Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Miguel A. López-Unzu | -- | 1490 | 2022-09-30 12:39:51 | | | |
| 2 | Beatrix Zheng | Meta information modification | 1490 | 2022-10-04 15:33:05 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Sans-Coma, V.; Pozo-Vilumbrales, B.; Fernández, M.C.; López-Unzu, M.�.; Soto-Navarrete, M.T.; Durán, A.C.; Arqué, J.M.; Fernández, B. Outflow Tract Valves of the Vertebrate Heart. Encyclopedia. Available online: https://encyclopedia.pub/entry/28171 (accessed on 24 July 2026).
Sans-Coma V, Pozo-Vilumbrales B, Fernández MC, López-Unzu M�, Soto-Navarrete MT, Durán AC, et al. Outflow Tract Valves of the Vertebrate Heart. Encyclopedia. Available at: https://encyclopedia.pub/entry/28171. Accessed July 24, 2026.
Sans-Coma, Valentín, Bárbara Pozo-Vilumbrales, María Carmen Fernández, Miguel Á. López-Unzu, María Teresa Soto-Navarrete, Ana Carmen Durán, Josep M. Arqué, Borja Fernández. "Outflow Tract Valves of the Vertebrate Heart" Encyclopedia, https://encyclopedia.pub/entry/28171 (accessed July 24, 2026).
Sans-Coma, V., Pozo-Vilumbrales, B., Fernández, M.C., López-Unzu, M.�., Soto-Navarrete, M.T., Durán, A.C., Arqué, J.M., & Fernández, B. (2022, September 30). Outflow Tract Valves of the Vertebrate Heart. In Encyclopedia. https://encyclopedia.pub/entry/28171
Sans-Coma, Valentín, et al. "Outflow Tract Valves of the Vertebrate Heart." Encyclopedia. Web. 30 September, 2022.
Copy Citation
The anatomical elements that in humans guard the unidirectional blood flow from the cardiac ventricles to the aortic and pulmonary arteries are the arterial (semilunar) valves. The valve that prevents blood backflow from the aorta to the left ventricle is the aortic valve, while that which performs this function between the pulmonary artery and the right ventricle is the pulmonary or pulmonic valve. The main medical interest in arterial valves is that their congenital malformations and diseases over a lifetime are clinically relevant. Although both valves are subject to similar complications, those affecting the aortic valve cause the most severe effects.
heart
outflow tract valves
vertebrates
medical concept
1. The Cardiac Outflow Tract Valves of Chondrichthyans and Actinopterygians
In chondrichthyans (reviewed in [1]) and early actinopterygians [2], the number and arrangement of the conal valves are highly variable; both traits may even diverge between members of the same species [3]. The valves are usually distributed in several transverse rows along the conus arteriosus, which connects distally with the bulbus arteriosus (Figure 1C). The pocket-like valves are by far the most common type (Figure 1C). They play the main role of preventing blood backflow from the aorta, similar to the function of the mammalian aortic valve. An additional function of the cardiac conal valves in fish is to collaborate in the reduction of aortic pressure to protect the delicate vasculature of the gills [4]. The other, smaller valves (Figure 1C’), when present, are mere protuberances that have much less effect in avoiding flow from the conus arteriosus into the ventricle. Descriptive studies on fossilized hearts of the extinct teleost species Rhacolepis buccalis have shed light on the debate about the direction of evolution of the cardiac outflow tract in osteichthyans. The finding of several rows of valves in the conus arteriosus of the species R. buccalis, representative of a basal group of teleosts, suggests that over the course of evolution the number of valves in the conus arteriosus of teleosts has been reduced, eventually dwindling to one or two at present (Maldanis et al., 2016). Indeed, in the extant teleosts, the number of conal valves is smaller in consonance with the reduction in length of the conus arteriosus. A few species belonging to ancient groups possess two transverse rows of valves. Most teleosts, however, have a single row composed of two major pocket-like valves, which often resemble anatomically the outflow tract valves of birds and mammals (Figure 1D). One or two minor pocket-like valves are also present in several species.
