 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Julián Rafael Dib | + 3033 word(s) | 3033 | 2020-11-12 04:49:40 | | | |
| 2 | Catherine Yang | Meta information modification | 3033 | 2020-11-30 03:56:00 | | |
Video Upload Options
Killer yeast for the Biological Control of Postharvest Fungal Crop Diseases
1. Introduction
Postharvest fungal diseases of fruit and vegetables cause major crop losses ranging from 25% in industrialized up to 50% in developing countries [1]. Counteractive measures include chemical, physical, and biological approaches. Fungicides most frequently serve to combat fungal infestations of field crops as such chemically synthesized compounds are rather inexpensive, can be stored for long periods of time, and they are sufficiently effective. However, concern is continuously rising over their routine use because, besides threatening human health [2][3][4] and the environment [5], resistant fungal pathogenic biotypes concomitantly arose [6][7][8]. It is, thus, not surprising that a number of countries, such as the USA and those that form the European Union, promote a project called “Integrated Pest Management (IPM)” which aims at reducing or, whenever possible, completely replacing chemical pesticides [9][10][11][12][13][14].
2. Yeasts as Biocontrol Agents
Besides rather sophisticated mechanisms enabling yeasts to restrict or kill competitors (see the following chapter), there are some general characteristics qualifying these unicellular eukaryotes (in its broadest sense) for protection: they use nutrients rapidly and proliferate abundantly; they have the ability to colonize fruit surfaces displaying rather long immobilization times enduring even dry conditions; they produce extra-cellular polysaccharides enhancing their own survival, concomitantly restricting pathogens; and they sufficiently tolerate commonly applied pesticides, such as imazalil, tiabendazole, pyrimethanil or fludioxonil [15][16][17][18][19].
Though numerous reports regarding the use of yeasts as biological control agents exist, the number of commercial products is limited and, indeed, covers the potential market only at a small fraction. Multiple and time-consuming steps are most critical for product development and implementation, and that is why the involvement of a commercial company with expertise in upscaling, formulation, registration procedures, and the marketing network is a must to successfully meet above challenges [20].
Reviews about killer yeasts and their applications in medicine, industry and agriculture have been written over the years [21][22][23][24]. Here, we focus on representatives which actually function or presumably can act as candidates for antagonizing harmful –most commonly filamentous– fungi known to cause postharvest rotting of edible fruit and vegetables. After dealing with killer yeasts in general, we present an up-to-date compilation of the literature with respect to the aforesaid application which is schematically outlined in Figure 1.
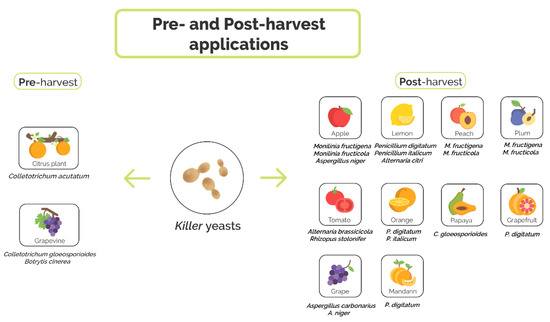
Figure 1. Applications of killer yeasts in agriculture to prevent fungal infections. Killer yeasts have been used for both pre- and postharvest treatments.
2.1. The Yeast Killer Phenotype
Killer substances are produced by pro- and eukaryotic cells [2][25][26][27][28]. They clearly improve the organisms’ capability to dominate a certain environmental niche by killing or inhibiting competing microorganisms. However, the killer phenotype is not always implemented solely for being advantageous for its host as the primary function rather often apparently ensures auto-selection of the extrachromosomal elements encoding the killer toxin (virus-like particles (VLPs) or virus-like elements (VLEs)) since it eliminates any cell that does not or no longer carries the cytoplasmic VLP or VLE [29]. Nevertheless, as a collateral action it is advantageous for the producing cell as it inhibits competing organisms.
The killer phenotype was first described for the baker’s yeast Saccharomyces cerevisiae by Bevan and Makower in the year 1963 [30]. They differentiated three phenotypes, i.e., killer, sensitive and neutral: a killer yeast is capable of producing toxins that cause the death of sensitive yeasts, a neutral yeast is not a toxin producer and is also not affected by the toxin. Woods and Bevan [31] realized the proteinaceous nature of the toxin, and they checked optimal conditions for its action: such as low pH, chemical composition of the growth medium, and the physiological state of the cells affecting the amount of the killing toxin. Initially, it was suggested that killer yeasts are lethal only to other yeasts, but further studies evidenced that they can be equally toxic to other fungi or even bacteria [32][33].
2.2. Genetics of the Killer Phenotype
First studies on the nature of the killer factor revealed the involvement of cytoplasmic genetic determinants [34]. Double stranded RNA (dsRNA) associated with VLPs was proposed and approved as the genetic basis [31][34][35][36][37][38][39]. Today, killer phenomena are known for a large number of yeasts [40][41]. Toxins cannot only be extra-chromosomally encoded but also chromosomally [23].
