Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Vikash Kumar | -- | 3486 | 2022-11-23 10:00:34 | | | |
| 2 | Amina Yu | + 11 word(s) | 3497 | 2022-11-24 01:55:44 | | | | |
| 3 | Amina Yu | Meta information modification | 3497 | 2022-11-24 01:57:33 | | | | |
| 4 | Amina Yu | -2 word(s) | 3495 | 2022-11-25 07:24:46 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Kumar, V.; Roy, S.; Behera, B.K.; Das, B.K. Heat Shock Proteins in Cellular Homeostasis. Encyclopedia. Available online: https://encyclopedia.pub/entry/36051 (accessed on 08 August 2026).
Kumar V, Roy S, Behera BK, Das BK. Heat Shock Proteins in Cellular Homeostasis. Encyclopedia. Available at: https://encyclopedia.pub/entry/36051. Accessed August 08, 2026.
Kumar, Vikash, Suvra Roy, Bijay Kumar Behera, Basanta Kumar Das. "Heat Shock Proteins in Cellular Homeostasis" Encyclopedia, https://encyclopedia.pub/entry/36051 (accessed August 08, 2026).
Kumar, V., Roy, S., Behera, B.K., & Das, B.K. (2022, November 23). Heat Shock Proteins in Cellular Homeostasis. In Encyclopedia. https://encyclopedia.pub/entry/36051
Kumar, Vikash, et al. "Heat Shock Proteins in Cellular Homeostasis." Encyclopedia. Web. 23 November, 2022.
Copy Citation
Heat shock proteins (Hsps) are a family of ubiquitously expressed stress proteins and extrinsic chaperones that are required for viability and cell growth in all living organisms. These proteins are highly conserved and produced in all cellular organisms when exposed to stress. Hsps play a significant role in protein synthesis and homeostasis, as well as in the maintenance of overall health in crustaceans against various internal and external environmental stresses. Recent reports have suggested that enhancing in vivo Hsp levels via non-lethal heat shock, exogenous Hsps, or plant-based compounds, could be a promising strategy used to develop protective immunity in crustaceans against both abiotic and biotic stresses.
heat shock proteins
crustaceans
protein homeostasis
protective immunity
1. Regulation of the Heat Shock Protein Response
Heat shock proteins (Hsps) are evolutionarily ancient and extremely conserved proteins found in almost all living organisms, ranging from archaebacteria, prokaryotes, and eukaryotes. Hsps play an essential role in regulating cellular metabolism in stressful conditions. These proteins are present in several intracellular locations, e.g., in the nucleus, mitochondria, endoplasmic reticulum (ER), and chloroplast and cytosol of eukaryotes [1][2][3]. In general, Hsps are classified into several families based on their function, molecular mass, and sequence homology (i.e., Hsp60, Hsp70, Hsp90, and sHsps) (Figure 1) and also, they can be grouped according to their nearest size family (e.g., Hsp84, Hsp85, and Hsp86 in the Hsp90 kDa family) [4].
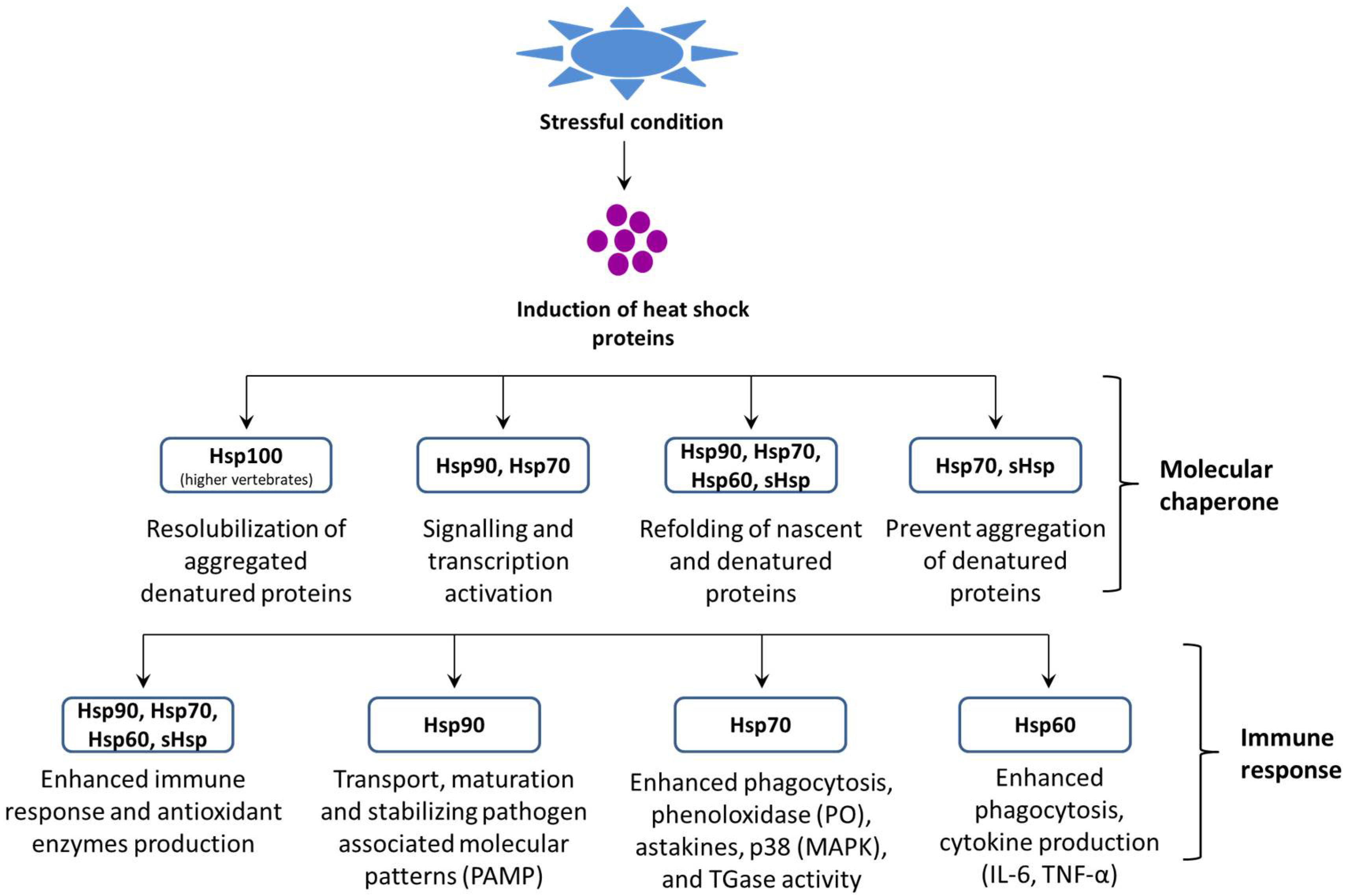
Figure 1. Role of heat shock proteins (Hsps) in proteostasis and the host immune response.
Hsps plays a key role in cellular processes that occur during and after exposure to oxidative stress that is caused by hazardous environmental and/or microbial agents. As a result, the normal intracellular reducing environment is compromised, which leads to oxidation and aggregation of key proteins and DNA, ultimately resulting in cellular dysfunction. Due to their versatile functions, Hsps can intervene following oxidative stress at several levels [5][6][7]. Firstly, some Hsps, mainly the Hsp70 family members, play a crucial role in protein sorting and quality control via selecting and directing abnormal proteins to the proteasome or lysosomes for degradation; thus, Hsps aid the clearance of damaged proteins [8]. In some cases, where misfolded proteins need to be rescued, the same machinery facilitates the correct folding of damaged proteins. Moreover, the Hsp families, such as Hsp27, Hsp70, and Hsp90, can negatively regulate apoptosis via the binding and inhibiting of members of the apoptotic cascade. Some Hsps have immuno-enhancing actions (for details, see the section on Hsps and immunity). Among the different Hsp families, Hsp70 is the largest and most highly conserved of the stress protein families. At least 121 proteins have been characterized within this family, and cross-hybridization occurs across various species, such as in mammals, fish, and mollusks [9][10][11].
After the discovery of heat shock proteins, it was clear that the heat shock protein response requires a specific transcription factor [12][13][14][15][16]. In crustaceans, a few reports have suggested that the heat shock transcription factor plays a significant role in determining the heat shock response and also in tolerance against stressful conditions. Tan and Macrae [17] reported that the heat shock factor 1 (Hsf1) transcription factor induces stress tolerance (drying) in diapausing brine shrimp cysts and is responsible for the improved growth and survival of A. franciscana [17]. In another study, Sornchuer et al. [18] demonstrated that Hsf1 has an important role in the thermal stress response and regulates the transcription of heat shock proteins and immune-related genes in P. monodon [18]. However, detailed investigations still need to be carried out and more information need to be obtained in order to characterize the mechanism of heat shock response regulation in crustaceans. Moreover, in eukaryotes, the heat shock response is well documented, and it was showed that the induction of Hsp transcription is mediated by a pre-existing transcription factor, i.e., the heat shock factor or the heat shock transcription factor (HSF) [19]. The HSF upon activation binds to the heat shock element (HSE) at the promoter region (5’ upstream end) of the Hsp gene and induces the transcription of Hsp [20]. The binding motif of the HSE is composed of nGAAn blocks (5 bp) in alternate orientation, and for the stable binding of the HSF and the HSE, at least three units are required [21]. In prokaryotes (e.g., E. coli), the σ32 regulatory protein is responsible for Hsp expression [22]. Additionally, as an alternate subunit of the bacterial RNA polymerase, σ32 replaces the σ70 normal regulatory proteins during heat stress [23]. HSF1 and σ32 share basic mechanistic properties, however their structure or sequence are not related; further, it has been found that protein homeostasis disturbance results in the activation of hsf1 and σ32 [24].
HSF genes have been reported from yeast (Saccharomyces cerevisiae), plants (Arabidopsis sp.), fruit flies (Drosophila sp.), chicken (Gallus sp.), A. franciscana, and P. monodon [25][26][27][28][29]. Sequence comparison of the HSF gene from different species showed that DNA binding and the oligomerization domain is strongly conserved. The HSF contains two highly conserved regions, i.e., a ~100 amino acids NH2-terminal DNA binding domain and an adjacent trimerization domain having 3 hydrophobic heptad repeats, leu zippers [3]. The activation of the HSF involves cellular factors as the intermediary sensors that regulate the activity of the HSF during non-stressful conditions [30]. In animals, the HSF is maintained as a monomeric form through transient interaction with Hsps. However, during stressful conditions, the HSF was released and formed trimers with the HSE, which resulted in the induction of Hsps. Subsequently, Hsps bind with denatured/misfolded protein aggregates and maintain the homeostatic condition in the cells [31][32][33][34][35].
