1. Regulation of the Heat Shock Protein Response
Heat shock proteins (Hsps) are evolutionarily ancient and extremely conserved proteins found in almost all living organisms, ranging from archaebacteria, prokaryotes, and eukaryotes. Hsps play an essential role in regulating cellular metabolism in stressful conditions. These proteins are present in several intracellular locations, e.g., in the nucleus, mitochondria, endoplasmic reticulum (ER), and chloroplast and cytosol of eukaryotes [
31,
32,
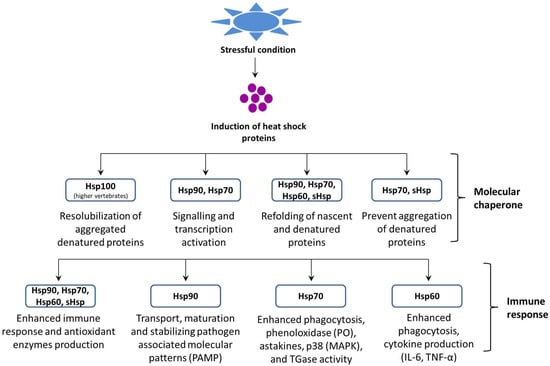
33]. In general, Hsps are classified into several families based on their function, molecular mass, and sequence homology (i.e., Hsp60, Hsp70, Hsp90, and sHsps) (
Figure 1) and also, they can be grouped according to their nearest size family (e.g., Hsp84, Hsp85, and Hsp86 in the Hsp90 kDa family) [
34].
Figure 1. Role of heat shock proteins (Hsps) in proteostasis and the host immune response.
Hsps plays a key role in cellular processes that occur during and after exposure to oxidative stress that is caused by hazardous environmental and/or microbial agents. As a result, the normal intracellular reducing environment is compromised, which leads to oxidation and aggregation of key proteins and DNA, ultimately resulting in cellular dysfunction. Due to their versatile functions, Hsps can intervene following oxidative stress at several levels [
18,
19,
35]. Firstly, some Hsps, mainly the Hsp70 family members, play a crucial role in protein sorting and quality control via selecting and directing abnormal proteins to the proteasome or lysosomes for degradation; thus, Hsps aid the clearance of damaged proteins [
36]. In some cases, where misfolded proteins need to be rescued, the same machinery facilitates the correct folding of damaged proteins. Moreover, the Hsp families, such as Hsp27, Hsp70, and Hsp90, can negatively regulate apoptosis via the binding and inhibiting of members of the apoptotic cascade. Some Hsps have immuno-enhancing actions (for details, see the section on Hsps and immunity). Among the different Hsp families, Hsp70 is the largest and most highly conserved of the stress protein families. At least 121 proteins have been characterized within this family, and cross-hybridization occurs across various species, such as in mammals, fish, and mollusks [
37,
38,
39].
After the discovery of heat shock proteins, it was clear that the heat shock protein response requires a specific transcription factor [
40,
41,
42,
43,
44]. In crustaceans, a few reports have suggested that the heat shock transcription factor plays a significant role in determining the heat shock response and also in tolerance against stressful conditions. Tan and Macrae [
45] reported that the heat shock factor 1 (Hsf1) transcription factor induces stress tolerance (drying) in diapausing brine shrimp cysts and is responsible for the improved growth and survival of
A. franciscana [
45]. In another study, Sornchuer et al. [
46] demonstrated that Hsf1 has an important role in the thermal stress response and regulates the transcription of heat shock proteins and immune-related genes in
P. monodon [
46]. However, we still need to carry out detailed investigations and obtain more information in order to characterize the mechanism of heat shock response regulation in crustaceans. Moreover, in eukaryotes, the heat shock response is well documented, and this study shows that the induction of Hsp transcription is mediated by a pre-existing transcription factor, i.e., the heat shock factor or the heat shock transcription factor (HSF) [
47]. The HSF upon activation binds to the heat shock element (HSE) at the promoter region (5’ upstream end) of the Hsp gene and induces the transcription of Hsp [
48]. The binding motif of the HSE is composed of nGAAn blocks (5 bp) in alternate orientation, and for the stable binding of the HSF and the HSE, at least three units are required [
49]. In prokaryotes (e.g.,
E. coli), the σ32 regulatory protein is responsible for Hsp expression [
50]. Additionally, as an alternate subunit of the bacterial RNA polymerase, σ32 replaces the σ70 normal regulatory proteins during heat stress [
51]. HSF1 and σ32 share basic mechanistic properties, however their structure or sequence are not related; further, it has been found that protein homeostasis disturbance results in the activation of hsf1 and σ32 [
52].
