Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Cui, Z.; Jiao, Y.; Pu, L.; Tang, J.Z.; Wang, G. Skeletal Muscle Gene Delivery. Encyclopedia. Available online: https://encyclopedia.pub/entry/36399 (accessed on 24 July 2026).
Cui Z, Jiao Y, Pu L, Tang JZ, Wang G. Skeletal Muscle Gene Delivery. Encyclopedia. Available at: https://encyclopedia.pub/entry/36399. Accessed July 24, 2026.
Cui, Zhanpeng, Yang Jiao, Linyu Pu, James Zhenggui Tang, Gang Wang. "Skeletal Muscle Gene Delivery" Encyclopedia, https://encyclopedia.pub/entry/36399 (accessed July 24, 2026).
Cui, Z., Jiao, Y., Pu, L., Tang, J.Z., & Wang, G. (2022, November 24). Skeletal Muscle Gene Delivery. In Encyclopedia. https://encyclopedia.pub/entry/36399
Cui, Zhanpeng, et al. "Skeletal Muscle Gene Delivery." Encyclopedia. Web. 24 November, 2022.
Copy Citation
Since Jon A. Wolff found skeletal muscle cells being able to express foreign genes and Russell J. Mumper increased the gene transfection efficiency into the myocytes by adding polymers, skeletal muscles have become a potential gene delivery and expression target. Different methods have been developing to deliver transgene into skeletal muscles. Among them, viral vectors may achieve potent gene delivery efficiency. Therefore, non-viral biomaterial-mediated methods with reliable biocompatibility are promising tools for intramuscular gene delivery in situ. A series of advanced non-viral gene delivery materials and related methods have been reported, such as polymers, liposomes, cell penetrating peptides, as well as physical delivery methods.
non-viral materials
gene delivery
skeletal muscle
1. Introduction
Due to the wide distribution and enormous number of skeletal muscles in the human body, it has high application value to deliver functional genes into skeletal muscle cells for target protein expression. Skeletal muscles can act as protein factories, wherein exogenous functional genes can serve as “blueprints” to produce target proteins for specific purposes, such as antibodies for cancer immunotherapy[1] and insulin analogues for diabetes treatment[2]. Delivery of exogenous genes into cells needs vectors, which are mainly divided into viral vectors and non-viral vectors. Although viral vectors had high gene delivery efficiency, their potential biosafety risks limited the clinical applications[3]. In 1990, Jon A. Wolff[4] first reported that skeletal muscles could take up plasmid DNA (pDNA) and express the reporter protein. In the experiments, naked pDNA carrying LacZ gene was directly injected into a quadriceps muscle of mice and seven days later, the whole muscle was removed and stained by X-Gal. The stained cross-section of the muscle tissue showed that the target gene could be expressed in muscle cells to a certain extent. However, it is difficult to acquire high gene delivery efficiency and expression level via direct injection of naked pDNA into skeletal muscles.
Transfection of pDNA into cultured cells is a process in which cells actively or passively acquire foreign DNA in a simple in vitro environment. The main barriers of gene transfection include cell plasma membrane, escape of genes from endosomes and lysosomes, detachment of genes from gene carriers, nuclease, and cell nuclear membrane, etc. [5][6]. In comparison, delivery of pDNA into skeletal muscle cells is carried out in a more complex in vivo environment, so it is confronted with extracellular and intracellular obstacles. Before penetrating the cell plasma membrane, it needs to overcome the extracellular obstacles first. Owing to the large size and negative charge of pDNA molecules, naked pDNAs can be easily trapped or damaged by extracellular obstacles, such as extracellular matrix (ECM), biomacromolecules with positive charge as well as nuclease in the ECM [5](Scheme 1). To solve this problem, Russell et al. combined polyvinyl pyrrolidone (PVP, Table 1) and polyvinyl alcohol (PVA) with pDNA in mixed solution and injected the solution into the tibialis anterior (TA) muscles, generating improved expression level of exogenous genes compared to naked pDNA injection [7]. The study showed that gene delivery efficiency into skeletal muscles could be improved by some non-viral biomaterials.