Killer toxins are often classified with respect to their cellular target. Essentially, there are four categories: (a) toxins capable of generating membrane pores followed by cellular lysis; (b) toxins displaying glucanase activity, eventually resulting in cell wall destruction and target cell lysis; (c) toxins disturbing nuclear functions by blocking replication; and (d) toxins targeting cellular RNAs such as specific tRNAs or rRNA (see also Figure 2).
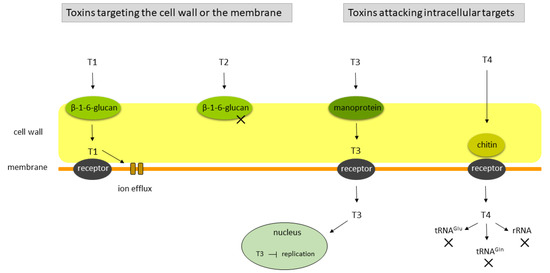
Figure 2. Modes of action of killer toxins according to Klassen et al. (2017), modified [40]. Killer toxins can be divided into four categories regarding the cell target. The T1 and T2 toxin types use β-1,6-glucan as the primary cell wall receptor (in figure, toxins targeting the cell wall or the membrane). T1-type toxins such as K1, K2 and PMKT [42][43][44][45][46] bind to a membrane receptor and induce membrane pore formation; T2-type toxins, widely recognized in the genus Wickerhamomyces [47], act as glucanases. T3 and T4 toxin types have intracellular targets. K28 encoded by S. cerevisiae [26][48] is an example of a T3 toxin, which uses mannoproteins as the primary receptor, reaches the nucleus via retrograde transport and inhibits DNA synthesis. T4-type toxins, such as zymocin, PaT and PiT [49][50][51][52] first bind to a chitin receptor in the cell wall. Subsequently, a membrane receptor is required for the cellular uptake of the toxic subunit which cleaves the respective RNA target (tRNAglu, tRNAgln and rRNA).
For more detailed information on killer toxin action and self-protection of killer yeasts, we refer to recent reviews and publications dealing with the specific subject in depth: Schaffrath et al. (2018) [23]; Schmitt and Breining (2002, 2006) [26][48]; Kast et al. (2015) [49]; Satwika et al. (2012) [29].
3. Application of Killer Yeasts as Biocontrol Agents
Fruit and vegetables seem to provide a favorable environment for yeasts in general and for killer yeasts in particular since approximately a quarter of the strains isolated from such source display the phenotype [21][24]. Killer yeasts have a great potential to act as biocontrol agents, producing toxins which do not harm humans or the fruit [53]. Moreover, killer yeasts display fruit-protective qualities in both pre- and postharvest applications.
3.1. Preharvest Application of Killer Yeast
Though there is only a limited number of studies dealing with killer yeasts as preharvest antagonists, such as in citrus and grapes, the reported cases clearly furnish evidence of their biocontrol potential when applied this way.
A rather common preharvest fungal pathogen of grapes is C. gloeosporioides which generates an anthracnose disease characterized by irregular-shaped black necrotic spots, the centers of which—in the course of the disease—become whitish gray surrounded by narrow reddish-brown to black margins [54]. For controlling anthracnose, grapes are routinely medicated with fungicides such as fluazinam and chlorothalonil [55].
Only rather recently, three killer yeasts were isolated displaying efficient antagonistic activity against the anthracnose causative pathogen. The protective potential of S. cerevisiae GA8, S. cerevisiae LFA802 and S. cerevisiae L24 was proven. Each of the three strains was able to produce extracellular antifungal agents such as β-1, 3-glucanases and chitinases; strain GA8 additionally synthesized volatile compounds and thermostable metabolites with the potential to obstruct C. gloeosporioides. In addition, such strains were able to colonize wounds and stably maintained a vital state on the surface of the berries. Moreover, each of them was able to inhibit germination of the phytopathogen’s spores [56].
B. cinerea is a wide spread, ubiquitously occurring fungus; it is the causative agent of the gray mold disease in a wide range of hosts provoking considerable yield losses of grape, lettuce, onion, potato, strawberry, tomato, and other fruits of commercial interest. In 2004, Santos et al. [57] purified and characterized the CYC 1106 killer toxin from Pichia membranifaciens, a strain isolated from olive brines to be used against grey mold. In in vivo assays, Vitis vinifera plants developed the characteristic grey mold symptoms when inoculated with B. cinerea (control treatment). However, all of the plants treated with a mixture of P. membranifaciens and B. cinerea remained healthy, while 20% of the plants treated solely with the purified toxin depicted the pathology. According to the authors, such differences could be attributed to other yeast action mechanisms such as the production of hydrolytic enzymes or the competition for nutrients [56].