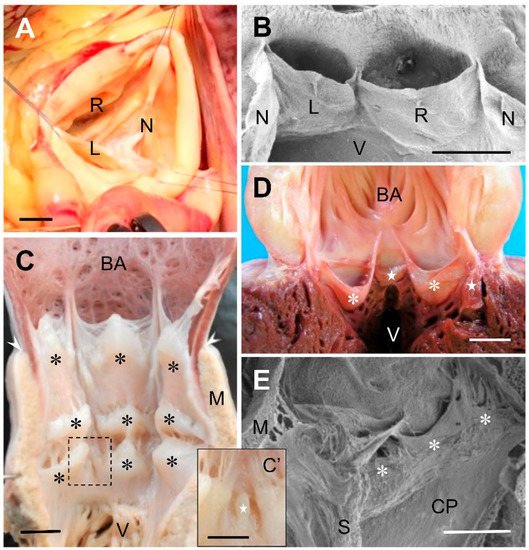
Figure 1. (A) Surgical view of a human normal, tricuspid, aortic valve. (B) Scanning electron micrograph of the aortic valve of a Syrian hamster (Mesocricetus auratus). The valve has been opened through the non-coronary sinus to expose the anterior aspect of the valve. (C) Ventral open-cut view of the cardiac outflow tract of the thresher shark, Alopias vulpinus. The conus arteriosus is furnished with three transverse rows of valves (asteriks), distal, intermediate, and proximal. The rows have three valves each, of similar size. The valves of the distal row are bigger. In the intermediate and proximal rows there are additional valves very reduced in size, ((C’), inset). The arrowheads indicate the anterior limit of the conus myocardium. (D) Ventral open-cut view of the cardiac outflow tract of the Atlantic bluefin tuna, Thunnus thynnus. A single row of four valves, two major (asterisks) and two minor (stars), similar in shape to human valves, are located in the conus, at the base of the bulbus arteriosus (BA). (E) Scanning electron micrograph of the distal region of the cardiac outflow tract of the African clawed frog, Xenopus laevis. The open-cut view shows a set of pocket-like valves (asterisks) located at the distal end of the cavum pulmo-cutaneum (CP). L, left coronary leaflet; M, myocardium; N, non-coronary (dorsal, posterior) leaflet; R, right coronary leaflet; S, spiral valve; V, ventricle. Scale bars: (A): 1 cm; (B–E): 0.5 mm; (C’): 0.25 mm.
2. The Cardiac Outflow Tract Valves of Early Sarcopterygians
In describing the cardiac outflow tract valves of the phylogenetically early groups of sarcopterygians (lobe-finned fishes), such as the crossopterygians (coelacanths), dipnoans (lungfishes), and amphibians, the concept has been usually applied that a valve is a leaflet or even any other anatomical element that might contribute to prevent the return of blood to the heart. As for the crossopterygians, exemplified by the coelacanth Latimeria chalumnae Smith 1939, it was briefly quoted that this species has 24 conal valves, without specifying their morphology [5].
In dipnoans (lungfishes), the concept of the cardiac outflow tract valve has been used ambiguously. The valves are located in the outflow tract portion that has myocardium in its walls. The valve shape is highly variable. There are pocket-like valves of different size, and also transverse protrusions, separated by furrows, and small incisura at the luminal side of the outflow tract, all of which have been described as valves [6][7][8][9].
The shape, size, and spatial distribution of the outflow tract valves are also notably variable in amphibians. In these animals, the valves are also placed in the myocardial portion of the outflow tract, the anatomy of which markedly differs between lung breathing and lungless species. In those having lungs, the outflow tract is partially divided into two channels, the cavum pulmo-cutaneum and the cavum aorticum, by a spiral fold that has been often termed ‘spiral valve’. The spiral fold is vestigial or absent in most lungless amphibians [10]. The other anatomical elements which have been described as valves differ in shape and size, and are usually located at the proximal and distal ends of the myocardial portion of the outflow tract [10][11][12][13]. Most often, they are pocket-like valves (Figure 1E) but other, simpler structures have also been included in the category of valves. In no case, however, has the portion of the outflow tract wall which supports the leaflet been regarded as a valve component.