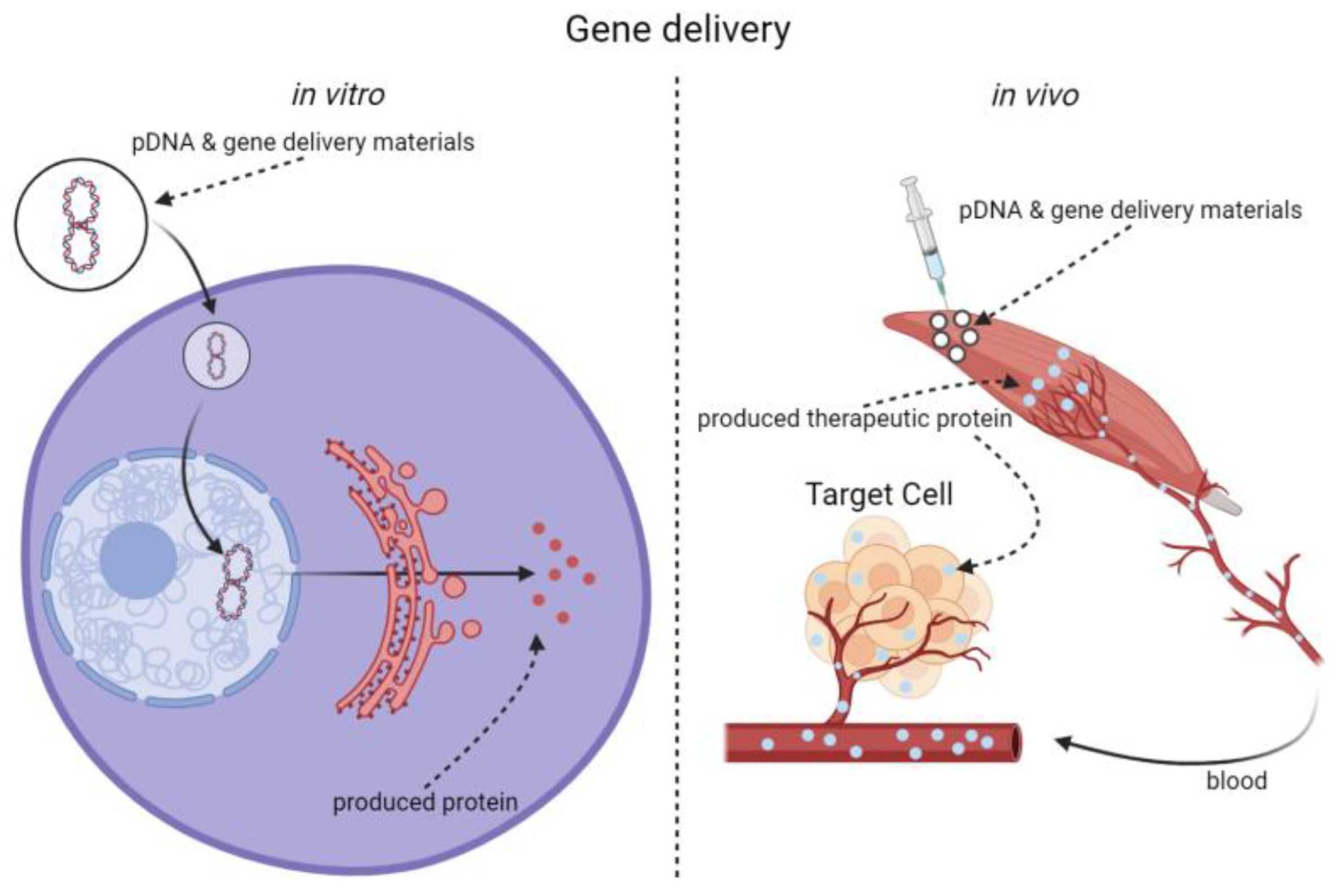
Scheme 1. Differences between gene delivery in vivo and transfection in vitro.
Table 1. Polymers for gene delivery to skeletal muscles.
| Polymers | Year | Reporter Genes | Functional Genes | Function |
|---|---|---|---|---|
| Polyvinyl pyrrolidone (PVP) | 1996 [7] | CAT, β-CAT | / |
|
| Pluronic P85® | 2005 [8] | Luc | / |
|
| 2009 [9] | GFP, Luc | / |
|
|
| 2016 [10] | Luc, GFP | pDRIVE5Lucia-mDesmin & pDRIVE5GFP-mDesmin |
|
|
| Multi-block copolymers from Pluronic P85 and di-(ethylene glycol) divinyl ether® | 2009 [11] | Luc | / |
|
| Pluronic SP1017® | 2005 [8] | Luc | / |
|
| Pluronic 25R2® | 2011 [12] | Luc | / |
|
| Pluronic L64® | 2002 [13] | Luc, β-Gal | / |
|
| 2014 [14] | / | HIF-1α |
|
|
| 2019 [15] | lacZ, Luc, E2-Crimson | GHRH |
|
|
| 2021 [1] | E2-Crimson, EGFP | mlFVII Icon |
|
|
| PEG13-PLGA10-PEG13 | 2007 [16] | Luc | VEGF |
|
| PEO–PTHF–PEO | 2008 [17] | Luc, lacZ | / |
|
| LPL | 2014 [18] | lacZ, Luc, E2-Crimson | mGH |
|
| rL2PL2, rL3PL3 | 2016 [19] | lacZ, Luc, E2-Crimson | GHRH |
|
| TBCPs | 2020 [20] | Luc | / |
|
| Lutrol | 2005 [21] | Luc | / |
|
| Poloxamine 304 | ||||
| poly(D, L-lactide-co-glycolide) | 2006 [22] | Luc | / |
|
| PLGA/E100 | 2009 [23] | / | IL-10 |
|
| HCPEI | 2008 [24] | / | VEGF |
|
| TAEI-PEI | 2012 [25] | GFP | / |
|
| PLGA | 2008 [26] | / | VEGF |
|
| PEG-PAMAM | 2009 [3] | EGFP | / |
|
| PAMAM-G5/SMTP/DBP/NLS mixture | 2021 [27] | EGFP | micro-dystrophin (µDys) |
|
In contrast to the prosperity of non-viral vectors regarding in vitro gene transfection of cells and in vivo targeted gene delivery through the bloodstream, there were limited studies on non-viral vectors and related methods for skeletal muscle gene delivery [28].