Colletotrichum acutatum is one of the most devastating fungi affecting citrus during preharvest stages. It is the causative agent of the post bloom fruit drop. By attacking flower tissues and changing normal fruit abscission persistent calyces and peduncles develop. Lopes et al. (2015) [58], investigated six S. cerevisiae killer yeast strains (ACB-CR1, ACB-KD1, ACB-CAT1, ACB-BG1, ACB-K1 and ACB-PE2) isolated from commercial ethanol fermentations in Brazil with respect to their bio-control potential for C. acutatum. ACB-CAT1 and ACB-CR1 in vitro most efficiently inhibited mycelial growth, i.e., 71% and 67%, respectively. All of the isolates were both, curative and preventive: 73–84% of the flowers were asymptomatic after curative treatments, and 50–86% remained healthy when the treatment was preventive. After analyzing putative action mechanisms, all strains were shown to produce antifungal compounds; they competed for nutrients, inhibited the pathogen´s spore germination, and produced killer toxins and hydrolytic enzymes.
3.2. Postharvest Application of Killer Yeasts
In spite of the rather promising results obtained in preharvest biocontrol attempts, postharvest use of killer yeasts is most common [59].
In citrus, the main fungal postharvest affliction is the “green mold disease” produced by P. digitatum. Control efforts routinely include chemical fungicides such as imazalil or thiabendazole [60]. Seeking for safer alternatives, we isolated more than 400 native yeasts originating from citrus, 8.5% of which showed killer activities [61]. In in vivo assays, two strains of Pichia sp. (strains 27 and 28), and Wickerhamomyces sp. strain 56 conferred significant shelter from the decay caused by P. digitatum. Efficiencies reached 93.6%, 82.5%, and 72.5%, respectively. Furthermore, the native killer yeast Clavispora lusitaniae 146 displayed, again in in vivo tests, strong antagonistic activities against the fungal pathogen (Figure 3), conferring consistent protection during the entire harvesting period [18]. The biocontrol potential of C. lusitaniae 146 was seen to be stronger than that of a commercial product containing C. oleophila. Moreover, the strain was able to grow with agriculturally applied concentrations of fungicides, which allows for the combined use of the yeast along with—then minor—doses of the fungicides. The strain’s commercial potential unequivocally became evident when we studied action mechanisms, such as wound colonization and the inhibition of P. digitatum spore germination, as C. lusitaniae 146 was seen to efficiently control even a fungicide-resistant P. digitatum strain. In addition, strain 146 recently demonstrated a broad range of protective activity in various citrus varieties besides lemons, such as mandarins, oranges, and grapefruits [62][63][64].
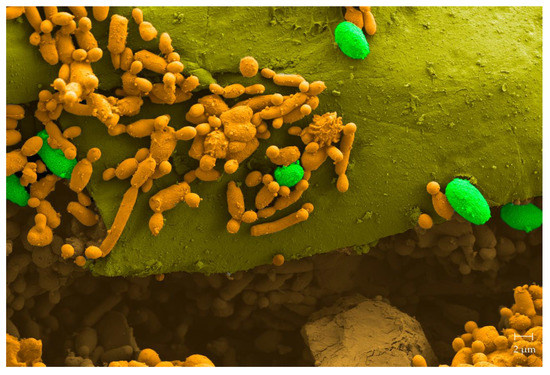
Figure 3. Colored scanning electron microscope (SEM) image of the surface of a lemon wound inoculated with the killer yeast C. lusitaniae 146. The image shows a cross section of a lemon wound where colonization by the killer yeast (orange) and ungerminated spores of citrus phypathogen P. digitatum (green) are observed.
Several killer species are able to control the green mold disease of the Tarocco orange (Citrus sinensis), a predominant cultivar in Eastern Sicily/Italy [65]. S. cerevisiae (strains BS46 and BCA61) and Wickerhamomyces anomalus (strains BCU24, BS91, BS92, and BCA15) isolated from fermented olives were tested in vivo and in vitro for their capability to restrain P. digitatum. Only W. anomalus gave positive results in vitro; the in vivo antagonism assays eventually identified W. anomalus BS91, BS92 and BCA15 to be most effective with respect to the reduction of the disease incidence (1, 4 and 44%, respectively). The deleterious and vice versa the positive effect on the pathogen and the cultivar, respectively, lasted up to 10 storage days counted from the beginning of the artificially provoked infection. In vitro, W. anomalus BS91 had an effect on mycelial structures of P. digitatum: the hyphae became wilted, folded, and coiled with a grainy appearance, an effect presumably due to the production of hydrolytic enzymes by BS91 (β-1, 3-glucanase) as previously reported by Muccilli et al. [47][66].
Only rather recently yeasts capable of antagonizing Penicillium italicum in “Valencia” sweet oranges ((Citrus sinensis (L.) Osbeck (rutaceae)) were isolated [67]. The fungus can infect injured fruit peels, generating the symptomatology known as “blue mold disease”. Sapping all parts of the fruit the fungus makes infected oranges uneatable. From 14 citrus producing areas of the São Paulo State/Brazil, ninety-seven yeast strains were isolated from leaves, flowers, fruits, and soils. Screening for protective representatives was performed in vivo and in vitro. Candida stellimalicola strains ACBL-04 and ACBL-05, and S. cerevisiae ACBL-11 and ACBL-10 were most successful in preventive treatments, whereas ACBL-08 additionally displayed curative effects. With respect to the control mechanism, the killer trait appeared to be crucial for the biocontrol of P. italicum.