Based on previous studies, a hypothetical illustration of the possible methods involved in the activation of the heat shock protein response, and its potential role in maintaining homeostasis and host health, is depicted in Figure 2. Briefly, the stressful stimuli, heat-shock-protein-inducing (Hspi) conditions (e.g., environmental, pathological, and physiological) [36][37], or compounds (e.g., plant-based, natural polymers and so on) [38][39] develop oxidative status inside the host. Subsequently, it facilitates the phosphorylation and nuclear translocation of the HSF (in the native form present in the non-phosphorylated form attached with heat shock proteins such as Hsp90) [40] where (inside the hemocyte nucleus) the HSF binds with the HSE at the promoter region (5’ upstream end) of the Hsp gene and induces the transcription of the Hsp gene and production of Hsps [41][42]. The Hsp (e.g., Hsp70) functions as a non-covalently molecular chaperone bind with a hydrophobic exposed segment of unfolded proteins [43][44]. Further, this prevents the aggregation of inappropriate or unfolded proteins; inhibits the misfolding of the polypeptides; transports immature polypeptides to target organelle for the purposes of final packaging and repair, as well as denaturation or degradation of misfolded proteins through proteasomes or lysosomes (proteolysis), which cannot be repaired [45]; and, also, maintains protein homeostasis. The extracellular Hsp also functions as a chaperokine and binds with Toll-like receptors (e.g., TLR-4), which are expressed on hemocytes that lead to the maturation and activation of hemocytes [46][47][48]. The activated hemocytes induce the production of astakines that binds with the binding site of a pathogen in the host cell and decrease the chance of the pathogen attaching to its host cells [49][50][51]. The hemocytes (activated by Hsps) also induce the expression of transglutaminase (Tgase) [52], which form a blood clot in the presence of Ca2+ and prevents the spread of pathogens and increases the expression of antimicrobial peptides (e.g., crustin, lysozyme and so on) that has bactericidal activity against Gram-positive and Gram-negative bacteria [53][54][55][56]. The p38 (mitogen-activated protein kinase, MAPK) activated by hemocytes, increases the expression of antimicrobial peptides (e.g., crustin, lysozyme and so on) and has a critical role in defense against bacterial and viral infection [57][58][59][60][61]. Hemocytes activate the ProPO cascade in the presence of Ca2+, leading to melanization and further killing of the pathogens [62][63][64][65].
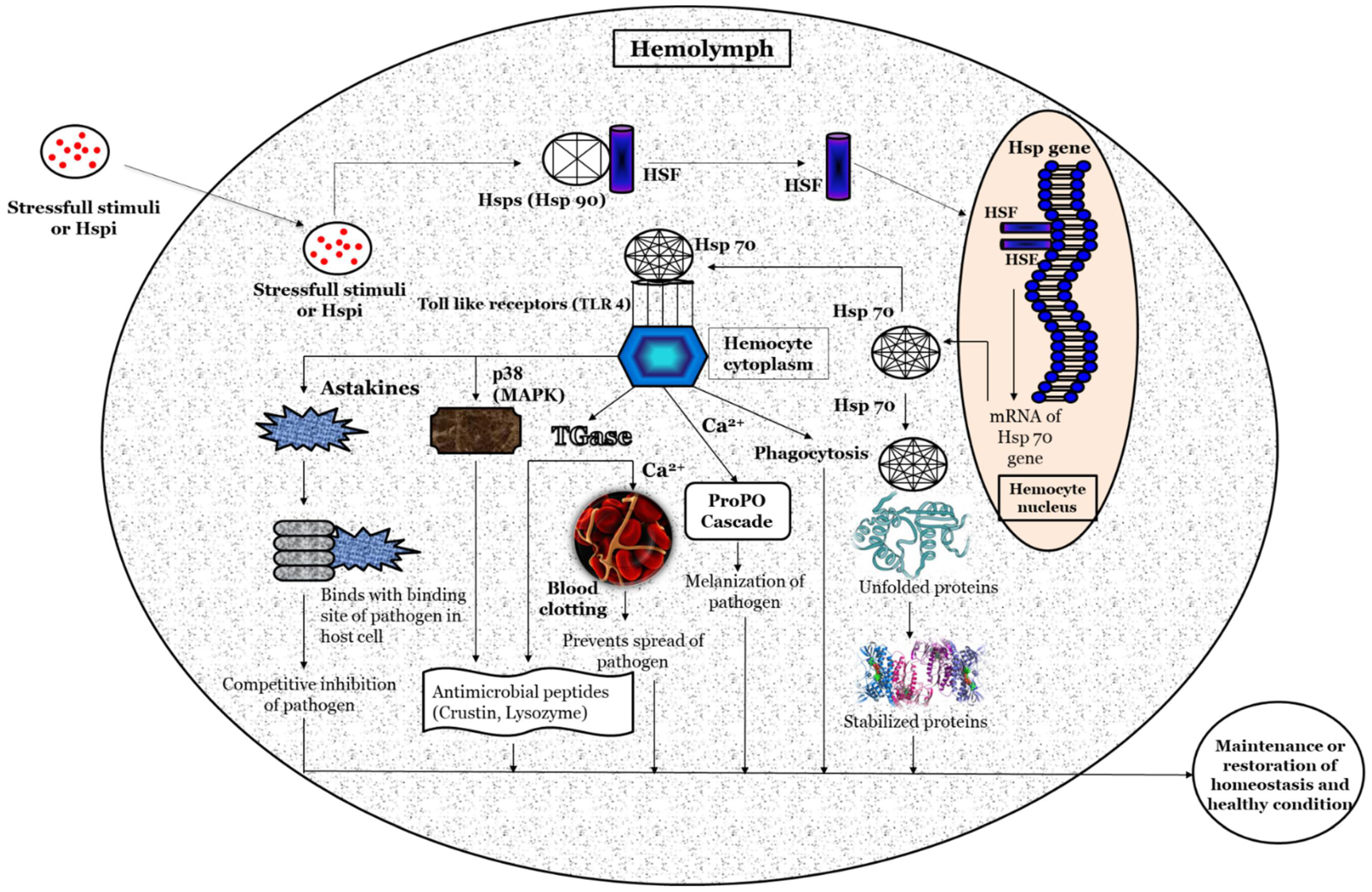
Figure 2. A schematic hypothetical illustration of the possible mechanism of action for Heat shock proteins (Hsps) in crustaceans.
2. Factors Modulate Heat Shock Protein Response
The induction of Hsps in response to cellular stressors was initially considered a short-term functional response, with a range of essential housekeeping and cytoprotective functions. These stressors, in general, induce protein damage and increase the susceptibility of host animals further to subsequent stressful conditions. However, accumulating pieces of evidence, over the past few decades, have suggested that Hsps play a significant role in the regulation of the immune response in invertebrates (Figure 3). The heat shock protein expressions influenced by either abiotic or biotic stresses are summarized in the section below.
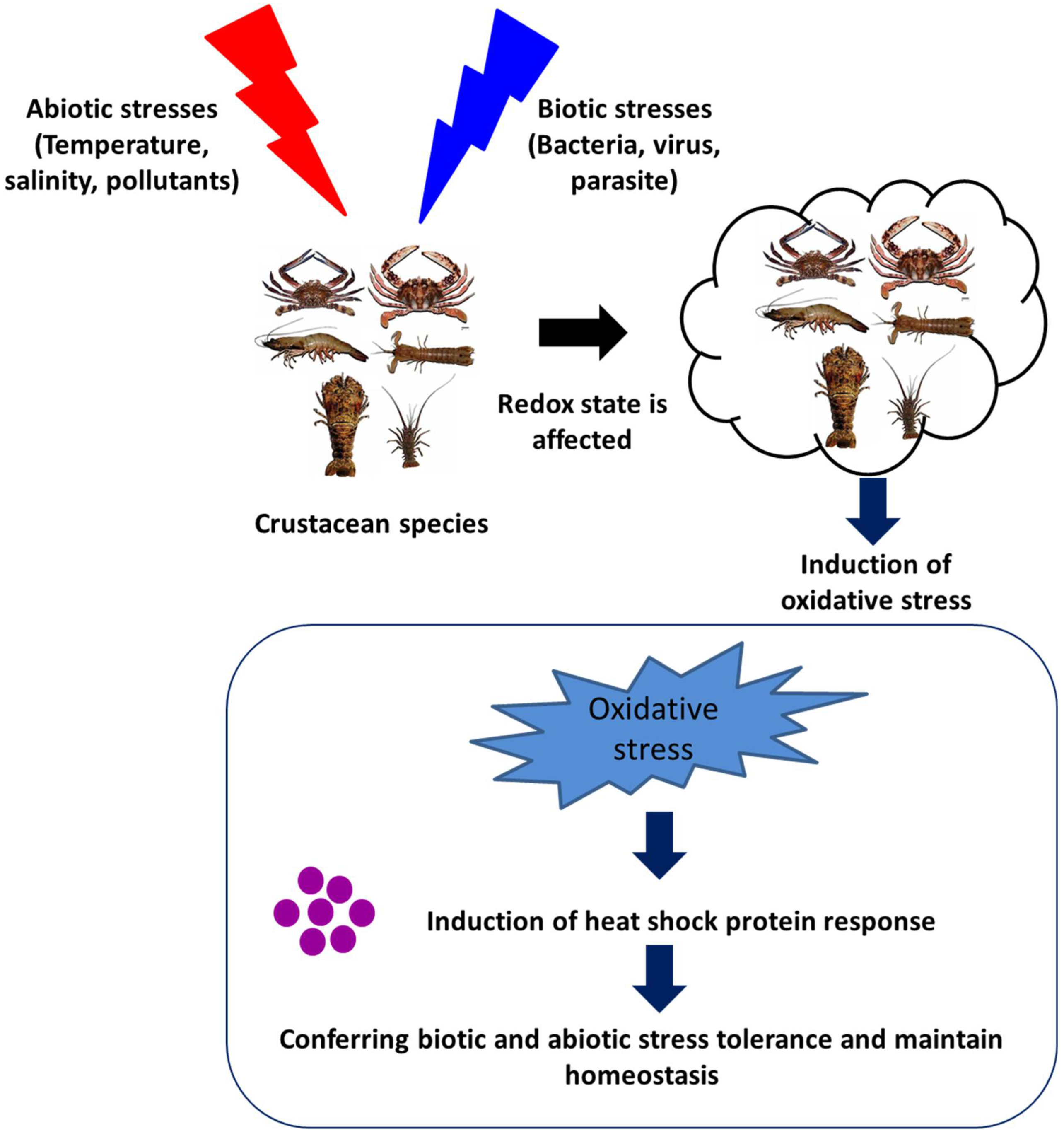
Figure 3. Effect of abiotic and biotic stresses on host health and heat shock protein response.