3. The Cardiac Outflow Tract Valves of Reptiles
The sauropsids included in the classic group of reptiles have a cardiac ventricle divided partially by one or two septa, giving rise to two or three ventricular cavities, respectively. The only ventricle with a complete septum is that of crocodilians. In all cases, blood flows from the heart to the lungs through a single pulmonary trunk that divides into right and left pulmonary arteries, and to the body through two aortic or systemic arteries, right and left. The outflow tract valves are located at the base of the pulmonary trunk and at the anatomical origin of each aorta. These cardiac valves have received little attention and what can be gained from the literature is that they are usually semilunar or pocket-like in shape [14][15][16][17]. At the base of each artery there are generally two valves. Those of the pulmonary trunk have barely been studied. The valves of the aortic trunks, especially those of snakes, have been described in more detail [17]. Interestingly, these authors used the term aortic valve to refer to the set of two valves existing at the base of each aorta, thus opting for the nomenclature applied to birds and mammals.
4. The Cardiac Outflow Tract Valves of Birds and Mammals
In birds, namely the remaining group of sauropsids, the basal portions of the aortic and pulmonary arteries are usually composed of three valves. These two sets of valves have been termed aortic and pulmonary valves, respectively, probably because of the influence of medical viewpoints [18][19][20].
From the zoological viewpoint, mammals (Figure 1A,B), and also birds, regularly have three valves at the base of the aortic and pulmonary trunks. Each of these six valves is, in fact, an anatomical unit composed of a leaflet and its sinus. However, the resulting trivalvular aortic and pulmonary complexes, established evolutionarily in concomitance with the regression of the conus arteriosus, have been successful in performing the enormous work to which they are subject over a lifetime. In this regard, it should be noted that in humans, the presence of three leaflets of similar size constitutes the most efficient geometrical condition to prevent blood backflow to the ventricles [21][22][23]. Valve complexes built up by leaflets of dissimilar sizes or by a different number of leaflets are less efficient; they are considered cardiac anomalies or malformations by physicians, because they very often entail the risk of clinically relevant complications. For example, the presence of only two leaflets in the aortic root, which is normal in the aortic trunks of reptiles, is regarded as an anomaly from the medical perspective [24][25]. In fact, this condition, termed bicuspid or bifoliate aortic valve, is the most frequent congenital cardiac defect in humans, with an estimated prevalence of between 0.5% and 2% [26][27][28]. Considering the serious complications occurring in at least one third of cases, the bicuspid aortic valve may be responsible for more deaths and morbidity than the combined effect of all other congenital heart defects [27]. Interestingly, this example illustrates that a valvular condition that is normal and does not lead to complications in certain vertebrates, such as reptiles, may be the cause of disease in their descendants, namely in mammals.
References
- Sans-Coma, V.; Gallego, A.; Muñoz-Chápuli, R.; De Andrés, A.V.; Durán, A.C.; Fernández, B. Anatomy and histology of the cardiac conal valves of the dogfish (Scyliorhinus canicula). Anat. Rec. 1995, 241, 496–504.
- Grimes, A.C. The cardiac outflow tract of primitive fishes. In Phylogeny, Anatomy and Physiology of Ancient Fishes; Zaccone, G., Dabrowski, K., Hedrick, M.S., Fernandes, J.M.O., Icardo, J.M., Eds.; CRC Press: Boca Raton, FL, USA, 2016; pp. 151–178.
- White, E. The heart valves of the Elasmobranch fishes. Am. Mus. Novitatis 1936, 838, 1–21.
- Satchell, G.H.; Jones, M.P. The function of the conus arteriosus in the Port Jackson shark, Heterodontus portusjacksoni. J. Exp. Biol. 1967, 46, 373–382.
- Anthony, J.; Millot, J.; Robineau, D. Le coeur et l’aorte ventrale de Latimeria chalumnae (Poisson coelacanthidé). Comptes Rendus De L’academie Des Sci., Paris 1965, 261, 223–226.
- Robertson, J.I. The development of the heart and vascular system of Lepidosiren paradoxa. Q. J. Microsc. Sci. 1913, 59, 53–132.
- Burggren, W.W.; Johansen, K. Circulation and respiration in lungfishes (Dipnoi). J. Morphol. 1986, 190, 217–236.
- Szidon, J.P.; Lahiri, S.; Lev, M.; Fishman, A.P. Heart and circulation of the African lungfish. Circ. Res. 1969, 25, 23–38.