Monilinia fructigena and Monilinia fructicola are major pathogens of stone fruit (peaches and plums). They cause a panoply of symptoms including blossom blighting, woody cankers, and preharvest fruit rotting; however, the most serious damage happens after harvesting during transport and storage. Since currently there is no efficient treatment to control Monilinia spp. infections, biocontrol agents, among them killer yeasts, were considered to presumably meet the challenge [68]. Debaryomyces hansenii MI1a and D. hansenii K12a, isolated from blue-veined Rokpol cheese, and W. anomalus BS91, previously shown control P. digitatum in oranges (see above and [65]), were selected due to their killer phenotype and antagonistic activity against other fungal pathogens. Exclusively W. anomalus BS91 was able to inhibit both of the Monilinia pathogens by producing volatile antifungal compounds in an in vitro dual culture method. In vivo antagonism tests identified D. hansenii MI1A as totally M. fructicola ineffective in both plum and peach, and there was only a weak impact on M. fructigena. D. hansenii K12a and W. anomalus BS91 conferred clear fruit protection. When peaches were treated 24 h prior to inoculation with M. fructigena, the disease severity was mitigated to 33 and 25%, respectively. For the plums, a similar effect against M. fructigena was seen. The effect observed for peaches was even more promising as both, D. hansenii K12a and W. anomalus BS91 lessened disease severity caused by M. fructicola to 55%, when applied 24 h before the fungus was added. For plums, the best results were obtained with W. anomalus BS91 that reduced disease severity to 30%.
Cells and killer toxins from D. hansenii strains (TEM8 and TEM7), previously isolated from a Turkish-style homemade dairy product [69], were tested for their biocontrol efficiency in tomato against Alternaria brassicicola and Rhizopus stolonifer [70]. A. brassicicola is a necrotrophic fungus that causes the black spot disease in a broad range of host plants [71] and R. stolonifer is an important soft rot-causing fungus affecting tomatoes during postharvest stages [72]. The efficiency of the toxin was compared with the action of viable yeast cells. When the fruit was inoculated with a killer yeast or treated with killer toxins prior to applying fungal spore suspensions, the lesion sizes were smaller than for the control, and there were even samples with no damages at all. Furthermore, the killer proteins and the cells also cropped up to be effective against Alternaria citri and A. niger in lemons and apples, indicating a wider application spectrum than solely for tomatoes.
In the north of Brazil where papaya crops agriculturally predominate, C. gleosporoides, the causative agent of anthracnosis, commonly produces severe postharvest losses. Additionally, the pathogen effectuates small reddish-brown surface patches unattractively modifying the fruit´s physical appearance. Two killer yeast species isolated from street markets and producer farms in the Ceará State/Brazil, W. anomalus 422 and Meyerozyma guilliermondii 443 [73] were able to reduce the diameter of disease lesions by 31.4 and 41.17%, respectively, when applied 24 h prior to the application of the pathogen. Such killer yeasts displayed protective rather than curative effects; they were able to reduce both the incidence and pathogenicity of C. gleosporoides presumably due to multiple factors such as competition for space and nutrients, mycoparasitism and secretion of β-1, 3-glucanases, the latter probably attacking the phytopathogen´s cell wall [74].
In apples, M. fructigena is the causative agent of the brown rot. The fungus infects wounds and subsequently–and rapidly–develops the rather firm rot. The respective areas are arranged as small cottony masses forming pathogen characteristic concentric circles [75]. Five killer yeasts, isolated from the peel of apples, were tested in vivo and in vitro for their potential to tame the pathogen: Schwanniomyces vanrijiae, Galactomyces geotrichum, Pichia kudriavzevii, D. hansenii and Rhodotorula glutinis [76]. In vivo tests included three different approaches: (1) simultaneous application of both the antagonist and the pathogen, (2) placing the yeast 24 h prior to and (3) 24 h after infection. Disease incidence decreases of 84.02–89.5%, 80.1–86.9% and 56.3–86.9%, respectively, were obtained. In addition, it was shown that hydrolytic enzymes such as chitinases, pectinases, β-1, 3-glucanases, and proteases were produced.