2. Advantages and Challenges of the Skeletal Muscle Gene Delivery
For the traditional gene therapy, the therapeutic genes were usually sent to the lesion cells, such as in cancer treatment[29][30][31]. However, for the skeletal muscle-based gene therapy, the delivery targets were skeletal muscle cells. Previous studies have screened the muscle-specific promoter [32], proving that skeletal muscle has microbubbles, which can bring the protein secreted out of the cells, affect the adjacent cells, and regulate the behaviour of them [33]. In addition, proteins secreted by muscle cells can also enter the circulatory system and affect physiological parameters, such as hormone secretion, and nervous and immune system activity [34]. Based on the above points, some scientists have successfully introduced the vector carrying insulin gene into skeletal muscle with genetic engineering technology and used it to treat Type 1 diabetes [35]. After that, another study showed that the co-expression of insulin and glucokinase could be applied to correct hyperglycaemia and prevent hypoglycaemia [36]. It can be concluded that in these systems, skeletal muscle cells played the role as protein factories, rather than a lesion site.
2.1. Advantages of the Skeletal Muscle as the Target for Gene Delivery
Skeletal muscles have many inherent anatomical, cellular, and physiological characteristics. It is a good target tissue for gene expression, especially for the production of proteins such as systemic therapeutic agents. The significant advantages of skeletal muscle as a target tissue for gene delivery can be mainly attributed to four points:
(1) The weight of skeletal muscles accounts for 30% of a normal adult’s weight, which means there is enough target tissue for gene delivery; (2) Skeletal muscles have rich vascular systems. Abundant capillary networks wrap each muscle fiber at regular intervals, thus providing an effective transportation system for secreted proteins to enter the circulation; (3) Skeletal muscle fibers contain terminally differentiated cells, and the nuclei in the fibers are postmitotic. A single muscle fiber can survive in the living body for a long time. Even if muscle fibers are damaged and only a short segment of some fibers is unimpaired, the nucleus of surviving muscle can still work [37]. It provides a stable environment for the continuous production of proteins; (4) In muscle fibers, foreign genes can be spread from a limited injection site to the nuclei of a large number of adjacent muscle cells in the fiber. This ability to diffuse foreign genes in muscle fibers is probably one of the reasons why foreign genes can be highly expressed in muscles [28].
Therefore, skeletal muscles are expected to become a potential target tissue for gene therapy. Gene delivery to skeletal muscle is simple as it only requires intramuscular injection, differing from long-term daily insulin injection or radiotherapy and chemotherapy.
2.2. Obstacles to Skeletal Muscles Gene Delivery
Systemic administration of genes transports foreign vectors through the circulatory system, which may face obstacles of off-target effects on other organs and stability changes in serum. As a local injection method, gene delivery to skeletal muscle cells allows direct injection of prepared plasmid DNA. Therefore, the obstacles mainly exist in the ECM, cytoplasmic membrane, endosome escape, and nuclear entry.
2.2.1. Obstructions in the Extracellular Matrix
Unlike other ECMs, the ECM of the skeletal muscle is a three-dimensional scaffold composed of various collagens, glycoproteins, proteoglycans, and elastin[38] , and many of these proteins are negatively charged [39].
Generally, the delivery efficiency of most gene carriers, such as liposomes and polymers, is much lower than that of viral vectors. Unlike other organs, muscle fibers are surrounded by a mechanically strong ECM, which is the glycosaminoglycan-rich basement membrane in the skeletal muscle [40][41][42] and the ECM is strongly negatively charged. As a result, cationic liposomes and polymers are very easily bound to the negatively charged biomacromolecules in the ECM, which hinders the entry of cationic DNA/carriers complexes into cells [43].
Naked plasmids have shown to be useful for gene transfer into skeletal muscles[4], but low transfer efficiency brought great challenges to clinical promotion. Studies have shown that adding nuclease inhibitors could improve the transfer efficiency of pDNA into muscles [44], which proved that nuclease was also one of the main factors in ECM that inhibited pDNA transfer efficiency.
2.2.2. Cytoplasmic Membrane
The existence of the cell membrane provides a relatively stable environment for the cell. The main structure of the cytoplasmic membrane is a phospholipid bilayer containing phospholipid molecules, cholesterol, and proteins embedded in the membrane or on the membrane surface. Highly hydrophilic and bulky pDNA molecules are easily blocked due to the existence of the amphiphilic bilayers in the cytoplasmic membrane. Therefore, how the gene and the carrier cooperate to cross the cytoplasmic membrane and enter the cell is also a key issue in gene delivery. So far, the methods used to improve the delivery efficiency of pDNA mainly included adding components that could promote the internalization of DNA into target cells, such as transferrin[45] , organic solvents, nonionic polymers or surfactants, etc. [5]. According to the properties of these substances, these additives were speculated to temporarily change the permeability of the cytoplasmic membrane by disturbing the cytoplasmic membrane, and therefore allowing pDNA to penetrate the membrane more easily [46]. In addition to adding the above substances, some physical methods could temporarily create non-lethal pores in the cell membrane to facilitate the passage of pDNA, such as gene gun[47] , electroporation[48] , ultrasonic microbubble[49] , and hydrodynamic[50] methods.