2.1. Environmental or Abiotic Stresses
The physiological status of crustaceans is greatly influenced by their environmental conditions. A slight variation in environmental parameters creates a stressful condition that attenuates the immune system and increases the susceptibility of animals to microbial infection.
2.1.1. Temperature
Temperature is considered an important abiotic stressor, as a slight change in water temperature can affect the body physiology and health of crustaceans [66][67][68]. Interestingly, heat shock proteins are amongst the most significant proteins that are induced by hypo and hyperthermia and their role in protection against thermal stress has been well documented [69][70][71][72][73]. Among the Hsp multigenic family, few proteins are expressed at extremely low levels under normal conditions, while the transcription of most Hsps increased significantly in response to stresses, e.g., stress-inducible proteins (Hsp70). However, Hsps that are expressed constitutively under normal conditions, and may be upregulated during stress conditions, are generally known as heat shock cognate proteins, e.g., Hsc70 [74][75][76][77]. Hsps play a central role in thermotolerance by promoting growth at moderately high temperatures and protecting the organism from mortality at extremely high temperatures [78]. In general, Hsps are induced in both hyperthermia and hypothermia conditions. For example, an increase in the water temperature has been found to induce the production of Hsp70 in Ferropenaeus chinensis, Chinese white shrimp [79], Gammarus pulex, freshwater crustacean [80], and A. franciscana, [81]. Moreover, increased Hsp (Hsp90 and Hsp40) expression was also reported during 6 h of cold shock at 1 and 6 °C in A. franciscana [82]. Hsp70 has been demonstrated to play an important role in protecting cells from damage in S. paramamosain in response to thermal stress (this was shown in an increase in 11 °C from normal growth temperature) [83]. In addition, the adult A. franciscana, when exposed to sub-lethal heat shock (37 °C for 30 min), induces the transcription of Hsp70, Hsp67, and Hsc70, resulting in an improved tolerance of brine shrimp to high temperatures [84]. Interestingly, marine invertebrates are very sensitive to high temperatures and there are a myriad of reports suggesting that Hsp70 is upregulated in response to heat stress [85][86][87]. These studies indicate that Hsps improve thermal tolerance in crustaceans and provide protection in both hyperthermia and hypothermia conditions.
2.1.2. Salinity
The concentration of dissolved inorganic salt concentrations or the salinity in the water is reported to affect the osmoregulation of crustaceans and induce cellular damage, including a deleterious effect on the folding and transformation of polypeptides [88][89][90][91]. Moreover, the osmotic-stress-induced Hsp production plays a crucial role in the maintenance of biological processes, as well as the protection of crustaceans against stressful conditions [92][93][94][95][96]. The study by Yang et al. [51] reported that high salinity stress increased the expression of Hsp70 and that this could lead to enhanced resistance in S. paramamosain against changes in water salinity. The possible mechanism behind the protective action of Hsps against salinity stress is that the osmotic change increases the metabolism rate and enhances the stress response resulting in increased Hsp production [97]. The increased Hsp response subsequently enhances the immune response, including lysozyme, phenoloxidase, and peroxide activity and provides rapid protection against osmotic stress until the organic osmolytes are fully accumulated [98][99]. Water salinity was reported as an important factor for the purposes of natural growth of crustaceans; further, studies on P. trituberculatus have shown that variable salinity significantly influences larval development [100]. Xu and Qin [33] found that Hsp60 has an important role in both the cellular and humoral stress response of the swimming crab, P. trituberculatus, and that these responses regulate the salinity stress via an intrinsic pathway. Further, these responses also play an essential role in protecting the swimming crab against salinity stress. In addition, a few reports have suggested that an increase in the expression of Hsp70 enhances immune response and confers protection to A. franciscana against hypersalinity stress [101].
2.1.3. Environmental Pollutants
The environmental pollutants induced heat shock proteins (Hsps) expression in crustaceans are the most frequently studied in the literature [102][103][104]. The study of heat shock proteins in invertebrates started in the 1990s, and the first observation conducted on Hsps was made by Köhler et al. [105]. The study showed that the exposure of three diplopods (Tachypodoiulus niger, Cylindroiulus punctatus, and Glomeris marginata), one isopod (Oniscus asellus) and two slugs (Arion ater and Deroceras reticulatum) to heavy metals/molluscicides resulted in the increased expression of Hsp70 [106].
Studies on the effect of environmental pollutants on crustaceans demonstrated that the Hsp gene expression is induced by several chemical stresses. For example, this can be found in: nonylphenol (NP) (used in the polymer industry); bisphenol A diglycidyl ether (BPA) (intermediate in the production of polycarbonate and epoxy resins) [107]; 17α-ethynyl estradiol (EE) (synthetic estrogen) [108]; bis(2-ethylhexyl) phthalate (DEHP) (plasticizer in polymer products); endosulfan (ES) (organochlorine insecticide); chloropyriphos (CP) (organophosphorus insecticide); paraquat dichloride (PQ) (oxygen radical generating herbicide); Cadmium (Cd); lead (Pb) and potassium dichromate (Cr) (heavy metals); and benzo[a] pyrene (BaP) (polycyclic aromatic hydrocarbon) [109]. Moreover, for instance, the sublethal concentration of endosulfan has been reported to enhance the synthesis of Hsp70 and Hsp90 in monsoon river prawn, i.e., M. malcolmsonii and P. monodon [110][111][112][113]. In another study, the mixture of environmental pollutant chemicals has been reported to modulate the physiological, as well as immunity and survival responses in crustaceans. For instance, Park and Kwak [61] demonstrated that the application of bisphenol A (BPA) and 4-nonlphenol (NP)—an endocrine disrupting chemical (EDCs)—at different concentrations and at different time intervals (12, 24, 48, and 96 h) induce the expression of Hsp90; further, when exposed to BPA and NP, the marine crab, Charybdis japonica, has a significantly increased survival [26]. Additionally, chemical stress was reported to induce the production of Hsps, which helps in maintaining the homeostasis and structural integrity of cells [114][115].
2.2. Biotic Stresses
The heat shock proteins that are induced by biotic stresses play a very crucial role in protein folding, immune enhancement, and cross-protection against infectious diseases [116][117][118]. Although, there are several reports that demonstrate that Hsps are easily induced by abiotic stresses including heat, salinity and so on [119], very little information is available on crustaceans’ Hsp response against biotic stresses, including bacteria, parasites, and viruses. Some of the recent research findings have suggested that members of the Hsp70 family have been identified in crustaceans, which are involved in the response to biotic stresses, mainly bacteria, parasites, and viruses [120][121]. Zhou et al. [80] demonstrated that L. vannamei when challenged with V. alginolyticus (Gram-negative) and S. aureus (Gram-positive) bacteria have a significantly increased expression of L. vannamei Hsp60 (LvHSP60) and Hsp70 (LvHSP70) gene in the gills, hepatopancreas, and hemocytes. In another study, temporal transcription of LvHSP70, following the white spot syndrome virus (WSSV) challenge, has been reported to induce an anti-WSSV innate immune response in L. vannamei [122].
In the swimming crab, Portunus trituberculatus, transcription of the P. trituberculatus, the Hsp70 (PtHsp70) gene was shown to increase very rapidly in response to the bacterial challenge with V. alginolyticus [123]. Similar findings were reported by Yang et al. [6], i.e., the fact that there was an increased transcription of Hsp70 in hemocytes of S. paramamosain after the V. alginolyticus challenge and were involved in generating cross-protection in the mud crab. Additionally, Hsp90, which plays a crucial role in protein biosynthesis, signal transduction, and immune responses, was also shown to induce protective immunity in crustaceans. Huang et al. [124] analyzed the role of Hsp90 in S. paramamosain in response to microbial infection, and the results showed that the transcription of SpHSP90 was upregulated in mud crabs after being challenged with Staphylococcus aureus, white spot syndrome virus (wssv), and V. harveyi [124].
Hsps exert their physiological effect via assisting in the formation of new polypeptides as well as in the protecting and maintaining of the host cell polypeptides and naïve proteins from denaturation during microbial infection [125][126][127]. In crustaceans, the host immune response against microbial infection [94] is often associated with reactive oxygen species (ROS) production [128]. It has been demonstrated that in L. vannamei, after bacterial challenge, the subsequent induction of the host immune response leads to an increase in ROS levels [129]. While microbial infection-induced ROS mostly has antimicrobial activity, the production of ROS can result in the denaturation of proteins (proteotoxicity) in the host cell itself; as such, in this condition, the induced Hsps display a cytoprotective role and act as chaperone proteins in order to maintain the protein homeostasis and preserve cellular structures [130].
3. An A. franciscana Model System to Investigate the Role of Hsps in Crustaceans
The A. franciscana is a small branchiopod crustacean that is highly osmotolerant and reported from several harsh environmental conditions worldwide [131][132][133][134]. They live in an environment of severe hypersalinity, high levels of ultraviolet radiation, fluctuating oxygen concentration, and extreme temperature [135][136]. The oviparous development in A. franciscana leads to the production of hard-shell covering diapause cysts, which are composed of stress-tolerant metabolically inactive embryos stalled at gastrulation and that which can remain in stasis for several years [137][138][139][140]. However, when the diapause cysts were immersed under appropriate conditions in seawater with aeration and temperatures, the hard-shell raptures and cysts develop, releasing swimming larvae within 24 h (Figure 4) [140][141][142].