More recently, W. anomalus BS91, D. hansenii K12a and D. hansenii AII4b were used to protect apples against M. fructicola [77]. All of the above strains were additionally and successfully tested against Monilinia spp. in stone fruits as well [68]. W. anomalus BS91 and D. hansenii K12a inhibited mycelial growth in vitro by 69 and 66%, respectively. D. hansenii AII4b was less effective (56%). Nevertheless, in vivo BS91 and K12a reduced the M. fructicola disease by 92 and 85%, respectively, while AII4b was less effective (70% disease reduction). Compared to untreated fruit, rot severity was reduced by approximately 52 up to 69%. Stress induction assays by measuring peroxidase and catalase levels in the peel of fruit indicated highest levels of peroxidase (POD) when wounds were treated with W. anomalus BS91. Lowest catalase (CAT) values were obtained with D. hansenii AII4b. The above findings led to the conclusion that the yeasts were able to modulate the activities of CAT and POD in apple tissues, leading the authors to suggest the yeasts promising candidates for developing effective biofungicides against M. fructicola.
Grapes are almost routinely infected by the rot-causing A. carbonarius and A. niger, both responsible for the accumulation of ochratoxin A, a potent nephrotoxic and carcinogenic compound that can even be found in secondary grape products [78][79].
Epiphytic yeasts isolated from grapes in Southern Italy, after molecular identification and inspection of the killer phenotype, were tested in vivo and in vitro to identify those representatives which are able to conquer the co-inoculated pathogenic fungi in their common ecological niches. Issatchenkia orientalis 17C2 and 16C2 efficiently reduced disease incidences of A. carbonarius and A. niger on grapes (100% growth reduction), due to the production of secondary antifungal metabolites and competition for essential nutrients [80].
References
- Nunes, C.A. Biological control of postharvest diseases of fruit. Eur. J. Plant Pathol. 2012, 133, 181–196.
- Costa, L.G.; Giordano, G.; Guizzetti, M.; Vitalone, A. Neurotoxicity of pesticides: A brief review. Front. Biosci. 2008, 13, 1240–1249.
- Jankowska, M.; Kaczynski, P.; Hrynko, I.; Lozowicka, B. Dissipation of six fungicides in greenhouse-grown tomatoes with processing and health risk. Environ. Sci. Pollut. Res. 2016, 23, 11885–11900.
- Muri, S.D.; van der Voet, H.; Boon, P.E.; van Klaveren, J.D.; Brüschweiler, B.J. Comparison of human health risks resulting from exposure to fungicides and mycotoxins via food. Food Chem. Toxicol. 2009, 47, 2963–2974.
- Wightwick, A.; Walters, R.; Allinson, G.; Reichman, S.; Menzies, N. Environmental risks of fungicides used in horticultural production systems. In Fungicides; Carisse, O., Ed.; InTech: Rijeka, Croatia, 2009; pp. 273–304.
- Brent, K.J.; Hollomon, D.W. Fungicide Resistance in Crop Pathogens: How Can It Be Managed? 2nd ed.; Fungicide Resistance Action Committee: Brussels, Belgium, 2007; p. 48.
- Eckert, J.W.; Sievert, J.R.; Ratnayake, M. Reduction of imazalil effectiveness against citrus green mold in California packinghouses by resistant biotypes of Penicillium digitatum. Plant Dis. 1994, 78, 971–974.
- Lucas, J.A.; Hawkins, N.J.; Fraaije, B.A. The evolution of fungicide resistance. Adv. Appl. Microbiol. 2015, 90, 29–92.
- Chandler, D.; Bailey, A.S.; Tatchell, G.M.; Davidson, G.; Greaves, J.; Grant, W.P. The development, regulation and use of biopesticides for integrated pest management. Philos. Trans. R. Soc. Lond. 2011, 366, 1987–1998.
- Ehler, L.E. Integrated pest management (IPM): Definition, historical development and implementation, and the other IPM. Pest Manag. Sci. 2006, 62, 787–789.
- Giles, K.L.; McCornack, B.P.; Royer, T.A.; Elliott, N.C. Incorporating biological control into IPM decision making. Curr. Opin. Insect Sci. 2017, 20, 84–89.
- Matyjaszczyk, E. Products containing microorganisms as a tool in integrated pest management and the rules of their market placement in the European Union. Pest Manag. Sci. 2015, 71, 1201–1206.
- Owen, M.D.; Beckie, H.J.; Leeson, J.Y.; Norsworthy, J.K.; Steckel, L.E. Integrated pest management and weed management in the United States and Canada. Pest Manag. Sci. 2018, 71, 357–376.
- Walker, J.T.; Suckling, D.M.; Wearing, C.H. Past, present, and future of integrated control of apple pests: The New Zealand experience. Annu. Rev. Entomol. 2017, 62, 231–248.
- Chalutz, E.; Ben-Arie, R.; Droby, S.; Cohen, L.; Weiss, B.; Wilson, C.L. Yeasts as biocontrol agents of postharvest diseases of fruits. Phytoparasit. Isr. J. Plant Prot. Sci. 1988, 16, 69.
- Janisiewicz, W.J. Postharvest biological control of blue mold on apples. Phytopathology 1987, 77, 481–485.
- Janisiewicz, W.J. Biocontrol of postharvest diseases of apples with antagonist mixtures. Phytopathology 1988, 78, 194–198.