HSF genes have been reported from yeast (
Saccharomyces cerevisiae), plants (
Arabidopsis sp.), fruit flies (
Drosophila sp.), chicken (
Gallus sp.),
A. franciscana, and
P. monodon [
53,
54,
55,
56,
57]. Sequence comparison of the HSF gene from different species showed that DNA binding and the oligomerization domain is strongly conserved. The HSF contains two highly conserved regions, i.e., a ~100 amino acids NH2-terminal DNA binding domain and an adjacent trimerization domain having 3 hydrophobic heptad repeats, leu zippers [
33]. The activation of the HSF involves cellular factors as the intermediary sensors that regulate the activity of the HSF during non-stressful conditions [
58]. In animals, the HSF is maintained as a monomeric form through transient interaction with Hsps. However, during stressful conditions, the HSF was released and formed trimers with the HSE, which resulted in the induction of Hsps. Subsequently, Hsps bind with denatured/misfolded protein aggregates and maintain the homeostatic condition in the cells [
59,
60,
61,
62,
63].
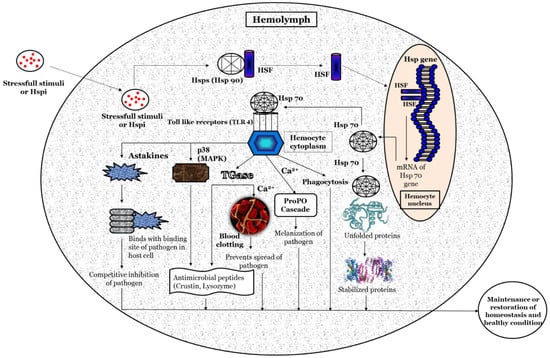
Based on previous studies, a hypothetical illustration of the possible methods involved in the activation of the heat shock protein response, and its potential role in maintaining homeostasis and host health, is depicted in
Figure 2. Briefly, the stressful stimuli, heat-shock-protein-inducing (Hspi) conditions (e.g., environmental, pathological, and physiological) [
64,
65], or compounds (e.g., plant-based, natural polymers, etc.) [
66,
67] develop oxidative status inside the host. Subsequently, it facilitates the phosphorylation and nuclear translocation of the HSF (in the native form present in the non-phosphorylated form attached with heat shock proteins such as Hsp90) [
68] where (inside the hemocyte nucleus) the HSF binds with the HSE at the promoter region (5’ upstream end) of the Hsp gene and induces the transcription of the Hsp gene and production of Hsps [
69,
70]. The Hsp (e.g., Hsp70) functions as a non-covalently molecular chaperone bind with a hydrophobic exposed segment of unfolded proteins [
71,
72]. Further, this prevents the aggregation of inappropriate or unfolded proteins; inhibits the misfolding of the polypeptides; transports immature polypeptides to target organelle for the purposes of final packaging and repair, as well as denaturation or degradation of misfolded proteins through proteasomes or lysosomes (proteolysis), which cannot be repaired [
73]; and, also, maintains protein homeostasis. The extracellular Hsp also functions as a chaperokine and binds with Toll-like receptors (e.g., TLR-4), which are expressed on hemocytes that lead to the maturation and activation of hemocytes [
74,
75,
76]. The activated hemocytes induce the production of astakines that binds with the binding site of a pathogen in the host cell and decrease the chance of the pathogen attaching to its host cells [
77,
78,
79]. The hemocytes (activated by Hsps) also induce the expression of transglutaminase (Tgase) [
80], which form a blood clot in the presence of Ca
2+ and prevents the spread of pathogens and increases the expression of antimicrobial peptides (e.g., crustin, lysozyme, etc.) that has bactericidal activity against Gram-positive and Gram-negative bacteria [
81,
82,
83,
84]. The p38 (mitogen-activated protein kinase, MAPK) activated by hemocytes, increases the expression of antimicrobial peptides (e.g., crustin, lysozyme, etc.) and has a critical role in defense against bacterial and viral infection [
85,
86,
87,
88,
89]. Hemocytes activate the ProPO cascade in the presence of Ca
2+, leading to melanization and further killing of the pathogens [
90,
91,
92,
93].