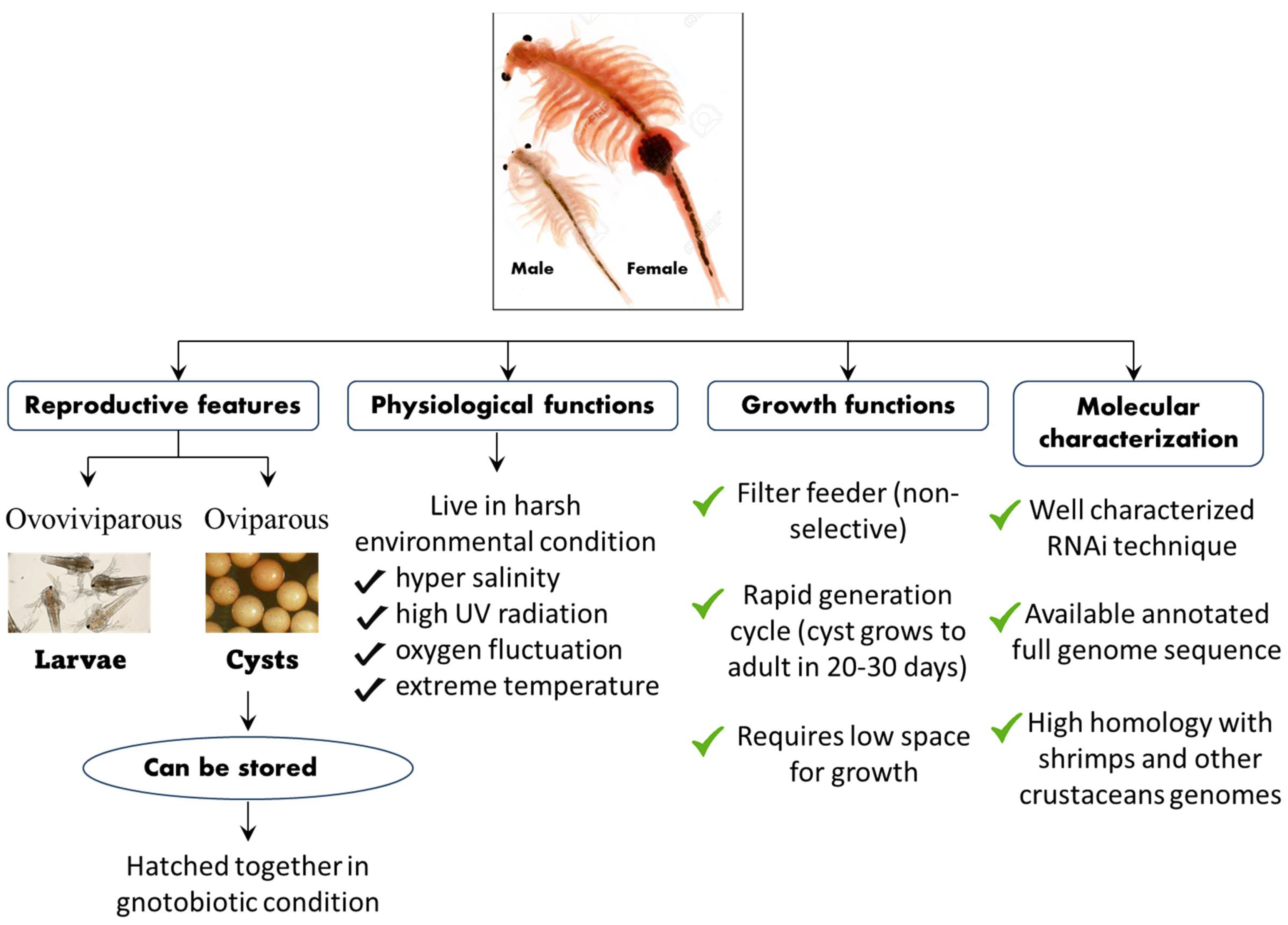
Figure 4. Schematic representation of the life cycle, physiological, growth, and molecular features of the A. franciscana model system.
Apart from its interesting life history, A. franciscana are non-selective filter feeders (which can be grown in a wide range of feed resources), have a rapid generation cycle (the cyst grows to adult in 20–30 days), require very low space for growth (hence have a relatively smaller cost to culture), and developmental stages are well characterized. This aspect of their stages being well characterized, for instance, is shown in the fact that the A. franciscana produces encysted gastrulae cysts during oviparous development, while ovoviviparous development provides live larvae in both sexual and asexual (parthenogenetic) stages. Additionally, gnotobiotic (germ-free) culture conditions (allowing full control over the host-associated microbial communities) and advanced molecular techniques including qPCR and RNAi are well established in A. franciscana, which makes this species an exceptional model organism that can be used in order to investigate the host–pathogen relationship and to study the biological activity of protective compounds [143][144][145][146][147][148][149][150][151][152]. In addition, cysts of A. franciscana can be stored for a couple of years in the fridge and, after terminating the diapause stage, the cysts from different generations can be used and hatched all together, simultaneously. This excellent facility of storage and hatching, permits one to perform experiments on demand, which can help to avoid or minimize environmental influences. Above all, the genome sequence of A. franciscana showed that it shares a very high homology with shrimps and other crustaceans’ genomes (Figure 5). Therefore, there is a high possibility that the outcome of studies based on A. franciscana would provide a fundamental basis to understand the host–pathogen interactions in other commercially important shrimp species.
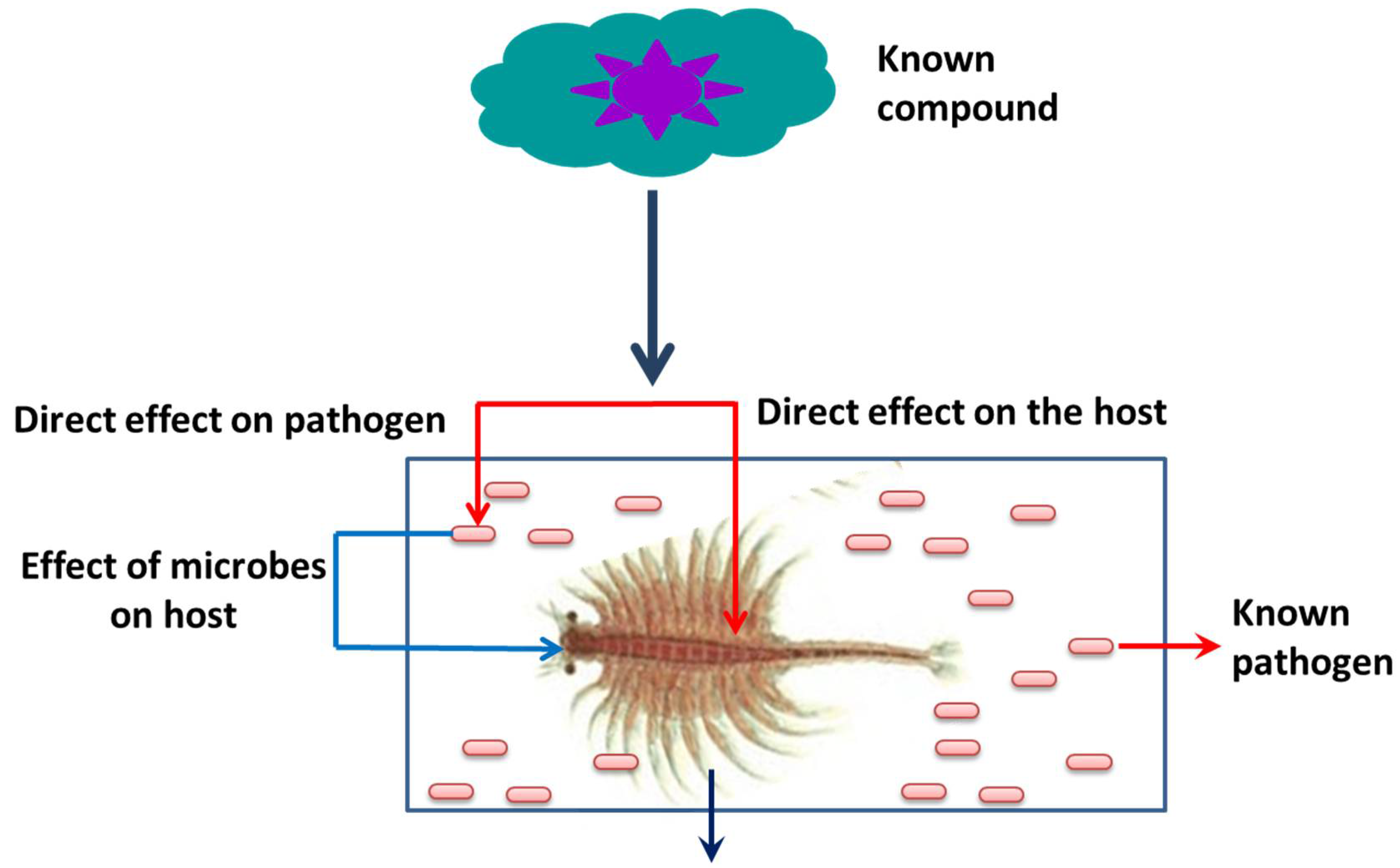
Figure 5. Advantage of a gnotobiotic (germ free) A. franciscana model system.
The induction of Hsp production inside the host in order to control diseases in aquaculture was investigated using the model organism, brine shrimp larvae [153][154]. In 1988, Miller and McLennan observed the presence of heat shock proteins in the early developmental stages of brine shrimp, i.e., in encysted gastrula embryos (cysts) and newly hatched nauplius larvae. They have reported that the larvae exhibited induced thermotolerance, which is associated with the synthesis and upregulation of heat shock proteins [155]. The role of heat shock protein response in adult A. franciscana, in response to high temperature (including LT50 determination, enhanced thermotolerance, and increased production of the Hsp70 family stress protein) was studied by Frankenberg et al. [156]. Results demonstrated that Hsp70 family proteins (mainly Hsp67 and Hsc70) levels were significantly upregulated during sublethal heat shock (37 °C for 30 min). The A. franciscana exposed to Zn-control/Cd-control treatment induced the expression of Hsp and increased the partitioning of Cd to the tropically available metal (TAM), which could result in the bio-enhancement of Cd trophic transfer to predators, which, in turn, leads to the suppression of Zn accumulation in A. franciscana [157].
Later, a gnotobiotic (germ-free) culture system was developed for brine shrimp and several studies have demonstrated that this provides a fully controlled and excellent host–pathogen environment and facilitates the determining of the effect of external stimuli on the host (Figure 5) [158]. The gnotobiotic system also avoids the interference generated by host-associated microorganisms as well as shifts in the composition of microbial diversity [159][160][161][162][163]. The effect of non-lethal heat shock (NLHS) on host survival, immune response, and protection against stressful conditions were also studied by several researchers. Results showed that NLHS increased the transcription of Hsp70, which enhances the immune response and provides cross-protection from environmental, physiological, and microbial stress in A. franciscana [164][165][166][167][168][169]. In another study, Sung et al. [36] demonstrated that non-pathogenic CAG 629 and CAG 626 E. coli strains, when heat shocked and administered through feed to Artemia larvae, resulted in enhanced Hsp70 expression (~two folds) and subsequent protection against V. campbellii infection [170].
References
- Nover, L.; Scharf, K.D.; Neumann, D. Cytoplasmic heat shock granules are formed from precursor particles and are associated with a specific set of mRNAs. Mol. Cell. Biol. 1989, 9, 1298–1308.
