2. RNA-Interference (RNAi)
RNAi is an endogenous, post-transcriptional gene regulation mechanism which has been identified in almost all eukaryotes; from plants, fungi, algae, protozoans, invertebrates, to vertebrates [
18,
19]. It was first reported by Napoli and Jorgensen in 1990 when they observed endogenous gene co-suppression while studying pigmented petunias [
20,
21]. In animals, RNAi was first documented in the nematode
Caenorhabditis elegans [
22,
23]. While the intracellular components of RNAi are similar across species, it was initially known as quelling in fungi, and post transcriptional gene silencing in plants [
24]. It is a highly conserved mechanism, which is highly sequence specific and selective in its activity [
25].
2.1. RNAi Mechanism
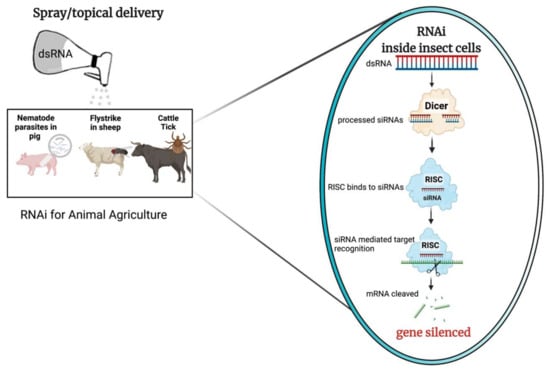
While a comprehensive review on RNAi mechanism discussing all the different facets of this technology is outside the scope of this review, the basic mechanism of RNAi when initiated via delivered exogenously applied dsRNA are well described and are illustrated in Figure 1.
Figure 1. Schematic example of RNA interference (RNAi) delivery via spray/topical application for animal agriculture; this image was made using BioRender. After application over the animal fur/surface, the dsRNA would enter the cells of the insect pest or pathogen. dsRNA is cleaved by dicer-2 into pre-siRNA duplexes, that are further processed into small-interfering RNA (siRNAs, 21–24 nt) effector molecules. The RNA-induced silencing complex (RISC) binds to the siRNAs and guides sequence-dependant degradation or translational inhibition of homologous mRNAs, which results in RNAi-mediated gene silencing.
RNAi-mediated post-transcriptional gene silencing is triggered by the processing of a dsRNA precursor into short single stranded RNA effector molecules [
18]. There are three RNAi pathways, depending on the RNA class of effector involved, small-interfering RNA (siRNA), piwi-interacting RNA (piRNA), and microRNA (miRNA) [
18,
25,
26,
27]. Of these, siRNA is considered to be the ‘classical’ pathway, with dsRNA being the trigger molecule for gene silencing in insects and other species [
27]. In the siRNA pathway, once taken up by cells dsRNA is cleaved by an endonuclease, Dicer, into pre-siRNA duplexes [
24]. The pre-siRNA is a 21–23 nucleotide (nt) long duplex with 2 nt overhangs at each of the 3′ termini. The pre-siRNA duplex is bound by the RNA-induced silencing complex (RISC), with one strand, the guide strand, being retained within the complex, while the complementary or passenger strand of the pre-siRNA duplex is degraded [
4,
28]. The guide siRNA strand with the Argonaut proteins (Ago) within the activated RISC, identify matching mRNA in a sequence dependent manner, resulting in suppression of translation or mRNA degradation [
29]. This results in loss of protein function which may lead to lethality or stunted growth of the target organism [
4,
27].
2.2. RNAi Targets
Target gene selection depends on the class of RNA effector molecule to be used. While both siRNA and piRNA are generated from long, complementary dsRNAs, miRNA is endogenous in nature, usually processed from stem-loops, and siRNA are exogenous and are directly generated from cleaved dsRNAs [
30]. The first step in developing an RNAi product is to identify potential target genes and design dsRNA targeting them. In 2010, a research study suggested that there are five factors which play a key role in influencing the efficiency of RNAi as biocontrol, namely, the concentration, nucleotide sequence, length of the dsRNA, effective period of the dsRNA induced silencing, and life stage of the target [
31].
The perfect RNAi target gene should be transcribed into an mRNA with a high turn overrate, that is translated into a protein with a short half-life, and is transcribed at all stages of life cycle, with the loss of function leading to mortality or severe impairment in the pest or pathogen of interest. In addition, the target transcript should be poorly conserved across species to maintain maximum specificity of mRNA for minimal environmental effects on non-target species [
27]. This is a challenging criterion to meet, as those genes that are essential to cell viability tend to be more conserved between species.
