 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Karishma Mody | -- | 2945 | 2022-11-10 00:36:44 | | | |
| 2 | Rita Xu | Meta information modification | 2945 | 2022-11-10 02:46:03 | | |
Video Upload Options
Insects and ectoparasites are causes for major concern throughout the world due to their economic and welfare impacts on livestock agriculture. Current control measures involve chemicals such as acaricides which pose challenges like chemical resistance and longer withholding periods. To enable more sustainable agriculture practices, it is important to develop technologies that combine targeted effectiveness with minimal environmental footprint. RNA interference (RNAi) is a eukaryotic process in which transcript expression is reduced in a sequence-specific manner. This makes it a perfect tool for developing efficient and effective biological control against pests and pathogens. Double-stranded RNA (dsRNA) is the key trigger molecule for inducing RNAi; this concept is widely studied for development of RNA-based biopesticides as an alternative to chemical controls in crop protection for targeting pests and pathogens with accuracy and specificity.
1. Introduction
2. RNA-Interference (RNAi)
2.1. RNAi Mechanism
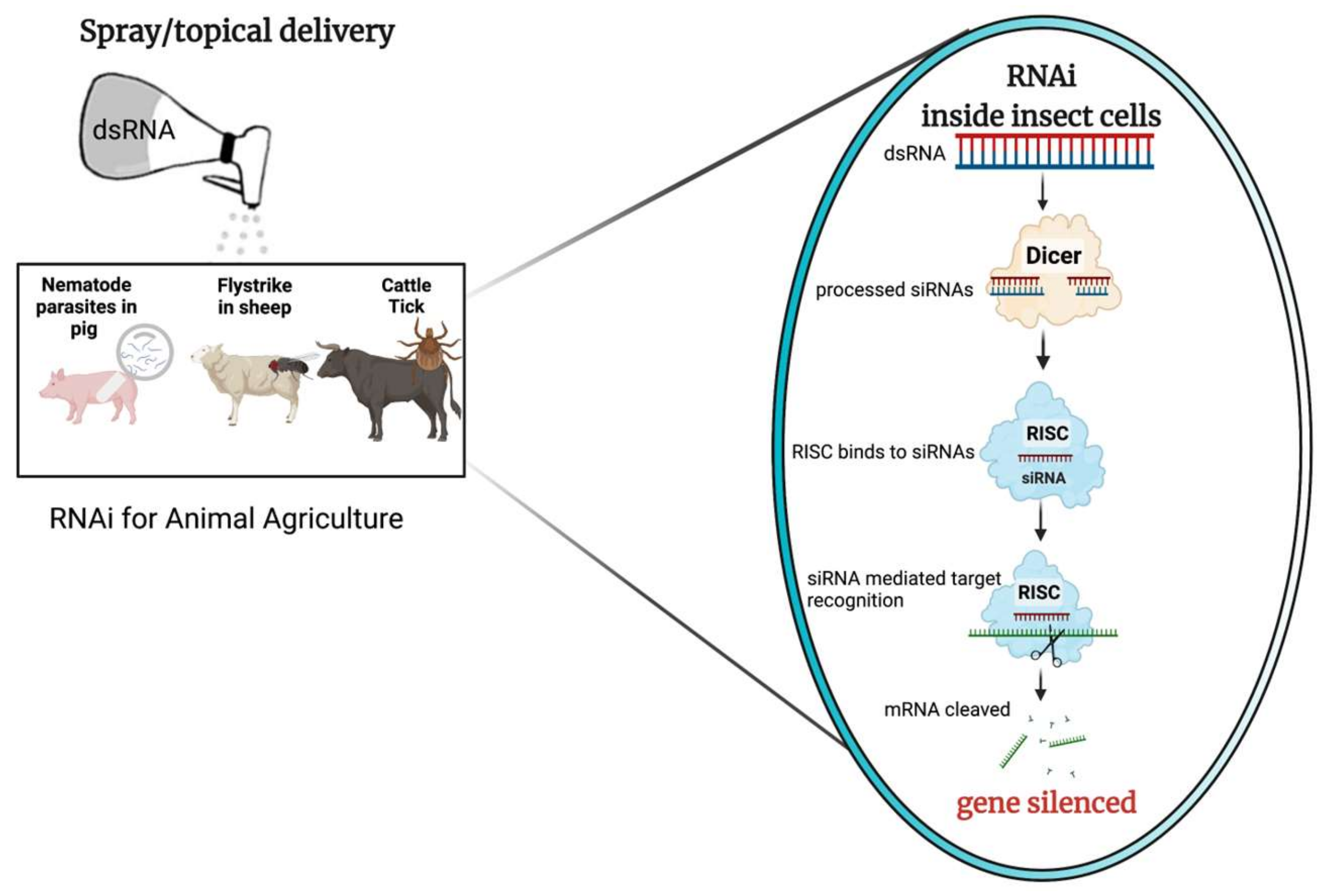
2.2. RNAi Targets
2.3. dsRNA Uptake Mechanism
3. Application of RNAi for Animal Health
| RNAi Target | Parasites | Delivery | Livestock | Efficiency | Ref |
|---|---|---|---|---|---|
| As-eft-1, As-eft-2, As-gmpr, As-tnc-1, As-rab-3, As-hb-1, As-unc-29, As-unc-38 |
Ascaris suum | Soaking | Pig | Different targets in adult A. suum were tested for gene knockdown. All targets were significantly silenced in different region of adult A. suum. Furthermore, all genes indicate they are susceptible to RNAi in adults A. suum, especially neuronal genes. | [39] |
| As-enol-1 | Ascaris suum | Soaking | Pig | Enolase gene expression was significantly silenced, and the mortality rate increased by 20.11% (p < 0.01) after soaking expressed sequence tag (EST) representing As-enol-1 dsRNA derived from A. suum for 72 h. | [34] |