2.2.3. Endosomal Escaping
For osmotic and invasive delivery methods, genes generally enter the cytoplasm directly through the cytoplasmic membrane and do not involve endosomal escape. The delivery method of genes into cells by endocytosis needs to consider the problem of endosomal escape. After genes undergoing endocytosis, endosomes/lysosomes may form, and lysosomes will decompose foreign substances into small molecules for reuse, which will make the therapeutic gene ineffective. Therefore, target genes should escape from endosomes after endocytosis to avoid being degraded.
Generally, cationic polymers mainly escape from endosomes to mediate gene delivery, which is represented by branched polyethyleneimine (bPEI, 25 kb). The bPEI may form PEI-DNA complexes first, enter cells through endocytosis, then escape from the endosome into the cytoplasm through the “proton sponge” effect [51].
2.2.4. Entering the Nucleus
For viral vectors, the virus can accomplish the processes of membrane entry, endosomal escape, and entry into the nucleus only by its shell. The delivery efficiency of non-viral gene delivery methods is generally low. The reason may be that the functionality of non-viral gene delivery materials is not complete. In general, numerous studies of non-viral delivery methods have focused on cytoplasmic membrane penetration and endosomal escape. However, in the whole process of gene delivery, whether the exogenous gene can enter the nucleus for expression stably is one of the key factors affecting the efficiency of gene expression. The nucleus is the centre of the cell surrounded by two membranes, and few small particles can freely pass in and out through the nuclear pores [6].
For dividing cells, gene delivery becomes relatively easy, because during cell division the nuclear envelope breaks, making it easier for foreign genes to enter the nucleus [6]. Studies have shown that low molecular weight bPEI/DNA complexes enter the nucleus more easily than the high molecular weight bPEI/DNA complexes [52]. But for cells without division, gene delivery becomes relatively complicated and difficult, and the nuclear membrane blocks larger-sized molecules, which is one of the major challenges of gene delivery.
For this problem, nuclear localization signal (NLS) peptides [53] were used, which can help larger particles to complete the nuclear entry by embedding nuclear pores in the nuclear envelope. Subsequent studies have shown that binding to NLS polypeptides could improve gene entry and expression in non-dividing cells [6][54].
2.2.5. Material Stability
Skeletal muscle gene delivery requires sufficient material stability in the ECM, and then easily to be degraded or cleared in cells. For cationic gene delivery materials, it is easy to form cationic material/DNA complex, but the cationic complex can electrostatically interact with negatively charged biomacromolecules in the ECM, leading to significantly reduced gene delivery efficiency. Meanwhile, the strong binding of material molecules to DNA does not necessarily improve the transfection efficiency, since it may hinder the unpacking of the cationic material and DNA [55]. For branched PEI, the representative of cationic polymers, high molecular weight polymers can bind to DNA better to protect DNA from degradation, and are more readily taken up by cells, resulting in higher delivery efficiency, but low molecular weight polymers have lower cytotoxicity and better DNA unpacking ability [56][57]. Therefore, it can be speculated that there should be an optimal molecular weight range of polymers, so that these polymers have suitable DNA compression and unpacking ability with better biosafety and gene delivery efficiency as well.
2.2.6. Biosecurity
Although there are many methods with high gene delivery efficiency, there are also biological safety issues, such as cytotoxicity, inflammatory response, and immune reaction. For in vivo gene delivery, biosafety is an unavoidable issue. Although viral vectors have high gene delivery efficiency and expression level, they can easily trigger excessive inflammatory responses [58] or immune reaction, while non-viral delivery methods avoid the occurrence of side effects to a great extent. For cationic polymers, the toxicity mainly comes from excessive positive charges. At present, polyethylene glycol (PEG) has been used to shield the cations on the surface of the complex to reduce cytotoxicity [59]. Grandinetti et al. showed that the polymer could directly participate in the nuclear localization of DNA through its membrane destruction characteristics, which may also be the reason for its cytotoxicity [60]. Therefore, it seems to be a better choice to use neutral block polymers since they are more effective than cationic polymers in skeletal muscle gene delivery [14][18][19][61].
References
- Ma, L.; Wang, G.; Liu, S.; Bi, F.; Liu, M.; Wang, G.; Intramuscular Expression of Plasmid-Encoded FVII-Fc Immunoconjugate for Tumor Immunotherapy by Targeting Tumoral Blood Vessels and Cells. Front. Oncol. 2021, 11, 638591.