- Perez, M.F.; Ibarreche, J.P.; Isas, A.S.; Sepulveda, M.; Ramallo, J.; Dib, J.R. Antagonistic yeasts for the biological control of Penicillium digitatum on lemons stored under export conditions. Biol. Control 2017, 115, 135–140.
- Vero, S.; Garmendia, G.; González, M.B.; Bentancur, O.; Wisniewski, M. Evaluation of yeasts obtained from Antarctic soil samples as biocontrol agents for the management of postharvest diseases of apple (Malus× domestica). FEMS Yeast Res. 2013, 13, 189–199.
- Droby, S.; Wisniewski, M.; Teixidó, N.; Spadaro, D.; Jijakli, M.H. The science, development, and commercialization of postharvest biocontrol products. Postharvest Biol. Technol. 2016, 122, 22–29.
- Magliani, W.; Conti, S.; Gerloni, M.; Bertolotti, D.; Polonelli, L. Yeast killer systems. Clin. Microbiol. Rev. 1997, 10, 369–400.
- Meinhardt, F.; Klassen, R. Yeast killer toxins: Fundamentals and applications. In Physiology and Genetics; Springer: Berlin/Heidelberg, Germany, 2009; pp. 107–130.
- Schaffrath, R.; Meinhardt, F.; Klassen, R. Yeast killer toxins: Fundamentals and applications. In Physiology and Genetics; Springer: Cham, Switzerland, 2018; pp. 87–118.
- Starmer, W.T.; Ganter, P.F.; Aberdeen, V.; Lachance, M.A.; Phaff, H.J. The ecological role of killer yeasts in natural communities of yeasts. Can. J. Microbiol. 1987, 33, 783–796.
- Riley, M.A.; Wertz, J.E. Bacteriocins: Evolution, ecology, and application. Annu. Rev. Microbiol. 2002, 56, 117–137.
- Schmitt, M.J.; Breinig, F. The viral killer system in yeast: From molecular biology to application. FEMS Microbiol. Rev. 2002, 26, 257–276.
- Kaufmann, G. Anticodon nucleases. Trends Biochem. Sci. 2000, 25, 70–74.
- Konisky, J. Colicins and other bacteriocins with established modes of action. Ann. Rev. Microbiol. 1982, 36, 122–145.
- Satwika, D.; Klassen, R.; Meinhardt, F. Anticodon nuclease encoding virus-like elements in yeast. Appl. Microbial. Biot. 2012, 96, 345–356.
- Bevan, E.A.; Makower, M. The physiological basis of the killer character in yeast. Proc. XIth Int. Congr. Genet 1963, 1, 202–203.
- Woods, D.R.; Bevan, E.A. Studies on the nature of the killer factor produced by Saccharomyces cerevisiae. Microbiology 1968, 51, 115–126.
- Ciani, M.; Fatichenti, F. Killer Toxin of Kluyveromyces phaffii DBVPG 6076 as a Biopreservative Agent to Control apiculate wine yeasts. Appl. Environ. Microbiol. 2001, 67, 3058–3063.
- Goretti, M.; Turchetti, B.; Buratta, M.; Branda, E.; Corazzi, L.; Vaughan-Martini, A.; Buzzini, P. In vitro antimycotic activity of a Williopsis saturnus killer protein against food spoilage yeasts. Int. J. Food Microbiol. 2009, 131, 178–182.
- Somers, J.M.; Bevan, E.A. The inheritance of the killer character in yeast. Genet. Res. 1969, 13, 71–83.
- Adler, J.; Wood, H.A.; Bozarth, R.F. Virus-like particles from killer, neutral, and sensitive strains of Saccharomyces cerevisiae. J. Virol. 1976, 17, 472–476.
- Berry, E.A.; Bevan, E.A. A new species of double-stranded RNA from yeast. Nature 1972, 239, 279–280.
- Herring, A.J.; Bevan, E.A. Virus-like particles associated with the double-stranded RNA species found in killer and sensitive strains of the yeast Saccharomyces cerevisiae. J. Gen. Virol. 1974, 22, 387–394.
- Vodkin, M.H.; Fink, G.R. A nucleic acid associated with a killer strain of yeast. Proc. Natl. Acad. Sci. USA 1973, 70, 1069–1072.
- Vodkin, M.H.; Katterman, F.; Fink, G.R. Yeast killer mutants with altered double-stranded ribonucleic acid. J. Bacteriol. 1974, 117, 681–686.
- Klassen, R.; Schaffrath, R.; Buzzini, P.; Philip Ganter, P.F. Antagonistic interactions and killer yeasts. In Yeasts in Natural Ecosystems: Ecology; Buzzini, P., Lachance, M.A., Yurkov, A., Eds.; Springer: Cham, Switzerland, 2017; pp. 229–275.
- Philliskirk, G.; Young, T.W. The occurrence of killer character in yeasts of various genera. Antonie Leeuwenhoek 1975, 41, 147.
- Young, T.W.; Yagiu, M. A comparison of the killer character in different yeasts and its classification. Antonie Leeuwenhoek 1978, 44, 59–77.