Figure 2. A schematic hypothetical illustration of the possible mechanism of action for Heat shock proteins (Hsps) in crustaceans.
2. Factors Modulate Heat Shock Protein Response
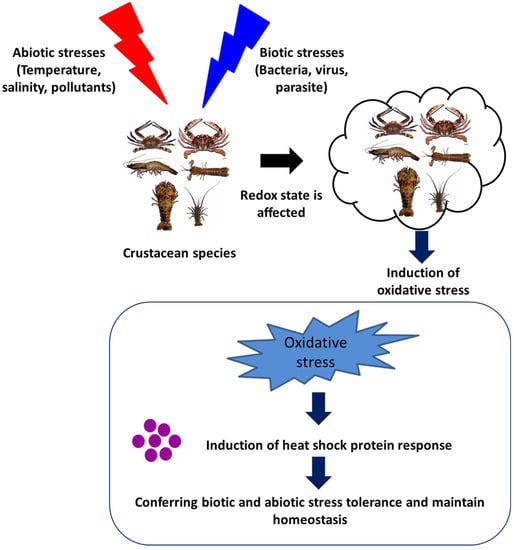
The induction of Hsps in response to cellular stressors was initially considered a short-term functional response, with a range of essential housekeeping and cytoprotective functions. These stressors, in general, induce protein damage and increase the susceptibility of host animals further to subsequent stressful conditions. However, accumulating pieces of evidence, over the past few decades, have suggested that Hsps play a significant role in the regulation of the immune response in invertebrates (Figure 3 and Table 1). The heat shock protein expressions influenced by either abiotic or biotic stresses are summarized in the section below.
Figure 3. Effect of abiotic and biotic stresses on host health and heat shock protein response.
2.1. Environmental or Abiotic Stresses
The physiological status of crustaceans is greatly influenced by their environmental conditions. A slight variation in environmental parameters creates a stressful condition that attenuates the immune system and increases the susceptibility of animals to microbial infection.
2.1.1. Temperature
Temperature is considered an important abiotic stressor, as a slight change in water temperature can affect the body physiology and health of crustaceans [
105,
106,
107]. Interestingly, heat shock proteins are amongst the most significant proteins that are induced by hypo and hyperthermia and their role in protection against thermal stress has been well documented [
108,
109,
110,
111,
112]. Among the Hsp multigenic family, few proteins are expressed at extremely low levels under normal conditions, while the transcription of most Hsps increased significantly in response to stresses, e.g., stress-inducible proteins (Hsp70). However, Hsps that are expressed constitutively under normal conditions, and may be upregulated during stress conditions, are generally known as heat shock cognate proteins, e.g., Hsc70 [
113,
114,
115,
116]. Hsps play a central role in thermotolerance by promoting growth at moderately high temperatures and protecting the organism from mortality at extremely high temperatures [
117]. In general, Hsps are induced in both hyperthermia and hypothermia conditions. For example, an increase in the water temperature has been found to induce the production of Hsp70 in
Ferropenaeus chinensis, Chinese white shrimp [
118],
Gammarus pulex, freshwater crustacean [
119], and
A. franciscana, [
120]. Moreover, increased Hsp (Hsp90 and Hsp40) expression was also reported during 6 h of cold shock at 1 and 6 °C in
A. franciscana [
121]. Hsp70 has been demonstrated to play an important role in protecting cells from damage in
S. paramamosain in response to thermal stress (this was shown in an increase in 11 °C from normal growth temperature) [
122]. In addition, the adult
A. franciscana, when exposed to sub-lethal heat shock (37 °C for 30 min), induces the transcription of Hsp70, Hsp67, and Hsc70, resulting in an improved tolerance of brine shrimp to high temperatures [
123]. Interestingly, marine invertebrates are very sensitive to high temperatures and there are a myriad of reports suggesting that Hsp70 is upregulated in response to heat stress [
124,
125,
126]. These studies indicate that Hsps improve thermal tolerance in crustaceans and provide protection in both hyperthermia and hypothermia conditions.