- Deng, L.; Yang, P.; Li, C.; Xie, L.; Lu, W.; Zhang, Y.; Liu, M.; Wang, G.; Prolonged control of insulin-dependent diabetes via intramuscular expression of plasmid-encoded single-strand insulin analogue. . Genes Dis. 2022, /, /, 10.1016/j.gendis.2022.05.009.
- Qi, R.; Gao, Y.; Tang, Y.; He, R.R.; Liu, T.L.; He, Y.; Sun, S.; Li, B.Y.; Li, Y.B.; Liu, G.; et al. PEG-conjugated PAMAM dendrimers mediate efficient intramuscular gene expression. Aaps j 2009, 11, 395-405, 10.1208/s12248-009-9116-1.
- Wolff, J.A.; Malone, R.W.; Williams, P.; Chong, W.; Acsadi, G.; Jani, A.; Felgner, P.L.; Direct gene transfer into mouse muscle in vivo. Science 1990, 247, 1465-1468.
- Gao, X.; Kim, K.S.; Liu, D.; Nonviral gene delivery: what we know and what is next.. Aaps j 2007, 9, E92-104, 10.1208/aapsj0901009..
- Aied, A.; Greiser, U.; Pandit, A.; Wang, W.; Polymer gene delivery: overcoming the obstacles.. Drug Discov. Today 2013, 18, 1090-1098, 10.1016/j.drudis.2013.06.014.
- Mumper, R.J.; Duguid, J.G.; Anwer, K.; Barron, M.K.; Nitta, H.; Rolland, A.P.; Polyvinyl derivatives as novel interactive polymers for controlled gene delivery to muscle.. Pharm Res 1996, 13, 701-709, 10.1023/a:1016039330870.
- Yang, Z.; Zhu, J.; Sriadibhatla, S.; Gebhart, C.; Alakhov, V.; Kabanov, A.; Yang, Z.; Zhu, J.; Sriadibhatla, S.; Gebhart, C.; Alakhov, V.; Kabanov, A.. J Control Release 2005, 108, 496-512.
- Gaymalov, Z.Z.; Yang, Z.; Pisarev, V.M.; Alakhov, V.Y.; Kabanov, A.V.; The effect of the nonionic block copolymer pluronic P85 on gene expression in mouse muscle and antigen-presenting cells.. Biomaterials 2009, 30, 1232-1245.
- Mahajan, V.; Gaymalov, Z.; Alakhova, D.; Gupta, R.; Zucker, I.H.; Kabanov, A.V.; Horizontal gene transfer from macrophages to ischemic muscles upon delivery of naked DNA with Pluronic block copolymers.. Biomaterials 2016, 75, 58-70.
- Namgung, R.; Nam, S.; Kim, S.K.; Son, S.; Singha, K.; Kwon, J.S.; Ahn, Y.; Jeong, M.H.; Park, I.K.; Garripelli, V.K.; et al.et al. An acid-labile temperature-responsive sol-gel reversible polymer for enhanced gene delivery to the myocardium and skeletal muscle cells. Biomaterials 2009, 30, 5225-5233, 10.1016/j.biomaterials.2009.05.073..
- Guiraud, S.; Alimi-Guez, D.; van Wittenberghe, L.; Scherman, D.; Kichler, A.; The reverse block copolymer Pluronic 25R2 promotes DNA transfection of skeletal muscle. Macromol Biosci 2011, 11, 590-594, 10.1002/mabi.201000463.
- Pitard, B.; Pollard, H.; Agbulut, O.; Lambert, O.; Vilquin, J.T.; Cherel, Y.; Abadie, J.; Samuel, J.L.; Rigaud, J.L.; Menoret, S.; et al.et al. A nonionic amphiphile agent promotes gene delivery in vivo to skeletal and cardiac muscles. Hum Gene Ther 2002, 13, 1767-1775, 10.1089/104303402760293592.
- Song, H.; Liu, S.; Li, C.; Geng, Y.; Wang, G.; Gu, Z.; Pluronic L64-mediated stable HIF-1α expression in muscle for therapeutic angiogenesis in mouse hindlimb ischemia. Int J Nanomedicine 2014, 9, 3439-3452, 10.2147/ijn.S65353.
- He, Y.; Liu, Y.; Sun, Z.; Han, F.; Tang, J.Z.; Gao, R.; Wang, G.; The proper strategy to compress and protect plasmid DNA in the Pluronic L64-electropulse system for enhanced intramuscular gene delivery. Regen Biomater 2019, 6, 289-298, 10.1093/rb/rby028.
- Chang, C.W.; Choi, D.; Kim, W.J.; Yockman, J.W.; Christensen, L.V.; Kim, Y.H.; Kim, S.W.; Non-ionic amphiphilic biodegradable PEG-PLGA-PEG copolymer enhances gene delivery efficiency in rat skeletal muscle. J Control Release 2007, 118, 245-253, 10.1016/j.jconrel.2006.11.025.
- Pomel, C.; Leborgne, C.; Cheradame, H.; Scherman, D.; Kichler, A.; Guegan, P.; Synthesis and evaluation of amphiphilic poly(tetrahydrofuran-b-ethylene oxide) copolymers for DNA delivery into skeletal muscle. Pharm Res 2008, 25, 2963-2971, 10.1007/s11095-008-9698-9.