- Dignard, D.; Whiteway, M.; Germain, D.; Tessier, D.; Thomas, D.Y. Expression in yeast of a cDNA copy of the K2 killer toxin gene. Mol. Gen. Genet. 1991, 227, 127–136.
- Lukša, J.; Podoliankaitė, M.; Vepštaitė, I.; Strazdaitė-Žielienė, Ž.; Urbonavičius, J.; Servienė, E. Yeast β-1, 6-glucan is a primary target for the Saccharomyces cerevisiae K2 toxin. Eukaryot. Cell 2015, 14, 406–414.
- Orentaite, I.; Poranen, M.M.; Oksanen, H.M.; Daugelavicius, R.; Bamford, D.H. K2 killer toxin-induced physiological changes in the yeast Saccharomyces cerevisiae. FEMS Yeast Res. 2016, 16, 3.
- Santos, A.; Marquina, D.; Leal, J.A.; Peinado, J.M. Biology of killer yeasts. Int. J. Microbiol. 2002, 5, 65–71.
- Muccilli, S.; Wemhoff, S.; Restuccia, C.; Meinhardt, F. Exoglucanase-encoding genes from three Wickerhamomyces anomalus killer strains isolated from olive brine. Yeast 2013, 30, 33–43.
- Schmitt, M.J.; Breinig, F. Yeast viral killer toxins: Lethality and self-protection. Nat. Rev. Microbiol. 2006, 4, 212–221.
- Kast, A.; Voges, R.; Schroth, M.; Schaffrath, R.; Klassen, R.; Meinhardt, F. Autoselection of Cytoplasmic Yeast Virus Like Elements Encoding Toxin/Antitoxin Systems Involves a Nuclear Barrier for Immunity Gene Expression. PLoS Genet. 2015, 11, e1005005.
- Schaffrath, R.; Meacock, P.A.; Meinhardt, F. Yeast killer plasmid pGKL2: Molecular analysis of UCS5, a cytoplasmic promoter element essential for ORF5 gene function. Mol. Gen. Genet. 1996, 250, 286–294.
- Meinhardt, F.; Schaffrath, R. Extranuclear inheritance: Cytoplasmic linear double-stranded DNA killer elements of the dairy yeast Kluyveromyces lactis. In Progress in Botany; Springer: Berlin/Heidelberg, Germany, 2001; Volume 62, pp. 51–70.
- Klassen, R.; Teichert, S.; Meinhardt, F. Novel yeast killer toxins provoke S-phase arrest and DNA damage checkpoint activation. Mol. Microbiol. 2004, 53, 263–273.
- Ocampo-Suarez, I.B.; López, Z.; Calderón-Santoyo, M.; Ragazzo-Sánchez, J.A.; Knauth, P. Are biological control agents, isolated from tropical fruits, harmless to potential consumers? Food Chem. Toxicol. 2017, 109, 1055–1062.
- Ellis, M.A.; Erincik, O. Anthracnose of grape. In Agriculture and Natural Resources; Keith, L.S., Ed.; The Ohio State University: Columbus, OH, USA, 2008; pp. 1–3
- Magarey, R.D.; Emmett, R.W.; Magarey, P.A.; Franz, P.R. Evaluation of control of grape anthracnose caused by Elsinoe ampelina by preinfection fungicides. Australas Plant Pathol. 1993, 22, 48–52.
- Liu, Z.; Du, S.; Ren, Y.; Liu, Y. Biocontrol ability of killer yeasts (Saccharomyces cerevisiae) isolated from wine against Colletotrichum gloeosporioides on grape. J. Basic Microbiol. 2018, 58, 60–67.
- Santos, A.; Marquina, D. Killer toxin of Pichia membranifaciens and its possible use as a biocontrol agent against grey mould disease of grapevine. Microbiology 2004, 150, 2527–2534.
- Lopes, M.R.; Klein, M.N.; Ferraz, L.P.; da Silva, A.C.; Kupper, K.C. Saccharomyces cerevisiae: A novel and efficient biological control agent for Colletotrichum acutatum during pre-harvest. Microbiol. Res. 2015, 175, 93–99.
- Viñas, I. Principios básicos de la patología de poscosecha. Frut 1990, 5, 285–292.
- Sánchez-Torres, P.; Tuset, J.J. Molecular insights into fungicide resistance in sensitive and resistant Penicillium digitatum strains infecting citrus. Postharvest Biol. Technol. 2011, 59, 159–165.
- Perez, M.F.; Contreras, L.; Garnica, N.M.; Fernández-Zenoff, M.V.; Farías, M.E.; Sepulveda, M. Native Killer Yeasts as Biocontrol Agents of Postharvest Fungal Diseases in Lemons. PLoS ONE 2016, 11, e0165590.
- Perez, M.F.; Díaz, M.A.; Pereyra, M.M.; Córdoba, J.M.; Isas, A.S.; Sepúlveda, M.; Dib, J.R. Biocontrol features of Clavispora lusitaniae against Penicillium digitatum on lemons. Postharvest Biol. Technol. 2019, 155, 57–64.