- Parsell, D.A.; Lindquist, S. The Function of Heat-Shock Proteins in Stress Tolerance: Degradation and Reactivation of Damaged Proteins. Annu. Rev. Genet. 1993, 27, 437–496.
- Craig, E.A.; Weissman, J.S.; Horwich, A.L. Heat shock proteins and molecular chaperones: Mediators of protein conformation and turnover in the cell. Cell 1994, 78, 365–372.
- Dong, H.; Roy, S.; Zheng, X.; Kumar, V.; Das, B.K.; Duan, Y.; Sun, Y.; Zhang, J. Dietary teprenone enhances non-specific immunity, antioxidative response and resistance to hypoxia induced oxidative stress in Lateolabrax maculatus. Aquaculture 2021, 533, 736126.
- Dong, H.; Zheng, X.; Kumar, V.; Roy, S.; Duan, Y.; Gao, H.; Zhang, J. Dietary supplementation of teprenone potentiates thermal and hypoxia tolerance as well as cellular stress protection of Epinephelus coioides juveniles reared under multiple stressors. Aquaculture 2020, 514, 734413.
- Yang, Y.; Ye, H.; Huang, H.; Li, S.; Liu, X.; Zeng, X.; Gong, J. Expression of Hsp70 in the mud crab, Scylla paramamosain in response to bacterial, osmotic, and thermal stress. Cell Stress Chaperones 2013, 18, 475–482.
- Lindquist, S.; Craig, E.A. The Heat-Shock Proteins. Annu. Rev. Genet. 1988, 22, 631–677.
- Iryani, M.T.M.; MacRae, T.H.; Panchakshari, S.; Tan, J.; Bossier, P.; Wahid, M.E.A.; Sung, Y.Y. Knockdown of heat shock protein 70 (Hsp70) by RNAi reduces the tolerance of Artemia franciscana nauplii to heat and bacterial infection. J. Exp. Mar. Biol. Ecol. 2017, 487, 106–112.
- Roberts, R.J.; Agius, C.; Saliba, C.; Bossier, P.; Sung, Y.Y. Heat shock proteins (chaperones) in fish and shellfish and their potential role in relation to fish health: A review. J. Fish Dis. 2010, 33, 789–801.
- Sung, Y.Y.; Macrae, T.H.; Sorgeloos, P.; Bossier, P. Stress response for disease control in aquaculture. Rev. Aquac. 2011, 3, 120–137.
- Robert, J. Evolution of heat shock protein and immunity. Dev. Comp. Immunol. 2003, 27, 449–464.
- Norouzitallab, P.; Baruah, K.; Muthappa, D.M.; Bossier, P. Non-lethal heat shock induces HSP70 and HMGB1 protein production sequentially to protect Artemia franciscana against Vibrio campbellii. Fish Shellfish Immunol. 2015, 42, 395–399.
- Norouzitallab, P.; Baruah, K.; Biswas, P.; Vanrompay, D.; Bossier, P. Probing the phenomenon of trained immunity in invertebrates during a transgenerational study, using brine shrimp Artemia as a model system. Sci. Rep. 2016, 6, 21166.
- Lindquist, S. The Heat-Shock Response. Annu. Rev. Biochem. 1986, 55, 1151–1191.
- Richter, K.; Haslbeck, M.; Buchner, J. The Heat Shock Response: Life on the Verge of Death. Mol. Cell 2010, 40, 253–266.
- Courgeon, A.-M.; Maisonhaute, C.; Best-Belpomme, M. Heat shock proteins are induced by cadmium in Drosophila cells. Exp. Cell Res. 1984, 153, 515–521.
- Tan, J.; Macrae, T.H. Stress tolerance in diapausing embryos of Artemia franciscana is dependent on heat shock factor 1 (Hsf1). PLoS ONE 2018, 13, e0200153.
- Sornchuer, P.; Junprung, W.; Yingsunthonwattana, W.; Tassanakajon, A. Heat shock factor 1 regulates heat shock proteins and immune-related genes in Penaeus monodon under thermal stress. Dev. Comp. Immunol. 2018, 88, 19–27.
- Yura, T.; Tobe, T.; Ito, K.; Osawa, T. Heat shock regulatory gene (htpR) of Escherichia coli is required for growth at high temperature but is dispensable at low temperature. Proc. Natl. Acad. Sci. USA 1984, 81, 6803–6807.
- Michel, G.P.; Starka, J. Effect of ethanol and heat stresses on the protein pattern of Zymomonas mobilis. J. Bacteriol. 1986, 165, 1040–1042.
- Ananthan, J.; Goldberg, A.L.; Voellmy, R. Abnormal Proteins Serve as Eukaryotic Stress Signals and Trigger the Activation of Heat Shock Genes. Science 1986, 232, 522–524.
- Csermely, P.; Schnaider, T.; So, C.; Prohászka, Z.; Nardai, G. The 90-kDa Molecular Chaperone Family: Structure, function, and clinical applications. a comprehensive review. Pharmacol. Ther. 1998, 79, 129–168.
- Bozaykut, P.; Ozer, N.K.; Karademir, B. Regulation of protein turnover by heat shock proteins. Free Radic. Biol. Med. 2014, 77, 195–209.
- Aridon, P.; Geraci, F.; Turturici, G.; D’Amelio, M.; Savettieri, G.; Sconzo, G. Protective Role of Heat Shock Proteins in Parkinson’s Disease. Neurodegener. Dis. 2011, 8, 155–168.
- Capy, P.; Gasperi, G.; Biémont, C.; Bazin, C. Stress and transposable elements: Co-evolution or useful parasites? Heredity 2000, 85, 101–106.
- Horowitz, A.; Horowitz, S. Disease control in shrimp aquaculture from a microbial ecology perspective. In The New Wave, Proceedings of the Special Session on Sustainable Shrimp Culture, Aquaculture; Browdy, C.L., Jory, D.E., Eds.; World Aquaculture Society, 2001; pp. 199–218. Available online: https://ag.arizona.edu/azaqua/tilapia/tilapia_shrimp/moriarty.PDF (accessed on 27 August 2022).
- De La Vega, E.; Degnan, B.; Hall, M.R.; Cowley, J.A.; Wilson, K.J. Quantitative real-time RT-PCR demonstrates that handling stress can lead to rapid increases of gill-associated virus (GAV) infection levels in Penaeus monodon. Dis. Aquat. Org. 2004, 59, 195–203.
- de la Vega, E.; Hall, M.R.; Degnan, B.M.; Wilson, K.J. Short-term hyperthermic treatment of Penaeus monodon increases expression of heat shock protein 70 (HSP70) and reduces replication of gill associated virus (GAV). Aquaculture 2006, 253, 82–90.
- Vidal, O.M.; Granja, C.B.; Aranguren, F.; Brock, J.A.; Salazar, M. A Profound Effect of Hyperthermia on Survival of Litopenaeus vannamei Juveniles Infected with White Spot Syndrome Virus. J. World Aquac. Soc. 2001, 32, 364–372.
- Ellis, R.J.; Van der Vies, S.M. Molecular chaperones. Annu. Rev. Biochem. 1991, 60, 321–347.
- Tomanek, L.; Somero, G. Evolutionary and acclimation-induced variation in the heat-shock responses of congeneric marine snails (genus Tegula) from different thermal habitats: Implications for limits of thermotolerance and biogeography. J. Exp. Biol. 1999, 202, 2925–2936.
- Tomanek, L. Variation in the heat shock response and its implication for predicting the effect of global climate change on species’ biogeographical distribution ranges and metabolic costs. J. Exp. Biol. 2010, 213, 971–979.
- Xu, Q.; Qin, Y. Molecular cloning of heat shock protein 60 (PtHSP60) from Portunus trituberculatus and its expression response to salinity stress. Cell Stress Chaperones 2012, 17, 589–601.
- Baruah, K.; Ranjan, J.; Sorgeloos, P.; MacRae, T.H.; Bossier, P. Priming the prophenoloxidase system of Artemia franciscana by heat shock proteins protects against Vibrio campbellii challenge. Fish Shellfish Immunol. 2011, 31, 134–141.
- Zhu, Y.; Zhu, G.; Guo, Q.; Zhu, Z.; Wang, C.; Liu, Z. A Comparative Proteomic Analysis of Pinellia ternata Leaves Exposed to Heat Stress. Int. J. Mol. Sci. 2013, 14, 20614–20634.
- Sung, Y.Y.; Dhaene, T.; Defoirdt, T.; Boon, N.; MacRae, T.H.; Sorgeloos, P.; Bossier, P. Ingestion of bacteria overproducing DnaK attenuates Vibrio infection of Artemia franciscana larvae. Cell Stress Chaperones 2009, 14, 603–609.
- Jakob, U.; Muse, W.; Eser, M.; Bardwell, J.C.A. Chaperone Activity with a Redox Switch. Cell 1999, 96, 341–352.
- Kumsta, C.; Jakob, U. Redox-Regulated Chaperones. Biochemistry 2009, 48, 4666–4676.
- Sharma, S.; Chakraborty, K.; Müller, B.K.; Astola, N.; Tang, Y.-C.; Lamb, D.C.; Hayer-Hartl, M.; Hartl, F.U. Monitoring Protein Conformation along the Pathway of Chaperonin-Assisted Folding. Cell 2008, 133, 142–153.
- Walter, S.; Buchner, J. Molecular chaperones—Cellular machines for protein folding. Angew. Chem. Int. Ed. 2002, 41, 1098–1113.
- Mayer, M.P.; Bukau, B. Hsp70 chaperones: Cellular functions and molecular mechanism. Cell. Mol. Life Sci. 2005, 62, 670.
- Flaherty, K.M.; McKay, D.B.; Kabsch, W.; Holmes, K.C. Similarity of the three-dimensional structures of actin and the ATPase fragment of a 70-kDa heat shock cognate protein. Proc. Natl. Acad. Sci. USA 1991, 88, 5041–5045.
- Lo, J.; Hayashi, M.; Woo-Kim, S.; Tian, B.; Huang, J.; Fearns, C.; Takayama, S.; Zapata, J.M.; Yang, Y.; Lee, J. Tid1, a co-chaperone of the heat shock 70 protein and the mammalian counterpart of the Drosophila tumor suppressor l(2)tid, is critical for early embryonic development and cell survival. Mol. Cell. Biol. 2004, 24, 2226–2236.
- Flaherty, K.M.; DeLuca-Flaherty, C.; McKay, D.B. Three-dimensional structure of the ATPase fragment of a 70K heat-shock cognate protein. Nature 1990, 346, 623–628.