| EST-06G09 | Ascaris suum | Soaking | Pig | The gene of EST 06G09 was silenced after 72 h soaking. The survival rate achieved highest peak at 48 h (20.37%, p < 0.01). The gene 06G09 has function in larval development. | [40] |
| TsSPIs | Trichinella spiralis | Soaking | Pig | TsSPI mRNA and protein expression levels in larvae decreased by 68.7% and 68.4%, respectively (p < 0.05). RNAi-mediated suppression of TsSPI expression in T. spiralis drastically decreased larval infectivity and survival inside the host. | [41] |
| LsalMS | Lepeophtheirus salmonis | Soaking | Salmon Lice | Myosuppressin (LsalMS) knockdown resulted in reduction in the amount of muscle, with skeletal and visceral muscles both showing anomalies in the lice. Additionally, LsalMS down-regulation also had an impact on feeding, spermatophore deposition, and moulting. | [42] |
| Ubiquitin-63E | Rhipicephalus microplus; | Injection | Cattle | All ticks treated with dsRNA did not have offspring. Ubiquitin-63E knockdown was confirmed by microarray and qRT-PCR. | [43] |
| HBP | Amblyomma mericanum | Injection | Cattle | dsRNA specific for histamine binding protein (HBP) transcript was injected into the haemolymph. Less expression of HBP mRNA was shown by molecular evidence, resulting in lower histamine binding ability. | [44] |
| Subolesin | Ornithodoros spp. | Injection | Cattle | By injecting dsRNA to silence the subolesin gene (Silencing 70–78%), oviposition was prevented (88.6% and 95.4%). | [45] |
| Fhteg1, Fhteg5, Fhteg8, | Fasciola hepatica | Soaking | Cattle | 92.9% transcript reduction of Fhteg1, 74.1% transcript reduction of Fhteg5. But no observable effect on phenotype. | [46] |
| SsGST-mu1 | Sarcoptes scabiei | Soaking | Sheep | Compared to controls, knockdown caused a 35% decrease in transcription of the target gene. | [47] |
| Hyaluronidase | Haemonchus contortus | Oral feeding | Sheep | A significant drop in worm burden and significant decrease in hyaluronidase activity in siRNA treated group. | [48] |
References
- FAO. The State of Food and Agriculture 2001; Food & Agriculture Organization: Rome, Italy, 2001.