It has reported that the efficiency of dsRNA uptake is length dependent, requiring an optimal length and dosage to induce RNAi successfully [
32]. Studies have shown that dsRNA as short as 30 and 60-bp can induce 30% and 70% gene knockdown in the red flour beetle (
Tribolium castaneum), and the optimum length 200–520 bps for most insects [
31,
33]. This ideal dosage changes according to the organism’s susceptibility to RNAi and their potential for systemic RNAi, formulation for delivery, gene expression abundance, life and development stage for gene expression [
27]. Unfortunately, genes that often would lead to mortality if targeted by RNAi are highly conserved across nature, due to their evident importance for homeostasis [
27].
Various selection methods can be used to develop efficient RNAi targets to minimize side effects and enhance expected silencing. Since RNAi relies on high gene specificity to the selected target species based on sequence divergence, dsRNA needs to be screened and designed to be specific to a target gene or to target genes on a broad spectrum, closer to related species [
34]. To increase knockdown or have higher than the expected RNAi result, multiple targets can be selected for a single transcript to ensure variation within the target species is accounted for. A common application that can be used to evaluate off-target genes is the Basic Local Alignment Search Tool (BLAST), and it has been used to find contiguous matching sections of 17 nt or more in the genomes of interesting organisms [
34]. BLAST is a common approach for finding regions of identity, and for identifying the functionality of siRNA. Thereafter, a wide range of specialised programs can be used to design the sequences. These include siRNA-Finder (si-Fi), siDirect, dsCheck, and RNAi Designer by ThermoFisher [
35].
Amongst insect researchers it is widely accepted that several dsRNA should be screened as a combination as each gene is highly unique to the target gene and the insect species [
33]. Whyard et al. demonstrated species specific insecticides could be achieved by targeting the variable 3′-UTR regions of the
y-Tubulin transcript, allowing for a specific species knockdown between four closely related
Drosophila species [
36]. The results of this study clearly demonstrate that with sufficient sequence information and thorough interrogation of this available data, the development of highly specific RNAi based insecticides is highly feasible.
While the application of RNAi animal agriculture is still in its infancy, there are more dosage trials in plant agriculture, such as efficiency of dosage needed per acre regarding insect pests. Despite the fact that sufficient field experimentation is lacking, it is estimated that near 2–10 g of dsRNA is needed per acre of plants [
30]. As this depends on the organisms response efficiency and target gene, this number is open to high levels of variability [
30].
2.3. dsRNA Uptake Mechanism
The use of RNAi to protect plants and animals by suppressing essential gene function in pest species has been well documented in model invertebrates such as
Caenorhabditis elegans, and
Drosophila melanogaster [
37]. While most of the reported experiments involved injecting dsRNA into the target organism, this is not a viable delivery approach for in-field applications in animal production, thus creating the need for autonomous dsRNA uptake [
31,
37]. Studies have reported two dsRNA uptake mechanisms, the transmembrane channel-mediated uptake and an alternate endocytosis-mediated uptake [
37].
Transmembrane channel-mediated uptake mechanism has been best studied in
C. elegans, more specifically with worms carrying mutant
sid genes that lack systemic RNAi capacity [
31]. This pathway involves two proteins, SID-1 and SID-2, with changes being made to the protein molecule affecting other cells and their activity. SID-1 is a hypothesized transmembrane protein which passively transports dsRNA into the cells [
31,
38]. The role played by SID-2 has been hypothesized to help facilitate RNAi by coordinating with SID-1 either by modifying the molecule to activate it or binding to dsRNA for delivery or by inducing endocytosis [
31]. It has been noted that SID-1 is considered to be a ‘cell surface conduit’ for dsRNA uptake [
32]. Meanwhile, endocytosis-mediated mechanism was first studied in
D. melanogaster, since it had no
sid gene orthologues and the mechanism was later confirmed in
C. elegans [
31]. This is a cell-autonomous pathway wherein dsRNA is transported intracellularly via vesicles [
31,
38]. Drosophila S2 cells were able to take up dsRNA and initiate an RNAi response by receptor-mediated endocytosis uptake which is an active process involving vacuolar H
+ (V-H) ATPase [
31,
32]. This pathway is active, specific and is conserved in other animal cells [
32].
Some organisms can take up exogenous dsRNA and trigger an RNAi response in the entire animal systemically, like
C. elegans, while others cannot transmit this RNAi response in another cell [
32]. In
Drosophila, it has been confirmed that transposon inversion can activate RNAi, and the homologous gene silencing caused by it is similar to co-inhibition in plants [
39]. In a
C. elegans assay, it was found that the quelling defective protein, QDE-1, a necessary protein for RNAi process, is homologous to RNA-dependent RNA polymerase (RDRP), supporting RNAi process potential involved in RNA replication and regulation [
40]. Similarly, dsRNA injection into phloem of the plant can spread throughout the whole plant to initiate systemic RNAi [
41]. Interestingly, RNAi can also be induced by immersing nematodes in dsRNA-containing liquids or by feeding nematodes with
E. coli bacteria engineered to express dsRNA [
42]. The pathway for entry of dsRNA into the target cell determines RNAi efficiency and provides a starting point for the development of dsRNA delivery systems [
32].