- Kiang, J.G. Heat Shock Protein 70 kDa Molecular Biology, Biochemistry, and Physiology. Pharmacol. Ther. 1998, 80, 183–201.
- Kregel, K.C. Heat shock proteins: Modifying factors in physiological stress responses and acquired thermotolerance. J. Appl. Physiol. 2002, 92, 2177–2186.
- Cottin, D.; Shillito, B.; Chertemps, T.; Thatje, S.; Léger, N.; Ravaux, J. Comparison of heat-shock responses between the hydrothermal vent shrimp Rimicaris exoculata and the related coastal shrimp Palaemonetes varians. J. Exp. Mar. Biol. Ecol. 2010, 393, 9–16.
- Cottin, D.; Roussel, D.; Foucreau, N.; Hervant, F.; Piscart, C. Disentangling the effects of local and regional factors on the thermal tolerance of freshwater crustaceans. Die Nat. 2012, 99, 259–264.
- Yost, H.; Lindquist, S. RNA splicing is interrupted by heat shock and is rescued by heat shock protein synthesis. Cell 1986, 45, 185–193.
- Huang, L.-H.; Wang, C.-Z.; Kang, L. Cloning and expression of five heat shock protein genes in relation to cold hardening and development in the leafminer, Liriomyza sativa. J. Insect Physiol. 2009, 55, 279–285.
- Yang, J.; Mu, Y.; Dong, S.; Jiang, Q.; Yang, J. Changes in the expression of four heat shock proteins during the aging process in Brachionus calyciflorus (rotifera). Cell Stress Chaperones 2013, 19, 33–52.
- Hartl, F.U.; Martin, J. Molecular chaperones in cellular protein folding. Bioessays 1995, 16, 689–692.
- Bukau, B.; Horwich, A.L. The Hsp70 and Hsp60 Chaperone Machines. Cell 1998, 92, 351–366.
- Hartl, F.U.; Hayer-Hartl, M. Molecular Chaperones in the Cytosol: From Nascent Chain to Folded Protein. Science 2002, 295, 1852–1858.
- Beckmann, R.P.; Mizzen, L.E.; Welch, W.J. Interaction of Hsp 70 with Newly Synthesized Proteins: Implications for Protein Folding and Assembly. Science 1990, 248, 850–854.
- Palleros, D.R.; Welch, W.J.; Fink, A.L. Interaction of hsp70 with unfolded proteins: Effects of temperature and nucleotides on the kinetics of binding. Proc. Natl. Acad. Sci. USA 1991, 88, 5719–5723.
- Sadis, S.; Hightower, L.E. Unfolded proteins stimulate molecular chaperone Hsc70 ATPase by accelerating ADP/ATP exchange. Biochemistry 1992, 31, 9406–9412.
- Frydman, J.; Nimmesgern, E.; Ohtsuka, K.; Hartl, F.U. Folding of nascent polypeptide chains in a high molecular mass assembly with molecular chaperones. Nature 1994, 370, 111–117.
- Gething, M.-J.; Sambrook, J. Protein folding in the cell. Nature 1992, 335, 33–45.
- Chen, Z.; Zhou, T.; Wu, X.; Hong, Y.; Fan, Z.; Li, H. Influence of cytoplasmic heat shock protein 70 on viral infection of Nicotiana benthamiana. Mol. Plant Pathol. 2008, 9, 809–817.
- Park, H.; Lee, J.; Huh, S.; Seo, J.; Choi, E. Hsp72 functions as a natural inhibitory protein of c-Jun N-terminal kinase. EMBO J. 2001, 20, 446–456.
- Dong, Y.; Dong, S. Induced thermotolerance and expression of heat shock protein 70 in sea cucumber Apostichopus japonicus. Fish. Sci. 2008, 74, 573–578.
- Wu, G.; Harris, M.K.; Guo, J.-Y.; Wan, F.-H. Response of multiple generations of beet armyworm, Spodoptera exigua (Hübner), feeding on transgenic Bt cotton. J. Appl. Èntomol. 2009, 133, 90–100.
- Tsan, M.-F.; Gao, B. Heat shock protein and innate immunity. Cell. Mol. Immunol. 2004, 1, 274–279.
- Jolesch, A.; Elmer, K.; Bendz, H.; Issels, R.D.; Noessner, E. Hsp70, a messenger from hyperthermia for the immune system. Eur. J. Cell Biol. 2012, 91, 48–52.
- Welch, W.J.; Feramisco, J.R. Purification of the major mammalian heat shock proteins. J. Biol. Chem. 1982, 257, 14949–14959.
- Aligue, R.; Akhavan-Niak, H.; Russell, P. A role for Hsp90 in cell cycle control: Wee1 tyrosine kinase activity requires inter-action with Hsp90. EMBO J. 1994, 13, 6099–6106.
- Jakob, U.; Lilie, H.; Meyer, I.; Buchner, J. Transient interaction of Hsp90 with early unfolding intermediates of citrate synthase: Implications for heat shock in vivo. J. Biol. Chem. 1995, 270, 7288–7294.
- Pearl, L.H.; Prodromou, C. Structure and Mechanism of the Hsp90 Molecular Chaperone Machinery. Annu. Rev. Biochem. 2006, 75, 271–294.
- Reddy, P.S.; Thirulogachandar, V.; Vaishnavi, C.; Aakrati, A.; Sopory, S.K.; Reddy, M.K. Molecular characterization and expression of a gene encoding cytosolic Hsp90 from Pennisetum glaucum and its role in abiotic stress adaptation. Gene 2011, 474, 29–38.
- Fu, D.; Chen, J.; Zhang, Y.; Yu, Z. Cloning and expression of a heat shock protein (HSP) 90 gene in the haemocytes of Crassostrea hongkongensis under osmotic stress and bacterial challenge. Fish Shellfish Immunol. 2011, 31, 118–125.
- Xu, D.; Sun, L.; Liu, S.; Zhang, L.; Yang, H. Polymorphisms of heat shock protein 90 (Hsp90) in the sea cucumber Apostichopus japonicus and their association with heat-resistance. Fish Shellfish Immunol. 2014, 41, 428–436.
- Zhao, H.; Yang, H.; Zhao, H.; Chen, M.; Wang, T. The molecular characterization and expression of heat shock protein 90 (Hsp90) and 26 (Hsp26) cDNAs in sea cucumber (Apostichopus japonicus). Cell Stress Chaperones 2011, 16, 481–493.
- Zhu, J.-Y.; Wu, G.-X.; Ye, G.-Y.; Hu, C. Heat shock protein genes (hsp20, hsp75 and hsp90) from Pieris rapae: Molecular cloning and transcription in response to parasitization by Pteromalus puparum. Insect Sci. 2012, 20, 183–193.
- Quintana, F.J.; Cohen, I.R. Heat Shock Proteins as Endogenous Adjuvants in Sterile and Septic Inflammation. J. Immunol. 2005, 175, 2777–2782.
- Vabulas, R.M.; Ahmad-Nejad, P.; da Costa, C.; Miethke, T.; Kirschning, C.J.; Häcker, H.; Wagner, H. Endocytosed HSP60s Use Toll-like Receptor 2 (TLR2) and TLR4 to Activate the Toll/Interleukin-1 Receptor Signaling Pathway in Innate Immune Cells. J. Biol. Chem. 2001, 276, 31332–31339.
- Choresh, O.; Ron, E.; Loya, Y. The 60-kDa Heat Shock Protein (HSP60) of the Sea Anemone Anemonia viridis: A Potential Early Warning System for Environmental Changes. Mar. Biotechnol. 2001, 3, 501–508.
- Choresh, O.; Loya, Y.; Müller, W.E.; Wiedenmann, J.; Azem, A. The mitochondrial 60-kDa heat shock protein in marine invertebrates: Biochemical purification and molecular characterization. Cell Stress Chaperones 2004, 9, 38–48.
- Clayton, M.E.; Steinmann, R.; Fent, K. Different expression patterns of heat shock proteins hsp 60 and hsp 70 in zebra mussels (Dreissena polymorpha) exposed to copper and tributyltin. Aquat. Toxicol. 2000, 47, 213–226.
- Zhou, J.; Wang, W.-N.; He, W.-Y.; Zheng, Y.; Wang, L.; Xin, Y.; Liu, Y.; Wang, A.-L. Expression of HSP60 and HSP70 in white shrimp, Litopenaeus vannamei in response to bacterial challenge. J. Invertebr. Pathol. 2010, 103, 170–178.
- Huang, W.-J.; Leu, J.-H.; Tsau, M.-T.; Chen, J.-C.; Chen, L.-L. Differential expression of LvHSP60 in shrimp in response to environmental stress. Fish Shellfish Immunol. 2011, 30, 576–582.
- Sun, Y.; Macrae, T.H. Small heat shock proteins: Molecular structure and chaperone function. Cell. Mol. Life Sci. 2005, 62, 2460–2476.
- Mchaourab, H.S.; Godar, J.A.; Stewart, P.L. Structure and Mechanism of Protein Stability Sensors: Chaperone Activity of Small Heat Shock Proteins. Biochemistry 2009, 48, 3828–3837.
- Laganowsky, A.; Benesch, J.; Landau, M.; Ding, L.; Sawaya, M.; Cascio, D.; Huang, Q.; Robinson, C.; Horwitz, J.; Eisenberg, D. Crystal structures of truncated alphaA and alphaB crystallins reveal structural mechanisms of polydispersity important for eye lens function. Protein Sci. 2010, 19, 1031–1043.
- Hilario, E.; Martin, F.J.M.; Bertolini, M.C.; Fan, L. Crystal Structures of Xanthomonas Small Heat Shock Protein Provide a Structural Basis for an Active Molecular Chaperone Oligomer. J. Mol. Biol. 2011, 408, 74–86.
- Waters, E.R. The evolution, function, structure, and expression of the plant sHSPs. J. Exp. Bot. 2013, 64, 391–403.
- Kriehuber, T.; Rattei, T.; Weinmaier, T.; Bepperling, A.; Haslbeck, M.; Buchner, J. Independent evolution of the core domain and its flanking sequences in small heat shock proteins. FASEB J. 2010, 24, 3633–3642.
- Van Montfort, R.L.; Basha, E.; Friedrich, K.L.; Slingsby, C.; Vierling, E. Crystal structure and assembly of a eukaryotic small heat shock protein. Nat. Genet. 2001, 8, 1025–1030.
- Horwitz, J. Alpha-crystallin. Exp. Eye Res. 2003, 76, 145–153.
