Part A Curr. Issues 2017, 80, 1166–1179.
- Han, Y.; Liang, C.; Yu, Y.; Manthari, R.K.; Cheng, C.; Tan, Y.; Li, X.; Tian, X.; Fu, W.; Yang, J.; et al. Chronic arsenic exposure lowered sperm motility via impairing ultra-microstructure and key proteins expressions of sperm acrosome and flagellum formation during spermiogenesis in male mice. Sci. Total Environ. 2020, 734, 139233.
- Han, Y.; Liang, C.; Manthari, R.K.; Yu, Y.; Gao, Y.; Liu, Y.; Jiang, S.; Tikka, C.; Wang, J.; Zhang, J. Arsenic influences spermatogenesis by disorganizing the elongation of spermatids in adult male mice. Chemosphere 2019, 238, 124650.
- Kesari, V.P.; Kumar, A.; Khan, P.K. Induction of sperm impairments in mice as a sensitive biomarker of arsenic toxicity. Environ. Monit. Assess. 2014, 186, 3115–3121.
- Souza, A.C.F.; Bastos, D.S.S.; Sertorio, M.N.; Santos, F.C.; Ervilha, L.O.G.; de Oliveira, L.L.; Machado-Neves, M. Combined effects of arsenic exposure and diabetes on male reproductive functions. Andrology 2019, 7, 730–740.
- Guvvala, P.R.; Sellappan, S.; Parameswaraiah, R.J. Impact of arsenic(V) on testicular oxidative stress and sperm functional attributes in Swiss albino mice. Environ. Sci. Pollut. Res. 2016, 23, 18200–18210.
- Rodriguez, K.F.; Mellouk, N.; Ungewitter, E.K.; Nicol, B.; Liu, C.; Brown, P.R.; Willson, C.J.; Yao, H.H.-C. In utero exposure to arsenite contributes to metabolic and reproductive dysfunction in male offspring of CD-1 mice. Reprod. Toxicol. 2020, 95, 95–103.
- Nava-Rivera, L.E.; Betancourt-Martínez, N.D.; Lozoya-Martínez, R.; Carranza-Rosales, P.; Guzmán-Delgado, N.E.; Carranza-Torres, I.E.; Delgado-Aguirre, H.; Zambrano-Ortíz, J.O.; Morán-Martínez, J. Transgenerational effects in DNA methylation, genotoxicity and reproductive phenotype by chronic arsenic exposure. Sci. Rep. 2021, 11, 1–16.
- Souza, A.C.F.; Machado-Neves, M.; Bastos, D.S.S.; Santos, F.C.; Ervilha, L.O.G.; Coimbra, J.L.D.P.; Araújo, L.D.S.; de Oliveira, L.L.; Guimarães, S.E.F. Impact of prenatal arsenic exposure on the testes and epididymides of prepubertal rats. Chem. Interact. 2020, 333, 109314.
- Couto-Santos, F.; Viana, A.G.D.A.; Souza, A.C.F.; Dutra, A.A.D.A.; Mendes, T.A.D.O.; Ferreira, A.T.D.S.; Aguilar, J.E.P.; Oliveira, L.L.; Machado-Neves, M. Prepubertal arsenic exposure alters phosphoproteins profile, quality, and fertility of epididymal spermatozoa in sexually mature rats. Toxicology 2021, 460, 152886.
- Souza, A.C.F.; Marchesi, S.C.; Ferraz, R.P.; Lima, G.D.D.A.; de Oliveira, J.A.; Machado-Neves, M. Effects of sodium arsenate and arsenite on male reproductive functions in Wistar rats. J. Toxicol. Environ. Health Part A 2016, 79, 274–286.
- Ram, A.K.S.S.; Reddy, K.P.; Girish, B.P.; Supriya, C.; Reddy, P.S. Arsenic aggravated reproductive toxicity in male rats exposed to lead during the perinatal period. Toxicol. Res. 2018, 7, 1191–1204.
- Liu, P.; Li, R.; Tian, X.; Zhao, Y.; Li, M.; Wang, M.; Ying, X.; Yuan, J.; Xie, J.; Yan, X.; et al. Co-exposure to fluoride and arsenic disrupts intestinal flora balance and induces testicular autophagy in offspring rats. Ecotoxicol. Environ. Saf. 2021, 222, 112506.
More
Information
Subjects:
Reproductive Biology
Contributor
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.0K
Revisions:
2 times
(View History)
Update Date:
13 Dec 2022
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No