- Pu, L.; Geng, Y.; Liu, S.; Chen, J.; Luo, K.; Wang, G.; Gu, Z.; .Electroneutralized amphiphilic triblock copolymer with a peptide dendron for efficient muscular gene delivery. ACS Appl Mater Interfaces 2014, 6, 15344-15351, 10.1021/am503808b.
- Pu, L.; Wang, J.; Li, N.; Chai, Q.; Irache, J.M.; Wang, G.; Tang, J.Z.; Gu, Z.; Synthesis of Electroneutralized Amphiphilic Copolymers with Peptide Dendrons for Intramuscular Gene Delivery. ACS Appl Mater Interfaces 2016, 8, 13724-13734, 10.1021/acsami.6b02592.
- Rasolonjatovo, B.; Illy, N.; Bennevault, V.; Mathé, J.; Midoux, P.; Le Gall, T.; Haudebourg, T.; Montier, T.; Lehn, P.; Pitard, B.; et al.et al. Temperature-Sensitive Amphiphilic Non-Ionic Triblock Copolymers for Enhanced In Vivo Skeletal Muscle Transfection. Macromol Biosci 2020, 20, e1900276, 10.1002/mabi.201900276.
- Richard, P.; Bossard, F.; Desigaux, L.; Lanctin, C.; Bello-Roufai, M.; Pitard, B.; Amphiphilic block copolymers promote gene delivery in vivo to pathological skeletal muscles. Hum Gene Ther 2005, 16, 1318-1324, 10.1089/hum.2005.16.1318.
- Jang, J.H.; Shea, L.D.; Intramuscular delivery of DNA releasing microspheres: microsphere properties and transgene expression. J Control Release 2006, 112, 120-128, 10.1016/j.jconrel.2006.01.013.
- Basarkar, A.; Singh, J.; Poly (lactide-co-glycolide)-polymethacrylate nanoparticles for intramuscular delivery of plasmid encoding interleukin-10 to prevent autoimmune diabetes in mice.. Pharm Res 2009, 26, 72-81, 10.1007/s11095-008-9710-4.
- Jeon, O.; Yang, H.S.; Lee, T.J.; Kim, B.S.; Heparin-conjugated polyethylenimine for gene delivery. J Control Release 2008, 132, 236-242, 10.1016/j.jconrel.2008.05.017.
- Wang, M.; Tucker, J.D.; Lu, P.; Wu, B.; Cloer, C.; Lu, Q.; Tris[2-(acryloyloxy)ethyl]isocyanurate cross-linked low-molecular-weight polyethylenimine as gene delivery carriers in cell culture and dystrophic mdx mice. Bioconjug Chem 2012, 23, 837-845, 10.1021/bc200674e.
- Kang, S.W.; Lim, H.W.; Seo, S.W.; Jeon, O.; Lee, M.; Nanosphere-mediated delivery of vascular endothelial growth factor gene for therapeutic angiogenesis in mouse ischemic limbs. Biomaterials 2008, 29, 1109-1117, 10.1016/j.biomaterials.2007.11.004.
- Hersh, J.; Condor Capcha, J.M.; Iansen Irion, C.; Lambert, G.; Noguera, M.; Singh, M.; Kaur, A.; Dikici, E.; Jiménez, J.J.; Shehadeh, L.A.; et al.et al. Peptide-Functionalized Dendrimer Nanocarriers for Targeted Microdystrophin Gene Delivery. Pharmaceutics 2021, 13, 2159-2175, 10.3390/pharmaceutics13122159.
- Lu, Q.L.; Bou-Gharios, G.; Partridge, T.A.; Non-viral gene delivery in skeletal muscle: a protein factory. Gene Ther 2003, 10, 131-142, 10.1038/sj.gt.3301874.
- Liu, J.; Song, L.; Liu, S.; Jiang, Q.; Liu, Q.; Li, N.; Wang, Z.G.; Ding, B.; A DNA-Based Nanocarrier for Efficient Gene Delivery and Combined Cancer Therapy. Nano Lett 2018, 18, 3328-3334 Nano Lett 2018, 18, 3328-3334, Nano Lett 2018, 18, 3328-3334, Nano Lett 2018, 18, 3328-3334.
- Zhou, X.; Xu, L.; Xu, J.; Wu, J.; Kirk, T.B.; Ma, D.; Xue, W.; Construction of a High-Efficiency Drug and Gene Co-Delivery System for Cancer Therapy from a pH-Sensitive Supramolecular Inclusion between Oligoethylenimine- graft-β-cyclodextrin and Hyperbranched Polyglycerol Derivative. ACS Appl Mater Interfaces 2018, 10, 35812-35829, 10.1021/acsami.8b14517.