- Haslbeck, M.; Franzmann, T.; Weinfurtner, D.; Buchner, J. Some like it hot: The structure and function of small heat-shock proteins. Nat. Struct. Mol. Biol. 2005, 12, 842–846.
- Mogk, A.; Deuerling, E.; Vorderwülbecke, S.; Vierling, E.; Bukau, B. Small heat shock proteins, ClpB and the DnaK system form a functional triade in reversing protein aggregation. Mol. Microbiol. 2003, 50, 585–595.
- Lee, G.J.; Roseman, A.M.; Saibil, H.R.; Vierling, E. A small heat shock protein stably binds heat-denatured model substrates and can maintain a substrate in a folding-competent state. EMBO J. 1997, 16, 659–671.
- Cashikar, A.G.; Duennwald, M.; Lindquist, S.L. A Chaperone Pathway in Protein Disaggregation: Hsp26 alters the nature of protein aggregates to facilitate reactivation by Hsp104s. J. Biol. Chem. 2005, 280, 23869–23875.
- Liberek, K.; Lewandowska, A.; Ziętkiewicz, S. Chaperones in control of protein disaggregation. EMBO J. 2008, 27, 328–335.
- Jackson, S.A.; Clegg, J.S. Ontogeny of low molecular weight stress protein p26 during early embryogenesis of the brine shrimp, Artemia franciscana. Dev. Growth Differ. 1996, 38, 153–160.
- Clegg, J.S. Stress-related proteins compared in diapause and in activated, anoxic encysted embryos of the animal extremophile, Artemia franciscana. J. Insect Physiol. 2011, 57, 660–664.
- King, A.M.; Macrae, T.H. The Small Heat Shock Protein p26 Aids Development of Encysting Artemia Embryos, Prevents Spontaneous Diapause Termination and Protects against Stress. PLoS ONE 2012, 7, e43723.
- King, A.M.; Toxopeus, J.; MacRae, T.H. Functional differentiation of small heat shock proteins in diapause-destined Artemia embryos. FEBS J. 2013, 280, 4761–4772.
- Chen, T.; Villeneuve, T.S.; Garant, K.A.; Amons, R.; MacRae, T.H. Functional characterization of artemin, a ferritin homolog synthesized in Artemia embryos during encystment and diapause. FEBS J. 2007, 274, 1093–1101.
- Hu, Y.; Bojikova-Fournier, S.; King, A.M.; MacRae, T.H. The structural stability and chaperone activity of artemin, a ferritin homologue from diapause-destined Artemia embryos, depend on different cysteine residues. Cell Stress Chaperones 2010, 16, 133–141.
- King, A.M.; Toxopeus, J.; MacRae, T.H. Artemin, a Diapause-Specific Chaperone, Contributes to the Stress Tolerance of Artemia franciscana Cysts and Influences Their Release from Females. J. Exp. Biol. 2014, 217, 1719–1724.
- Macrae, T.H. Stress tolerance during diapause and quiescence of the brine shrimp, Artemia. Cell Stress Chaperones 2015, 21, 9–18.
- Dai, L.; Chen, D.-F.; Liu, Y.-L.; Zhao, Y.; Yang, F.; Yang, J.-S.; Yang, W.-J. Extracellular Matrix Peptides of Artemia Cyst Shell Participate in Protecting Encysted Embryos from Extreme Environments. PLoS ONE 2011, 6, e20187.
- Wu, B.J.; Kingston, R.E.; Morimoto, R.I. Human HSP70 promoter contains at least two distinct regulatory domains. Proc. Natl. Acad. Sci. USA 1986, 83, 629–633.
- Köhler, H.R.; Triebskorn, R.; Stöcker, W.; Kloetzel, P.-M.; Alberti, G. The 70 kD heat shock protein (hsp 70) in soil invertebrates: A possible tool for monitoring environmental toxicants. Arch. Environ. Contam. Toxicol. 1992, 22, 334–338.
- Viswanathan, C.; Khanna-Chopra, R. Heat shock proteins-Role in thermotolerance of crop plants. Curr. Sci. 1996, 71, 275–284.
- Morimoto, R.I. Cells in Stress: Transcriptional Activation of Heat Shock Genes. Science 1993, 259, 1409–1410.
- Amin, J.; Ananthan, J.; Voellmy, R. Key features of heat shock regulatory elements. Mol. Cell. Biol. 1988, 8, 3761–3769.
- Grossman, A.D.; Erickson, J.W.; Gross, C.A. The htpR gene product of E. coli is a sigma factor for heat-shock promoters. Cell 1984, 38, 383–390.
- Sorger, P.K.; Pelham, H.R. Yeast heat shock factor is an essential DNA-binding protein that exhibits temperature-dependent phosphorylation. Cell 1988, 54, 855–864.
- Kong, X.-C.; Zhang, D.; Qian, C.; Liu, G.-T.; Bao, X.-Q. FLZ, a novel HSP27 and HSP70 inducer, protects SH-SY5Y cells from apoptosis caused by MPP+. Brain Res. 2011, 1383, 99–107.
- Fang, H.; Wu, Y.; Huang, X.; Wang, W.; Ang, B.; Cao, X.; Wan, T. Toll-like Receptor 4 (TLR4) Is Essential for Hsp70-like Protein 1 (HSP70L1) to Activate Dendritic Cells and Induce Th1 Response. J. Biol. Chem. 2011, 286, 30393–30400.
- Triantafilou, M.; Triantafilou, K. Heat-shock protein 70 and heat-shock protein 90 associate with Toll-like receptor 4 in response to bacterial lipopolysaccharide. Biochem. Soc. Trans. 2004, 32, 636–639.
- Sato, S.; Fujita, N.; Tsuruo, T. Modulation of Akt kinase activity by binding to Hsp90. Proc. Natl. Acad. Sci. USA 2000, 97, 10832–10837.
- Lin, X.; Söderhäll, I. Crustacean hematopoiesis and the astakine cytokines. Blood 2011, 117, 6417–6424.
- Maningas, M.B.B.; Kondo, H.; Hirono, I.; Saito-Taki, T.; Aoki, T. Essential function of transglutaminase and clotting protein in shrimp immunity. Mol. Immunol. 2008, 45, 1269–1275.
- Fagutao, F.F.; Maningas, M.B.B.; Kondo, H.; Aoki, T.; Hirono, I. Transglutaminase regulates immune-related genes in shrimp. Fish Shellfish Immunol. 2012, 32, 711–715.
- Zhu, Y.-T.; Li, D.; Zhang, X.; Li, X.-J.; Li, W.-W.; Wang, Q. Role of transglutaminase in immune defense against bacterial pathogens via regulation of antimicrobial peptides. Dev. Comp. Immunol. 2016, 55, 39–50.
- Yu, Z.; Geng, Y.; Huang, A.; Wang, K.; Huang, X.; Chen, D.; Ou, Y.; Wang, J. Molecular characterization of a p38 mitogen-activated protein kinase gene from Scylla paramamosain and its expression profiles during pathogenic challenge. J. Invertebr. Pathol. 2017, 144, 32–36.
- He, S.; Qian, Z.; Yang, J.; Wang, X.; Mi, X.; Liu, Y.; Hou, F.; Liu, Q.; Liu, X. Molecular characterization of a p38 MAPK from Litopenaeus vannamei and its expression during the molt cycle and following pathogen infection. Dev. Comp. Immunol. 2013, 41, 217–221.
- Yan, H.; Zhang, S.; Li, C.-Z.; Chen, Y.-H.; Chen, Y.-G.; Weng, S.-P.; He, J.-G. Molecular characterization and function of a p38 MAPK gene from Litopenaeus vannamei. Fish Shellfish Immunol. 2013, 34, 1421–1431.
- Sanders, B.M. Stress Proteins in Aquatic Organisms: An Environmental Perspective. Crit. Rev. Toxicol. 1993, 23, 49–75.
- Feder, M.E.; Hofmann, G.E. Heat-Shock Proteins, Molecular Chaperones, and The Stress Response: Evolutionary and Ecological Physiology. Annu. Rev. Physiol. 1999, 61, 243–282.
- Huang, A.-M.; Geng, Y.; Wang, K.-Y.; Zeng, F.; Liu, Q.; Wang, Y.; Sun, Y.; Liu, X.-X.; Zhou, Y. Molecular Cloning and Expression Analysis of Heat Shock Protein 90 (Hsp90) of the Mud Crab, Scylla Paramamosain. J. Agric. Sci. 2013, 5, 1.
- Junprung, W.; Norouzitallab, P.; De Vos, S.; Tassanakajon, A.; Viet, D.N.; Van Stappen, G.; Bossier, P. Sequence and expression analysis of HSP70 family genes in Artemia franciscana. Sci. Rep. 2019, 9, 8391.
- Ravi, V.; Kubofcik, J.; Bandopathyaya, S.; Geetha, M.; Narayanan, R.; Nutman, T.; Kaliraj, P. Wuchereria bancrofti: Cloning and characterization of heat shock protein 70 from the human lymphatic filarial parasite. Exp. Parasitol. 2004, 106, 1–10.
- Luan, W.; Li, F.; Zhang, J.; Wen, R.; Li, Y.; Xiang, J. Identification of a novel inducible cytosolic Hsp70 gene in Chinese shrimp Fenneropenaeus chinensis and comparison of its expression with the cognate Hsc70 under different stresses. Cell Stress Chaperones 2009, 15, 83–93.
- Gbotsyo, Y.A. The Effect of Cold Stress on Heat Shock Proteins in Nauplii (larvae) of the Brine Shrimp, Artemia franciscana. Honours Thesis, Saint Mary’s University, Halifax, NS, Canada, 2017; pp. 1–39.
- Chapple, J.; Smerdon, G.R.; Berry, R.; Hawkins, A.J. Seasonal changes in stress-70 protein levels reflect thermal tolerance in the marine bivalve Mytilus edulis L. J. Exp. Mar. Biol. Ecol. 1998, 229, 53–68.
- Browne, R.; Bowen, S. Taxonomy and population genetics of Artemia. In Artemia Biology; Browne, R.A., Sorgeloos, P., Trotman, C.N.A., Eds.; CRC Press: Boca Raton, FL, USA, 1991; pp. 221–235.
- Triantaphyllidis, G.; Abatzopoulos, T.; Sorgeloos, P. Review of the biogeography of the genus Artemia (Crustacea, Anostraca). J. Biogeogr. 1998, 25, 213–226.
- Sorgeloos, P.; Bossuyt, E.; Laviña, E.; Baeza-Mesa, M.; Persoone, G. Decapsulation of Artemia cysts: A simple technique for the improvement of the use of brine shrimp in aquaculture. Aquaculture 1977, 12, 311–315.