- Pereyra, M.M.; Díaz, M.A.; Meinhardt, F.; Dib, J.R. Effect of stress factors associated with postharvest citrus conditions on the viability and biocontrol activity of Clavispora lusitaniae strain 146. PLoS ONE 2020, 15, e0239432.
- Díaz, M.A.; Pereyra, M.M.; Santander, F.F.S.; Perez, M.F.; Córdoba, J.M.; Alhussein, M.; Dib, J.R. Protection of Citrus Fruits from Postharvest Infection with Penicillium digitatum and Degradation of Patulin by Biocontrol Yeast Clavispora lusitaniae 146. Microorganisms 2020, 8, 1477.
- Platania, C.; Restuccia, C.; Muccilli, S.; Cirvilleri, G. Efficacy of killer yeasts in the biological control of Penicillium digitatum on Tarocco orange fruits (Citrus sinensis). Food Microbiol. 2012, 30, 219–225.
- Muccilli, S.; Wemhoff, S.; Restuccia, C.; Meinhardt, F. Molecular genetics of Pichia anomala killer strains isolated from naturally fermented olive brine. J. Biotechnol. 2010, 150, 302.
- Da Cunha, T.; Ferraz, L.P.; Wehr, P.P.; Kupper, K.C. Antifungal activity and action mechanisms of yeasts isolates from citrus against Penicillium italicum. Int. J. Food Microbiol. 2018, 276, 20–27.
- Grzegorczyk, M.; Żarowska, B.; Restuccia, C.; Cirvilleri, G. Postharvest biocontrol ability of killer yeasts against Monilinia fructigena and Monilinia fructicola on stone fruit. Food Microbiol. 2017, 61, 93–101.
- Corbacı, C.; Ucar, F.B.; Yalcin, H.T. Isolation and characterization of yeasts associated with Turkish-style homemade dairy products and their potential as starter cultures. Afr. J. Microbiol. Res. 2012, 6, 534–542.
- Çorbacı, C.; Uçar, F.B. Purification, characterization and in vivo biocontrol efficiency of killer toxins from Debaryomyces hansenii strains. Int. J. Biol. Macromol. 2018, 119, 1077–1082.
- Macioszek, V.K.; Lawrence, C.B.; Kononowicz, A.K. Infection cycle of Alternaria brassicicola on Brassica oleracea leaves under growth room conditions. Plant Pathol. J. 2018, 67, 1088–1096.
- Bautista-Baños, S.; Velaquez-Del Valle, M.G.; Hernandez-Lauzardoa, A.N.; Barka, E.A. The Rhizopus stolonifer-Tomato interaction. In Plant-Microbe Interactions; Ait Barka, E., Clément, C., Eds.; Research Signpost: Kerala, India, 2008; pp. 269–289.
- Lima, J.R.; Goncalves, L.R.B.; Brandão, L.R.; Rosa, C.A.; Viana, F.M.P. Isolation, identification and activity in vitro of killer yeasts against Colletotrichum gloeosporioides isolated from tropical fruits. J. Basic Microbiol. 2012, 52, 1–10.
- Lima, J.R.; Gondim, D.M.; Oliveira, J.T.A.; Oliveira, F.S.; Gonçalves, L.R.; Viana, F.M. Use of killer yeast in the management of postharvest papaya anthracnose. Postharvest Biol. Technol. 2013, 83, 58–64.
- Di Francesco, A.; Mari, M. Monilinia species of fruit decay: A comparison between biological and epidemiological data. Ital. J. Mycol. 2018, 47, 13–23.
- Madbouly, A.K.; Elyousr, K.A.A.; Ismail, I.M. Biocontrol of Monilinia fructigena, causal agent of brown rot of apple fruit, by using endophytic yeasts. Biol. Control 2020, 144, 104239.
- Czarnecka, M.; Żarowska, B.; Połomska, X.; Restuccia, C.; Cirvilleri, G. Role of biocontrol yeasts Debaryomyces hansenii and Wickerhamomyces anomalus in plants’ defence mechanisms against Monilinia fructicola in apple fruits. Food Microbiol. 2019, 83, 1–8.
- Battilani, P.; Pietri, A.; Bertuzzi, T.; Languasco, L.; Giorni, P.; Kozakiewicz, Z. Occurrence of ochratoxin A-producing fungi in grapes grown in Italy. J. Food Prot. 2003, 66, 633–636.
- Zimmerli, B.; Dick, R. Ochratoxin A in table wine and grape juice: Occurrence and risk assessment. Food Addit. Contam. 1996, 13, 655–668.
- Bleve, G.; Grieco, F.; Cozzi, G.; Logrieco, A.; Visconti, A. Isolation of epiphytic yeasts with potential for biocontrol of Aspergillus carbonarius and A. niger on grape. Int. J. Food Microbiol. 2006, 108, 204–209.