2.1.2. Salinity
The concentration of dissolved inorganic salt concentrations or the salinity in the water is reported to affect the osmoregulation of crustaceans and induce cellular damage, including a deleterious effect on the folding and transformation of polypeptides [
127,
128,
129,
130]. Moreover, the osmotic-stress-induced Hsp production plays a crucial role in the maintenance of biological processes, as well as the protection of crustaceans against stressful conditions [
131,
132,
133,
134,
135]. The study by Yang et al. [
79] reported that high salinity stress increased the expression of Hsp70 and that this could lead to enhanced resistance in
S. paramamosain against changes in water salinity. The possible mechanism behind the protective action of Hsps against salinity stress is that the osmotic change increases the metabolism rate and enhances the stress response resulting in increased Hsp production [
136]. The increased Hsp response subsequently enhances the immune response, including lysozyme, phenoloxidase, and peroxide activity and provides rapid protection against osmotic stress until the organic osmolytes are fully accumulated [
137,
138]. Water salinity was reported as an important factor for the purposes of natural growth of crustaceans; further, studies on
P. trituberculatus have shown that variable salinity significantly influences larval development [
139]. Xu and Qin [
61] found that Hsp60 has an important role in both the cellular and humoral stress response of the swimming crab,
P. trituberculatus, and that these responses regulate the salinity stress via an intrinsic pathway. Further, these responses also play an essential role in protecting the swimming crab against salinity stress. In addition, a few reports have suggested that an increase in the expression of Hsp70 enhances immune response and confers protection to
A. franciscana against hypersalinity stress [
140].
2.1.3. Environmental Pollutants
The environmental pollutants induced heat shock proteins (Hsps) expression in crustaceans are the most frequently studied in the literature [
141,
142,
143]. The study of heat shock proteins in invertebrates started in the 1990s, and the first observation conducted on Hsps was made by Köhler et al. [
144]. The study showed that the exposure of three diplopods (
Tachypodoiulus niger,
Cylindroiulus punctatus, and
Glomeris marginata), one isopod (
Oniscus asellus) and two slugs (
Arion ater and
Deroceras reticulatum) to heavy metals/molluscicides resulted in the increased expression of Hsp70 [
145].
Studies on the effect of environmental pollutants on crustaceans demonstrated that the Hsp gene expression is induced by several chemical stresses. For example, this can be found in: nonylphenol (NP) (used in the polymer industry); bisphenol A diglycidyl ether (BPA) (intermediate in the production of polycarbonate and epoxy resins) [
146]; 17α-ethynyl estradiol (EE) (synthetic estrogen) [
147]; bis(2-ethylhexyl) phthalate (DEHP) (plasticizer in polymer products); endosulfan (ES) (organochlorine insecticide); chloropyriphos (CP) (organophosphorus insecticide); paraquat dichloride (PQ) (oxygen radical generating herbicide); Cadmium (Cd); lead (Pb) and potassium dichromate (Cr) (heavy metals); and benzo[a] pyrene (BaP) (polycyclic aromatic hydrocarbon) [
148]. Moreover, for instance, the sublethal concentration of endosulfan has been reported to enhance the synthesis of Hsp70 and Hsp90 in monsoon river prawn, i.e.,
M. malcolmsonii and
P. monodon [
149,
150,
151,
152]. In another study, the mixture of environmental pollutant chemicals has been reported to modulate the physiological, as well as immunity and survival responses in crustaceans. For instance, Park and Kwak [
89] demonstrated that the application of bisphenol A (BPA) and 4-nonlphenol (NP)—an endocrine disrupting chemical (EDCs)—at different concentrations and at different time intervals (12, 24, 48, and 96 h) induce the expression of Hsp90; further, when exposed to BPA and NP, the marine crab,
Charybdis japonica, has a significantly increased survival [
54]. Additionally, chemical stress was reported to induce the production of Hsps, which helps in maintaining the homeostasis and structural integrity of cells [
153,
154].