- Kumar, V.; Das, B.K.; Swain, H.S.; Chowdhury, H.; Roy, S.; Bera, A.K.; Das, R.; Parida, S.N.; Dhar, S.; Jana, A.K.; et al. Outbreak of Ichthyophthirius multifiliis associated with Aeromonas hydrophila in Pangasianodon hypophthalmus: The role of turmeric oil in enhancing immunity and inducing resistance against co-infection. Front. Immunol. 2022, 13, 956478.
- Clegg, J.S.; Drinkwater, L.E.; Sorgeloos, P. The Metabolic Status of Diapause Embryos of Artemia franciscana (SFB). Physiol. Zool. 1996, 69, 49–66.
- Tran, P.T.N.; Kumar, V.; Bossier, P. Do acute hepatopancreatic necrosis disease-causing PirABVP toxins aggravate vibriosis? Emerg. Microbes Infect. 2020, 9, 1919–1932.
- Criel, G.R.J.; Macrae, T.H. Artemia morphology and structure. In Artemia: Basic and Applied Biology. Biology of Aquatic Organisms; Abatzopoulos, T.J., Beardmore, J.A., Clegg, J.S., Sorgeloos, P., Eds.; Springer: Dordrecht, The Netherlands, 2002; pp. 1–37. ISBN 978-94-017-0791-6.
- Van Stappen, G. Zoogeography. In Artemia: Basic and Applied Biology. Biology of Aquatic Organisms; Abatzopoulos, T.J., Beardmore, J.A., Clegg, J.S., Sorgeloos, P., Eds.; Springer: Dordrecht, The Netherlands, 2002; pp. 171–224.
- Robbins, H.M.; Van Stappen, G.; Sorgeloos, P.; Sung, Y.Y.; MacRae, T.H.; Bossier, P. Diapause termination and development of encysted Artemia embryos: Roles for nitric oxide and hydrogen peroxide. J. Exp. Biol. 2010, 213, 1464–1470.
- Kumar, V.; De Bels, L.; Couck, L.; Baruah, K.; Bossier, P.; Broeck, W.V.D. PirABVP Toxin Binds to Epithelial Cells of the Digestive Tract and Produce Pathognomonic AHPND Lesions in Germ-Free Brine Shrimp. Toxins 2019, 11, 717.
- Han, B.; Kaur, V.I.; Baruah, K.; Nguyen, V.D.; Bossier, P. High doses of sodium ascorbate act as a prooxidant and protect gnotobiotic brine shrimp larvae (Artemia franciscana) against Vibrio harveyi infection coinciding with heat shock protein 70 activation. Dev. Comp. Immunol. 2018, 92, 69–76.
- Kumar, V.; Nguyen, D.V.; Baruah, K.; Bossier, P. Probing the mechanism of VPAHPND extracellular proteins toxicity purified from Vibrio parahaemolyticus AHPND strain in germ-free Artemia test system. Aquaculture 2019, 504, 414–419.
- Kumar, V.; Baruah, K.; Bossier, P. Bamboo powder protects gnotobiotically-grown brine shrimp against AHPND-causing Vibrio parahaemolyticus strains by cessation of PirABVP toxin secretion. Aquaculture 2021, 539, 736624.
- De Vos, S.; Bossier, P.; Vuylsteke, M. Genomic Tools and Sex Determination in the Extremophile Brine Shrimp Artemia franciscana; Ghent University: Ghent, Belgium, 2014.
- Defoirdt, T.; Crab, R.; Wood, T.K.; Sorgeloos, P.; Verstraete, W.; Bossier, P. Quorum Sensing-Disrupting Brominated Furanones Protect the Gnotobiotic Brine Shrimp Artemia franciscana from Pathogenic Vibrio harveyi, Vibrio campbellii, and Vibrio parahaemolyticus Isolates. Appl. Environ. Microbiol. 2006, 72, 6419–6423.
- Crab, R.; Lambert, A.; Defoirdt, T.; Bossier, P.; Verstraete, W. The application of bioflocs technology to protect brine shrimp (Artemia franciscana) from pathogenic Vibrio harveyi. J. Appl. Microbiol. 2010, 109, 1643–1649.
- Miller, D.; McLennan, A.G. The heat shock response of the cryptobiotic brine Shrimp Artemia—I. A comparison of the thermotolerance of cysts and larvae. J. Therm. Biol. 1988, 13, 119–123.
- Seebaugh, D.R.; Wallace, W. Importance of metal-binding proteins in the partitioning of Cd and Zn as trophically available metal (TAM) in the brine shrimp Artemia franciscana. Mar. Ecol. Prog. Ser. 2004, 272, 215–230.
- Marques, A.; Ollevier, F.; Verstraete, W.; Sorgeloos, P.; Bossier, P. Gnotobiotically grown aquatic animals: Opportunities to investigate host-microbe interactions. J. Appl. Microbiol. 2006, 100, 903–918.
- El-Magsodi, M.O.; Bossier, P.; Sorgeloos, P.; Van Stappen, G. Effect of Light Colour, Timing, and Duration of Light Exposure on the Hatchability of Artemia Spp. (Branchiopoda: Anostraca). Eggs. J. Crustac. Biol. 2016, 36, 515–524.
- Clegg, J.S.; Jackson, S.A.; Van Hoa, N.; Sorgeloos, P. Thermal resistance, developmental rate and heat shock proteins in Artemia franciscana, from San Francisco Bay and southern Vietnam. J. Exp. Mar. Biol. Ecol. 2000, 252, 85–96.
- Kumar, V.; Bossier, P. Importance of plant—Derived compounds and/or natural products in aquaculture. Aquafeed 2018, 10, 28–31.
- Kumar, V.; Roy, S. Aquaculture Drugs: Sources, Active Ingredients, Pharmaceutic Preparations and Methods of Administration. J. Aquac. Res. Dev. 2017, 8, 510.
- Kumar, V.; Bossier, P. Novel plant-based compounds could be useful in protecting shrimp species against AHPND Vibrio parahaemolyticus. J. Inland Fish. Soc. India 2019, 51, 3–5.
- Kumar, V.; Wille, M.; Lourenço, T.M.; Bossier, P. Biofloc-Based Enhanced Survival of Litopenaeus vannamei Upon AHPND-Causing Vibrio parahaemolyticus Challenge Is Partially Mediated by Reduced Expression of Its Virulence Genes. Front. Microbiol. 2020, 11, 1270.
- Baruah, K.; Norouzitallab, P.; Phong, H.P.P.D.; Smagghe, G.; Bossier, P. Enhanced resistance against Vibrio harveyi infection by carvacrol and its association with the induction of heat shock protein 72 in gnotobiotic Artemia franciscana. Cell Stress Chaperones 2017, 22, 377–387.
- Frankenberg, M.; Jackson, S.; Clegg, J. The heat shock response of adult Artemia franciscana. J. Therm. Biol. 2000, 25, 481–490.
- Wandinger, S.K.; Richter, K.; Buchner, J. The Hsp90 Chaperone Machinery. J. Biol. Chem. 2008, 283, 18473–18477.
- Vos, M.; Hageman, J.; Carra, S.; Kampinga, H.H. Structural and Functional Diversities between Members of the Human HSPB, HSPH, HSPA, and DNAJ Chaperone Families. Biochemistry 2008, 47, 7001–7011.
- Kumar, V.; Roy, S.; Behera, B.K.; Swain, H.S.; Das, B.K. Biofloc Microbiome With Bioremediation and Health Benefits. Front. Microbiol. 2021, 12, 3499.
- Roy, S.; Baruah, K.; Bossier, P.; Vanrompay, D.; Norouzitallab, P. Induction of transgenerational innate immune memory against Vibrio infections in a brine shrimp (Artemia franciscana) model. Aquaculture 2022, 557, 738309.
- Roy, S.; Kumar, V.; Behera, B.K.; Parhi, J.; Mohapatra, S.; Chakraborty, T.; Das, B.K. CRISPR/Cas Genome Edit-ing-Can It Become a Game Changer in Future Fisheries Sector? Front. Mar. Sci. 2022, 9, 924475.
- Meimaridou, E.; Gooljar, S.B.; Chapple, J.P. From hatching to dispatching: The multiple cellular roles of the Hsp70 molecular chaperone machinery. J. Mol. Endocrinol. 2008, 42, 1–9.
- Vabulas, R.M.; Raychaudhuri, S.; Hayer-Hartl, M.; Hartl, F.U. Protein Folding in the Cytoplasm and the Heat Shock Response. Cold Spring Harb. Perspect. Biol. 2010, 2, a004390.
- Hartl, F.U.; Hayer-Hartl, M. Converging concepts of protein folding in vitro and in vivo. Nat. Struct. Mol. Biol. 2009, 16, 574–581.
- Yam, A.Y.; Xia, Y.; Lin, H.-T.J.; Burlingame, A.; Gerstein, M.; Frydman, J. Defining the TRiC/CCT interactome links chaperonin function to stabilization of newly made proteins with complex topologies. Nat. Struct. Mol. Biol. 2008, 15, 1255–1262.
- Shi, J.; Fu, M.; Zhao, C.; Zhou, F.; Yang, Q.; Qiu, L. Characterization and function analysis of Hsp60 and Hsp10 under different acute stresses in black tiger shrimp, Penaeus monodon. Cell Stress Chaperones 2015, 21, 295–312.
- Sung, Y.Y.; Ashame, M.F.; Chen, S.; MacRae, T.H.; Sorgeloos, P.; Bossier, P. Feeding Artemia franciscana (Kellogg) larvae with bacterial heat shock protein, protects from Vibrio campbellii infection. J. Fish Dis. 2009, 32, 675–685.
- Kumar, V.; Roy, S.; Behera, B.K.; Das, B.K. RNA Interference and Its Potential Applications in Aquatic Animal Health Management. In Biotechnological Advances in Aquaculture Health Management; Springer: Singapore, 2021; pp. 25–41.
- Kumar, V.; Roy, S.; Behera, B.K.; Das, B.K. Disease Diagnostic Tools for Health Management in Aquaculture. In Advances in Fisheries Biotechnology; Springer: Singapore, 2021; pp. 363–382.
- Roy, S.; Kumar, V.; Behera, B.K.; Das, B.K. Epigenetics: Perspectives and Potential in Aquaculture. In Advances in Fisheries Biotechnology; Springer: Singapore, 2021; pp. 133–150.
More
Information
Subjects:
Fisheries
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.8K
Revisions:
4 times
(View History)
Update Date:
29 Nov 2022
Table of Contents



Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No