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.D.; McRoberts, N.; Nelson, A.D. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439.
- Narladkar, B. Projected economic losses due to vector and vector-borne parasitic diseases in livestock of India and its significance in implementing the concept of integrated practices for vector management. Vet. World 2018, 11, 151.
- Worrall, E.; Hamid, A.; Mody, K.; Mitter, N.; Pappu, H. Nanotechnology for Plant Disease Management. Agronomy 2018, 8, 285.
- Chagnon, M.; Kreutzweiser, D.; Mitchell, E.A.D.; Morrissey, C.A.; Noome, D.A.; Van der Sluijs, J.P. Risks of large-scale use of systemic insecticides to ecosystem functioning and services. Environ. Sci. Pollut. Res. Int. 2014, 22, 119–134.
- Bai, G.; Shaner, G. Management and resistance in wheat and barley to Fusarium head blight. Annu. Rev. Phytopathol. 2004, 42, 135–161.
- Kim, Y.H.; Issa, M.S.; Cooper, A.M.; Zhu, K.Y. RNA interference: Applications and advances in insect toxicology and insect pest management. Pestic. Biochem. Physiol. 2015, 120, 109–117.
- Jain, R.G.; Robinson, K.E.; Fletcher, S.J.; Mitter, N. RNAi-based functional genomics in Hemiptera. Insects 2020, 11, 557.
- Schuster, S.; Miesen, P.; van Rij, R.P. Antiviral RNAi in insects and mammals: Parallels and differences. Viruses 2019, 11, 448.
- Chen, J.; Peng, Y.; Zhang, H.; Wang, K.; Zhao, C.; Zhu, G.; Reddy Palli, S.; Han, Z. Off-target effects of RNAi correlate with the mismatch rate between dsRNA and non-target mRNA. RNA Biol. 2021, 18, 1747–1759.
- Napoli, C.; Lemieux, C.; Jorgensen, R. Introduction of a chimeric chalcone synthase gene into petunia results in reversible co-suppression of homologous genes in trans. Plant Cell 1990, 2, 279–289.
- Sen, G.L.; Blau, H.M. A brief history of RNAi: The silence of the genes. FASEB J. 2006, 20, 1293–1299.
- Guo, S.; Kemphues, K.J. par-1, a gene required for establishing polarity in C. elegans embryos, encodes a putative Ser/Thr kinase that is asymmetrically distributed. Cell 1995, 81, 611–620.
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811.
- Wytinck, N.; Manchur, C.L.; Li, V.H.; Whyard, S.; Belmonte, M.F. dsRNA uptake in plant pests and pathogens: Insights into RNAi-based insect and fungal control technology. Plants 2020, 9, 1780.
- Bramlett, M.; Plaetinck, G.; Maienfisch, P. RNA-based biocontrols—A new paradigm in crop protection. Engineering 2020, 6, 522–527.
- Darlington, M.; Reinders, J.D.; Sethi, A.; Lu, A.L.; Ramaseshadri, P.; Fischer, J.R.; Boeckman, C.J.; Petrick, J.S.; Roper, J.M.; Narva, K.E. RNAi for Western Corn Rootworm Management: Lessons Learned, Challenges, and Future Directions. Insects 2022, 13, 57.
- Vogel, E.; Santos, D.; Mingels, L.; Verdonckt, T.-W.; Broeck, J.V. RNA interference in insects: Protecting beneficials and controlling pests. Front. Physiol. 2019, 9, 1912.
- Nunes, C.C.; Dean, R.A. Host-induced gene silencing: A tool for understanding fungal host interaction and for developing novel disease control strategies. Mol. Plant Pathol. 2012, 13, 519–529.
- You, L.; Zhang, F.; Huang, S.; Merchant, A.; Zhou, X.; Li, Z. Over-expression of rna interference (rnai) core machinery improves susceptibility to rnai in silkworm larvae. Insect Mol. Biol. 2020, 29, 353–362.