- Icardo, J.M.; Brunelli, E.; Perrotta, I.; Colvee, E.; Wong, W.P.; Ip, Y.K. Ventricle and outflow tract of the African lungfish Protopterus dolloi. J. Morphol. 2005, 265, 43–51.
- Putnam, J.L.; Parkerson, J.B. Anatomy of the heart of the Amphibia II. Cryptobranchus alleganiensis. Herpetologica 1985, 41, 287–298.
- Davies, F.; Francis, E.T.B. The heart of the salamander (Salamandra salamandra, L.), with special reference to the conducting (connecting) system and its bearing on the phylogeny of the conducting systems of avian and mammalian hearts. Philos. Trans. R. Soc. Lond. 1941, 231, 99–130.
- Morris, R.W. Function of the anuran conus arteriosus. J. Exp. Biol. 1974, 61, 503–520.
- Lawson, R. The comparative anatomy of the circulatory system. In Hyman’s Comparative Anatomy; Wake, M.H., Ed.; University of Chicago Press: Chicago, IL, USA, 1979; pp. 448–554.
- Kashyap, H.V. The structure of the heart of Typhlops (Reptilia: Ophidia). J. Zool. Soc. India 1950, 2, 42–49.
- Webb, G.J.W.; Heatwole, H.; De Bavay, J. Comparative cardiac anatomy of the Reptilia: II. A critique of the literature on the Squamata and Rhynchocephalia. J. Morphol. 1974, 142, 1–20.
- Webb, G.J.W. Comparative cardiac anatomy of the Reptilia: III. The heart of crocodilians and hypothesis on the completion of the interventricular septum of crocodilians and birds. J. Morphol. 1979, 161, 221–240.
- Young, B.A.; Lillywhite, H.B.; Wassersug, R.J. On the structure of the aortic valves in snakes (Reptilia: Serpentes). J. Morphol. 1993, 216, 141–159.
- López, D.; Durán, A.C.; Sans-Coma, V. Formation of cartilage in semilunar valves of chick and quail. Ann. Anat. 2000, 182, 349–359.
- Bartyzel, B.J.; Charuta, A.; Barszcz, K.; Kolesnik, A.; Kobryn, H. Morphology of the aortic valve of Gallus gallus f. domestica. Bull. Vet. Inst. Pulawy 2009, 53, 147–151.
- Tadjalli, M.; Ghazi, S.R.; Parto, P. Gross anatomy of the heart in ostrich (Struthio camelus). Iran. J. Vet. Res. Schiraz Univ. 2009, 10, 21–27.
- Thubrikar, M.; Piepgrass, W.C.; Shaner, T.W.; Nolan, S.P. The design of the normal aortic valve. Am. J. Physiol. 1981, 241, H795–H801.
- Bierbach, B.O.; Aicher, D.; Issa, O.A.; Bomberg, H.; Glombitza, P.; Schäfers, H.J. Aortic root configuration determine aortic valve function. Eur. J. Cardiothorac. Surg. 2010, 38, 400–406.
- Rankin, J.S.; Bone, M.C.; Fries, P.M.; Aicher, D.; Schäfers, H.J.; Crooke, P.S. A refined hemispheric model of normal aortic valve root geometry. J. Thorac. Cardiovasc. Surg. 2013, 146, 103–108.
- Fedak, P.W. Bicuspid aortic valve syndrome: Heterogeneous but predictable? Eur. Heart J. 2008, 29, 432–433.
- Siu, S.C.; Silversides, C.K. Bicuspid aortic valve disease. J. Am. Coll. Cardiol. 2010, 55, 2789–2800.
- Roberts, W.C. The congenitally bicuspid aortic valve. A study of 85 autopsy cases. Am. J. Cardiol. 1970, 26, 72–83.
- Ward, C. Clinical significance of the bicuspid aortic valve. Heart 2000, 83, 81–85.
- Basso, C.; Boschello, M.; Perrone, C.; Mecenero, A.; Cera, A.; Bicego, D.; Thiene, G.; de Dominicis, E. An echocardiographic survey of primary school children for bicuspid aortic valve. Am. J. Cardiol. 2004, 93, 661–663.
More
Information
Subjects:
Cardiac & Cardiovascular Systems
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.2K
Revisions:
2 times
(View History)
Update Date:
04 Oct 2022
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No