2.2. Biotic Stresses
The heat shock proteins that are induced by biotic stresses play a very crucial role in protein folding, immune enhancement, and cross-protection against infectious diseases [
155,
156,
157]. Although, there are several reports that demonstrate that Hsps are easily induced by abiotic stresses including heat, salinity, etc. [
158], very little information is available on crustaceans’ Hsp response against biotic stresses, including bacteria, parasites, and viruses. Some of the recent research findings have suggested that members of the Hsp70 family have been identified in crustaceans, which are involved in the response to biotic stresses, mainly bacteria, parasites, and viruses [
159,
160]. Zhou et al. [
119] demonstrated that
L. vannamei when challenged with
V. alginolyticus (Gram-negative) and
S. aureus (Gram-positive) bacteria have a significantly increased expression of
L. vannamei Hsp60 (LvHSP60) and Hsp70 (LvHSP70) gene in the gills, hepatopancreas, and hemocytes. In another study, temporal transcription of LvHSP70, following the white spot syndrome virus (WSSV) challenge, has been reported to induce an anti-WSSV innate immune response in
L. vannamei [
161].
In the swimming crab,
Portunus trituberculatus, transcription of the
P. trituberculatus, the Hsp70 (PtHsp70) gene was shown to increase very rapidly in response to the bacterial challenge with
V. alginolyticus [
162]. Similar findings were reported by Yang et al. [
19], i.e., the fact that there was an increased transcription of Hsp70 in hemocytes of
S. paramamosain after the
V. alginolyticus challenge and were involved in generating cross-protection in the mud crab. Additionally, Hsp90, which plays a crucial role in protein biosynthesis, signal transduction, and immune responses, was also shown to induce protective immunity in crustaceans. Huang et al. [
20] analyzed the role of Hsp90 in
S. paramamosain in response to microbial infection, and the results showed that the transcription of SpHSP90 was upregulated in mud crabs after being challenged with
Staphylococcus aureus, white spot syndrome virus (wssv), and
V. harveyi [
20].
Hsps exert their physiological effect via assisting in the formation of new polypeptides as well as in the protecting and maintaining of the host cell polypeptides and naïve proteins from denaturation during microbial infection [
163,
164,
165]. In crustaceans, the host immune response against microbial infection [
133] is often associated with reactive oxygen species (ROS) production [
166]. It has been demonstrated that in
L. vannamei, after bacterial challenge, the subsequent induction of the host immune response leads to an increase in ROS levels [
167]. While microbial infection-induced ROS mostly has antimicrobial activity, the production of ROS can result in the denaturation of proteins (proteotoxicity) in the host cell itself; as such, in this condition, the induced Hsps display a cytoprotective role and act as chaperone proteins in order to maintain the protein homeostasis and preserve cellular structures [
168].
3. An A. franciscana Model System to Investigate the Role of Hsps in Crustaceans
The
A. franciscana is a small branchiopod crustacean that is highly osmotolerant and reported from several harsh environmental conditions worldwide [
169,
170,
171,
172]. They live in an environment of severe hypersalinity, high levels of ultraviolet radiation, fluctuating oxygen concentration, and extreme temperature [
173,
174]. The oviparous development in
A. franciscana leads to the production of hard-shell covering diapause cysts, which are composed of stress-tolerant metabolically inactive embryos stalled at gastrulation and that which can remain in stasis for several years [
175,
176,
177,
178]. However, when the diapause cysts were immersed under appropriate conditions in seawater with aeration and temperatures, the hard-shell raptures and cysts develop, releasing swimming larvae within 24 h (
Figure 4) [
178,
179,
180].
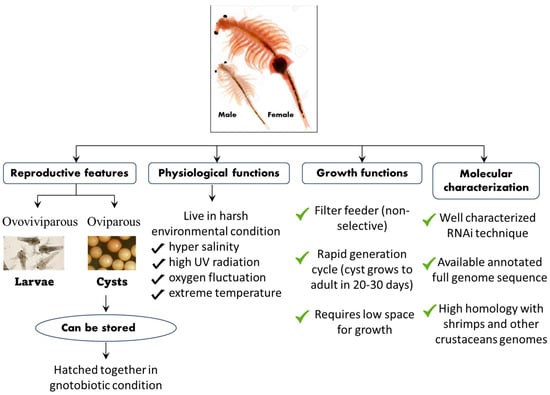
Figure 4. Schematic representation of the life cycle, physiological, growth, and molecular features of the A. franciscana model system.