- Mahendra, G.; Kumar, S.; Isayeva, T.; Mahasreshti, P.J.; Curiel, D.T.; Stockardt, C.R.; Grizzle, W.E.; Alapati, V.; Singh, R.; Siegal, G.P.; et al.et al. Antiangiogenic cancer gene therapy by adeno-associated virus 2-mediated stable expression of the soluble FMS-like tyrosine kinase-1 receptor. Cancer Gene Ther 2005, 12, 26-34, 10.1038/sj.cgt.7700754.
- Piekarowicz, K.; Bertrand, A.T.; Azibani, F.; Beuvin, M.; Julien, L.; Machowska, M.; Bonne, G.; Rzepecki, R.; A Muscle Hybrid Promoter as a Novel Tool for Gene Therapy. Mol Ther Methods Clin Dev 2019, 15, 157-169, 10.1016/j.omtm.2019.09.001.
- Le Bihan, M.C.; Bigot, A.; Jensen, S.S.; Dennis, J.L.; Rogowska-Wrzesinska, A.; Laine, J.; Gache, V.; Furling, D.; Jensen, O.N.; Voit, T.; et al.et al. In-depth analysis of the secretome identifies three major independent secretory pathways in differentiating human myoblasts. J Proteomics 2012, 77, 344-356, 10.1016/j.jprot.2012.09.008.
- Ost, M.; Coleman, V.; Kasch, J.; Klaus, S.; Regulation of myokine expression: Role of exercise and cellular stress. Free Radic Biol Med 2016, 98, 78-89, 10.1016/j.freeradbiomed.2016.02.018.
- Riu, E.; Mas, A.; Ferre, T.; Pujol, A.; Gros, L.; Otaegui, P.; Montoliu, L.; Bosch, F.; Counteraction of type 1 diabetic alterations by engineering skeletal muscle to produce insulin: insights from transgenic mice. Diabetes 2002, 51, 704-711, 10.2337/diabetes.51.3.704.
- Callejas, D.; Mann, C.J.; Ayuso, E.; Lage, R.; Grifoll, I.; Roca, C.; Andaluz, A.; Ruiz-de Gopegui, R.; Montané, J.; Muñoz, S.; et al.et al. Treatment of diabetes and long-term survival after insulin and glucokinase gene therapy. Diabetes 2013, 62, 1718-1729, 10.2337/db12-1113.
- Blaveri, K.; Heslop, L.; Yu, D.S.; Rosenblatt, J.D.; Gross, J.G.; Partridge, T.A.; Morgan, J.E.; Patterns of repair of dystrophic mouse muscle: studies on isolated fibersPatterns of repair of dystrophic mouse muscle: studies on isolated fibers. Dev Dyn 1999, 216, 244-256, 10.1002/(sici)1097-0177(199911)216:3<244::Aid-dvdy3>3.0.Co;2-9.
- Csapo, R.; Gumpenberger, M.; Wessner, B.; Skeletal Muscle Extracellular Matrix - What Do We Know About Its Composition, Regulation, and Physiological Roles? A Narrative Review. Front Physiol 2020, 11, 253, 10.3389/fphys.2020.00253.
- Gillies, A.R.; Lieber, R.L.; Structure and function of the skeletal muscle extracellular matrix. Muscle Nerve 2011, 44, 318-331, 10.1002/mus.22094.
- Ruponen, M.; Rönkkö, S.; Honkakoski, P.; Pelkonen, J.; Tammi, M.; Urtti, A.; Extracellular glycosaminoglycans modify cellular trafficking of lipoplexes and polyplexes. J Biol Chem 2001, 276, 33875-33880, 10.1074/jbc.M011553200.
- Caron, N.J.; Torrente, Y.; Camirand, G.; Bujold, M.; Chapdelaine, P.; Leriche, K.; Bresolin, N.; Tremblay, J.P.; Intracellular delivery of a Tat-eGFP fusion protein into muscle cells. Mol Ther 2001, 3, 310-318, 10.1006/mthe.2001.0279.
- Ruponen, M.; Honkakoski, P.; Rönkkö, S.; Pelkonen, J.; Tammi, M.; Urtti, A.; Extracellular and intracellular barriers in non-viral gene delivery. J Control Release 2003, 93, 213-217, 10.1016/j.jconrel.2003.08.004.
- Itaka, K.; Osada, K.; Morii, K.; Kim, P.; Yun, S.H.; Kataoka, K.; Polyplex nanomicelle promotes hydrodynamic gene introduction to skeletal muscle. J Control Release 2010, 143, 112-119, 10.1016/j.jconrel.2009.12.014.
- Walther, W.; Stein, U.; Siegel, R.; Fichtner, I.; Schlag, P.M.; Use of the nuclease inhibitor aurintricarboxylic acid (ATA) for improved non-viral intratumoral in vivo gene transfer by jet-injection. J Gene Med 2005, 7, 477-485, 10.1002/jgm.690.
- Sato, Y.; Yamauchi, N.; Takahashi, M.; Sasaki, K.; Fukaura, J.; Neda, H.; Fujii, S.; Hirayama, M.; Itoh, Y.; Koshita, Y.; et al.et al. In vivo gene delivery to tumor cells by transferrin-streptavidin-DNA conjugate. Faseb j 2000, 14, 2108-2118, 10.1096/fj.99-1052com.