- Zotti, M.; Dos Santos, E.A.; Cagliari, D.; Christiaens, O.; Taning, C.N.T.; Smagghe, G. RNA interference technology in crop protection against arthropod pests, pathogens and nematodes. Pest Manag. Sci. 2018, 74, 1239–1250.
- Huvenne, H.; Smagghe, G. Mechanisms of dsRNA uptake in insects and potential of RNAi for pest control: A review. J. Insect Physiol. 2010, 56, 227–235.
- Saleh, M.-C.; van Rij, R.P.; Hekele, A.; Gillis, A.; Foley, E.; O’Farrell, P.H.; Andino, R. The endocytic pathway mediates cell entry of dsRNA to induce RNAi silencing. Nat. Cell Biol. 2006, 8, 793–802.
- Joga, M.R.; Zotti, M.J.; Smagghe, G.; Christiaens, O. RNAi Efficiency, Systemic Properties, and Novel Delivery Methods for Pest Insect Control: What We Know So Far. Front. Physiol. 2016, 7, 553.
- Fletcher, S.J.; Reeves, P.T.; Hoang, B.T.; Mitter, N. A Perspective on RNAi-Based Biopesticides. Front. Plant Sci. 2020, 11, 51.
- Luck, S.; Kreszies, T.; Strickert, M.; Schweizer, P.; Kuhlmann, M.; Douchkov, D. siRNA-Finder (si-Fi) Software for RNAi-Target Design and Off-Target Prediction. Front. Plant Sci. 2019, 10, 1023.
- Whyard, S.; Singh, A.D.; Wong, S. Ingested double-stranded RNAs can act as species-specific insecticides. Insect Biochem. Mol. Biol. 2009, 39, 824–832.
- Katoch, R.; Sethi, A.; Thakur, N.; Murdock, L.L. RNAi for insect control: Current perspective and future challenges. Appl. Biochem. Biotechnol. 2013, 171, 847–873.
- Feinberg, E.H.; Hunter, C.P. Transport of dsRNA into cells by the transmembrane protein SID-1. Science 2003, 301, 1545–1547.
- Okamura, K.; Chung, W.-J.; Ruby, J.G.; Guo, H.; Bartel, D.P.; Lai, E.C. The Drosophila hairpin RNA pathway generates endogenous short interfering RNAs. Nature 2008, 453, 803–806.
- Mourrain, P.; Béclin, C.; Elmayan, T.; Feuerbach, F.; Godon, C.; Morel, J.-B.; Jouette, D.; Lacombe, A.-M.; Nikic, S.; Picault, N. Arabidopsis SGS2 and SGS3 genes are required for posttranscriptional gene silencing and natural virus resistance. Cell 2000, 101, 533–542.
- Price, D.R.; Gatehouse, J.A. RNAi-mediated crop protection against insects. Trends Biotechnol. 2008, 26, 393–400.
- Alper, S.; McBride, S.J.; Lackford, B.; Freedman, J.H.; Schwartz, D.A. Specificity and complexity of the Caenorhabditis elegans innate immune response. Mol. Cell. Biol. 2007, 27, 5544–5553.
- Chen, N.; Xu, M.-J.; Nisbet, A.J.; Huang, C.-Q.; Lin, R.-Q.; Yuan, Z.-G.; Song, H.-Q.; Zhu, X.-Q. Ascaris suum: RNAi mediated silencing of enolase gene expression in infective larvae. Exp. Parasitol. 2011, 127, 142–146.
- Jex, A.R.; Liu, S.; Li, B.; Young, N.D.; Hall, R.S.; Li, Y.; Yang, L.; Zeng, N.; Xu, X.; Xiong, Z. Ascaris suum draft genome. Nature 2011, 479, 529–533.
- Peter, R.; Van den Bossche, P.; Penzhorn, B.L.; Sharp, B. Tick, fly, and mosquito control—Lessons from the past, solutions for the future. Vet. Parasitol. 2005, 132, 205–215.