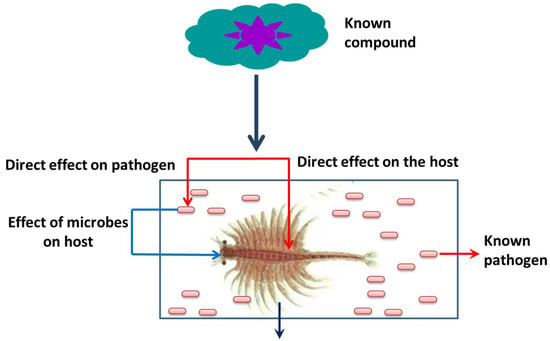
Apart from its interesting life history,
A. franciscana are non-selective filter feeders (which can be grown in a wide range of feed resources), have a rapid generation cycle (the cyst grows to adult in 20–30 days), require very low space for growth (hence have a relatively smaller cost to culture), and developmental stages are well characterized. This aspect of their stages being well characterized, for instance, is shown in the fact that the
A. franciscana produces encysted gastrulae cysts during oviparous development, while ovoviviparous development provides live larvae in both sexual and asexual (parthenogenetic) stages. Additionally, gnotobiotic (germ-free) culture conditions (allowing full control over the host-associated microbial communities) and advanced molecular techniques including qPCR and RNAi are well established in
A. franciscana, which makes this species an exceptional model organism that can be used in order to investigate the host–pathogen relationship and to study the biological activity of protective compounds [
181,
182,
183,
184,
185,
186,
187,
188,
189,
190]. In addition, cysts of
A. franciscana can be stored for a couple of years in the fridge and, after terminating the diapause stage, the cysts from different generations can be used and hatched all together, simultaneously. This excellent facility of storage and hatching, permits one to perform experiments on demand, which can help to avoid or minimize environmental influences. Above all, the genome sequence of
A. franciscana showed that it shares a very high homology with shrimps and other crustaceans’ genomes (
Figure 5). Therefore, there is a high possibility that the outcome of studies based on
A. franciscana would provide a fundamental basis to understand the host–pathogen interactions in other commercially important shrimp species.
Figure 5. Advantage of a gnotobiotic (germ free) A. franciscana model system.
The induction of Hsp production inside the host in order to control diseases in aquaculture was investigated using the model organism, brine shrimp larvae [
191,
192]. In 1988, Miller and McLennan observed the presence of heat shock proteins in the early developmental stages of brine shrimp, i.e., in encysted gastrula embryos (cysts) and newly hatched nauplius larvae. They have reported that the larvae exhibited induced thermotolerance, which is associated with the synthesis and upregulation of heat shock proteins [
193]. The role of heat shock protein response in adult
A. franciscana, in response to high temperature (including LT50 determination, enhanced thermotolerance, and increased production of the Hsp70 family stress protein) was studied by Frankenberg et al. [
95]. Results demonstrated that Hsp70 family proteins (mainly Hsp67 and Hsc70) levels were significantly upregulated during sublethal heat shock (37 °C for 30 min). The
A. franciscana exposed to Zn-control/Cd-control treatment induced the expression of Hsp and increased the partitioning of Cd to the tropically available metal (TAM), which could result in the bio-enhancement of Cd trophic transfer to predators, which, in turn, leads to the suppression of Zn accumulation in
A. franciscana [
194].
Later, a gnotobiotic (germ-free) culture system was developed for brine shrimp and several studies have demonstrated that this provides a fully controlled and excellent host–pathogen environment and facilitates the determining of the effect of external stimuli on the host (
Figure 5) [
195]. The gnotobiotic system also avoids the interference generated by host-associated microorganisms as well as shifts in the composition of microbial diversity [
196,
197,
198,
199,
200]. The effect of non-lethal heat shock (NLHS) on host survival, immune response, and protection against stressful conditions were also studied by several researchers. Results showed that NLHS increased the transcription of Hsp70, which enhances the immune response and provides cross-protection from environmental, physiological, and microbial stress in
A. franciscana [
201,
202,
203,
204,
205,
206]. In another study, Sung et al. [
64] demonstrated that non-pathogenic CAG 629 and CAG 626
E. coli strains, when heat shocked and administered through feed to
Artemia larvae, resulted in enhanced Hsp70 expression (~two folds) and subsequent protection against
V. campbellii infection [
207].
This entry is adapted from the peer-reviewed paper 10.3390/life12111777