- Chen, J.; Luo, J.; Zhao, Y.; Pu, L.; Lu, X.; Gao, R.; Wang, G.; Gu, Z.; Increase in transgene expression by pluronic L64-mediated endosomal/lysosomal escape through its membrane-disturbing action. ACS Appl Mater Interfaces 2015, 7, 7282-7293, 10.1021/acsami.5b00486.
- Yang, N.S.; Burkholder, J.; Roberts, B.; Martinell, B.; McCabe, D.; In vivo and in vitro gene transfer to mammalian somatic cells by particle bombardment. Proc Natl Acad Sci U S A 1990, 87, 9568-9572, 10.1073/pnas.87.24.9568.
- Mir, L.M.; Banoun, H.; Paoletti, C.; Introduction of definite amounts of nonpermeant molecules into living cells after electropermeabilization: direct access to the cytosol. Exp Cell Res 1988, 175, 15-25, 10.1016/0014-4827(88)90251-0.
- Mitragotri, S.; Healing sound: the use of ultrasound in drug delivery and other therapeutic applications. Nat Rev Drug Discov 2005, 4, 255-260, 10.1038/nrd1662.
- Liu, F.; Song, Y.; Liu, D.; Hydrodynamics-based transfection in animals by systemic administration of plasmid DNA. Gene Ther 1999, 6, 1258-1266, 10.1038/sj.gt.3300947.
- Akinc, A.; Thomas, M.; Klibanov, A.M.; Langer, R.; Exploring polyethylenimine-mediated DNA transfection and the proton sponge hypothesis. J Gene Med 2005, 7, 657-663, 10.1002/jgm.696.
- Wightman, L.; Kircheis, R.; Rössler, V.; Carotta, S.; Ruzicka, R.; Kursa, M.; Wagner, E.; Different behavior of branched and linear polyethylenimine for gene delivery in vitro and in vivo. J Gene Med 2001, 3, 362-372.
- Hu, Q.; Wang, J.; Shen, J.; Liu, M.; Jin, X.; Tang, G.; Chu, P.K.; Intracellular pathways and nuclear localization signal peptide-mediated gene transfection by cationic polymeric nanovectors. Biomaterials 2012, 33, 1135-1145, 10.1016/j.biomaterials.2011.10.023.
- Yi, W.J.; Yang, J.; Li, C.; Wang, H.Y.; Liu, C.W.; Tao, L.; Cheng, S.X.; Zhuo, R.X.; Zhang, X.Z.; Enhanced nuclear import and transfection efficiency of TAT peptide-based gene delivery systems modified by additional nuclear localization signals. Bioconjug Chem 2012, 23, 125-134, 10.1021/bc2005472.
- Ketola, T.M.; Hanzlíková, M.; Urtti, A.; Lemmetyinen, H.; Yliperttula, M.; Vuorimaa, E.; Role of polyplex intermediate species on gene transfer efficiency: polyethylenimine-DNA complexes and time-resolved fluorescence spectroscopy. J Phys Chem B 2011, 115, 1895-1902, 10.1021/jp109984c.
- Cai, J.; Yue, Y.; Rui, D.; Zhang, Y.; Liu, S.; Wu, C.; Effect of Chain Length on Cytotoxicity and Endocytosis of Cationic Polymers. Macromolecules 2011, 44, 2050-2057, 10.1021/ma102498g.
- Kunath, K.; von Harpe, A.; Fischer, D.; Petersen, H.; Bickel, U.; Voigt, K.; Kissel, T.; Low-molecular-weight polyethylenimine as a non-viral vector for DNA delivery: comparison of physicochemical properties, transfection efficiency and in vivo distribution with high-molecular-weight polyethylenimine. J Control Release 2003, 89, 113-125, 10.1016/s0168-3659(03)00076-2.
- Wirth, T.; Parker, N.; Ylä-Herttuala, S.; History of gene therapy. Gene 2013, 525, 162-169, 10.1016/j.gene.2013.03.137.
- Lai, T.C.; Kataoka, K.; Kwon, G.S.; Bioreducible polyether-based pDNA ternary polyplexes: balancing particle stability and transfection efficiency. Colloids Surf B Biointerfaces 2012, 99, 27-37, 10.1016/j.colsurfb.2011.09.026.
- Grandinetti, G.; Smith, A.E.; Reineke, T.M.; Membrane and nuclear permeabilization by polymeric pDNA vehicles: efficient method for gene delivery or mechanism of cytotoxicity?. Mol Pharm 2012, 9, 523-538, 10.1021/mp200368p.
- Roques, C.; Bouchemal, K.; Ponchel, G.; Fromes, Y.; Fattal, E.; Parameters affecting organization and transfection efficiency of amphiphilic copolymers/DNA carriers. J Control Release 2009, 138, 71-77, 10.1016/j.jconrel.2009.04.030.
More
Information
Subjects:
Materials Science, Biomaterials
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.4K
Entry Collection:
Biopharmaceuticals Technology
Revisions:
2 times
(View History)
Update Date:
25 Nov 2022
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No