- de la Fuente, J.; Estrada-Pena, A.; Venzal, J.M.; Kocan, K.M.; Sonenshine, D.E. Overview: Ticks as vectors of pathogens that cause disease in humans and animals. Front. Biosci. 2008, 13, 6938–6946.
- Fang, E.; Liu, X.; Li, M.; Zhang, Z.; Song, L.; Zhu, B.; Wu, X.; Liu, J.; Zhao, D.; Li, Y. Advances in COVID-19 mRNA vaccine development. Signal Transduct. Target. Ther. 2022, 7, 94.
- McCoy, C.J.; Warnock, N.D.; Atkinson, L.E.; Atcheson, E.; Martin, R.J.; Robertson, A.P.; Maule, A.G.; Marks, N.J.; Mousley, A. RNA interference in adult Ascaris suum–an opportunity for the development of a functional genomics platform that supports organism-, tissue-and cell-based biology in a nematode parasite. Int. J. Parasitol. 2015, 45, 673–678.
- Xu, M.; Chen, N.; Song, H.; Lin, R.; Huang, C.; Yuan, Z.; Zhu, X. RNAi-mediated silencing of a novel Ascaris suum gene expression in infective larvae. Parasitol. Res. 2010, 107, 1499–1503.
- Yi, N.; Yu, P.; Wu, L.; Liu, Z.; Guan, J.; Liu, C.; Liu, M.; Lu, Y. RNAi-mediated silencing of Trichinella spiralis serpin-type serine protease inhibitors results in a reduction in larval infectivity. Vet. Res. 2020, 51, 139.
- Komisarczuk, A.Z.; Kongshaug, H.; Li, M.; Nilsen, F. RNAi mediated myosuppressin deficiency affects muscle development and survival in the salmon louse (Lepeophtheirus salmonis). Sci. Rep. 2019, 9, 6944.
- Lew-Tabor, A.; Kurscheid, S.; Barrero, R.; Gondro, C.; Moolhuijzen, P.; Valle, M.R.; Morgan, J.; Covacin, C.; Bellgard, M. Gene expression evidence for off-target effects caused by RNA interference-mediated gene silencing of Ubiquitin-63E in the cattle tick Rhipicephalus microplus. Int. J. Parasitol. 2011, 41, 1001–1014.
- Aljamali, M.; Bior, A.; Sauer, J.; Essenberg, R. RNA interference in ticks: A study using histamine binding protein dsRNA in the female tick Amblyomma americanum. Insect Mol. Biol. 2003, 12, 299–305.
- Manzano-Román, R.; Díaz-Martín, V.; Oleaga, A.; Siles-Lucas, M.; Pérez-Sánchez, R. Subolesin/akirin orthologs from Ornithodoros spp. soft ticks: Cloning, RNAi gene silencing and protective effect of the recombinant proteins. Vet. Parasitol. 2012, 185, 248–259.
- McCusker, P.; Toet, H.; Rathinasamy, V.; Young, N.; Beddoe, T.; Anderson, G.; Dempster, R.; McVeigh, P.; McCammick, E.; Wells, D. Molecular characterisation and vaccine efficacy of two novel developmentally regulated surface tegument proteins of Fasciola hepatica. Vet. Parasitol. 2020, 286, 109244.
- Fernando, D.D.; Marr, E.J.; Zakrzewski, M.; Reynolds, S.L.; Burgess, S.T.; Fischer, K. Gene silencing by RNA interference in Sarcoptes scabiei: A molecular tool to identify novel therapeutic targets. Parasites Vectors 2017, 10, 289.
- Yang, X.; Khan, S.; Zhao, X.; Zhang, J.; Nisar, A.; Feng, X. Suppression of hyaluronidase reduces invasion and establishment of Haemonchus contortus larvae in sheep. Vet. Res. 2020, 51, 106.
- Cagliari, D.; Dias, N.P.; Galdeano, D.M.; Dos Santos, E.Á.; Smagghe, G.; Zotti, M.J. Management of pest insects and plant diseases by non-transformative RNAi. Front. Plant Sci. 2019, 10, 1319.