3. Application of RNAi for Animal Health
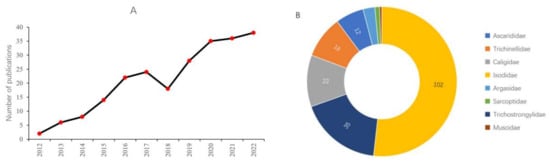
Over the past ten years, there has been a significant increase in published papers regarding application of RNAi to control in parasites affecting livestock, the data of which has been figuratively described in Figure 2A. This section highlights those RNAi studies which have been focused on the control of pests that affect pigs, sheep, and cattle pest.
Figure 2. Recent publications of RNAi based pest management associated with livestock. (A) Number of publications in the past 10 years based on a query-based PubMed search. (B) Recent quantitative distribution of RNAi studies in major livestock pest families. Based on PubMed search: More than 203 publications have been identified with queries “RNAi, gene silencing, specific species name, pest management, and specific livestock animals” by PubMed search.
RNAi technology has been described as a novel method for determining gene function and has provided an appealing approach to pest and disease management [
65]. There has been abundant transcriptomic/genomic resources available for
Ascaris suum, making it an experimental model among nematode parasites [
66]. Apart from studies focused on nematode parasite management in pigs, there were studies done with regard to tick control in cattle. Ticks are ectoparasites of both animals and humans and are a significant disease vector affecting cattle husbandry globally [
67,
68].
With regard to human health, large-scale production of mRNA has been achieved for commercial production with the development of mRNA vaccines for COVID in the recent years [
69]. Lipid nanoparticles (LNPs) which are composed of ionisable lipids, helper phospholipids, cholesterol, and PEGylated lipids are used to deliver mRNA vaccines developed by biopharmaceutical companies such as Moderna against COVID-19 and Zika viruses and BioNTech against COVID-19 spike proteins [
69]. Over the past few years, research in pest management has further extended towards other livestock pest species.
Table 1 enumerates some of the major experiments conducted in the past decade on the application of RNAi based pest control in the agricultural sector.
Table 1. A summary of the RNAi studies focused on the management of pest associated with livestock production.
The data from Table 1 demonstrates that RNAi has the potential to be successfully applied for parasite management and improve animal husbandry practices. RNAi technology has been described as a novel method for determining gene function and has provided an appealing disease management approach in parasitic nematodes. Out of the examples illustrated, the most promising results were obtained for nematode management, highlighting its potential for improving pig health. More than half of the studies focusing on tick control were on Ixodidae and Argasidae families, covering the following species, Amblyomma mericanum, Ixodes scapularis, Haemaphysalis longicornis, Rhipicephalus sanguineus, Dermacentor marginatus, and Boophilus microlpolu.
The field application of RNAi product for pest control In animal is still in an early stage. One specific research example is the application of RNAi against
Haemonchus contortus commonly known as barber’s pole worm in sheep. Researchers successfully used siRNA treatment for 8000 barber’s pole worms [
78]. They used a dosage of ~1 mg/10,000 larval to observe a RNAi effect. For this study, the quantity of siRNA needed for RNAi effect was ~1 mg per animal [
78]. In animal agriculture, each animal needs to be treated individually against pests and the exact quantity of RNA precursors required RNAi silencing can be very subjective depending on various factors like: (1) Size, animal types, animal density and living conditions of the farm. (2) Delivery system (nanoparticle/directly) and methods (injection/feeding). (3) Knockdown of genes with different function. (4) Species, stages of the pest. (5) Infection abilities associated with different pest and animal also need to be considered.
DsRNA has been delivered to the various tick species using different techniques, including microinjection, soaking, and feeding. As a major parasite of cattle, research studies focusing on RNAi strategy for tick species have achieved considerable gene knockdown or mortality increase [
67,
68]. Many parasites that affect livestock remain significant research subjects for flystrike in sheep, anaplasmosis in cattle, nematode infection in pigs and ectoparasites of salmon. While large-scale production of dsRNA has been expensive in the past, there has been a significant drop in production costs. This is due to the development of mass production systems, such as bacterial expression vectors for in vivo synthesis [
79]. Thus, the use of RNAi as biopesticide has an excellent potential as control strategy for these pests affecting livestock industries.