 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | José Pérez de la Lastra | -- | 1226 | 2022-09-27 05:21:31 | | | |
| 2 | Bruce Ren | -21 word(s) | 1205 | 2022-09-29 08:09:24 | | | | |
| 3 | Peter Tang | Meta information modification | 1205 | 2022-09-30 05:34:38 | | |
Video Upload Options
Living species are continuously subjected to all extrinsic forms of reactive oxidants and others that are produced endogenously. There is extensive literature on the generation and effects of reactive oxygen species (ROS) in biological processes, both in terms of alteration and their role in cellular signaling and regulatory pathways. Cells produce ROS as a controlled physiological process, but increasing ROS becomes pathological and leads to oxidative stress and disease. The induction of oxidative stress is an imbalance between the production of radical species and the antioxidant defense systems, which can cause damage to cellular biomolecules, including lipids, proteins and DNA. Cellular and biochemical experiments have been complemented in various ways to explain the biological chemistry of ROS oxidants. However, it is often unclear how this translates into chemical reactions involving redox changes.
1. Introduction
2. Genesis and Effects of ROS
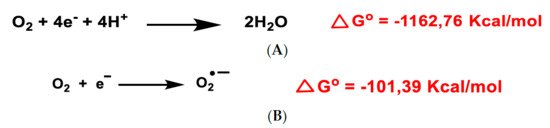

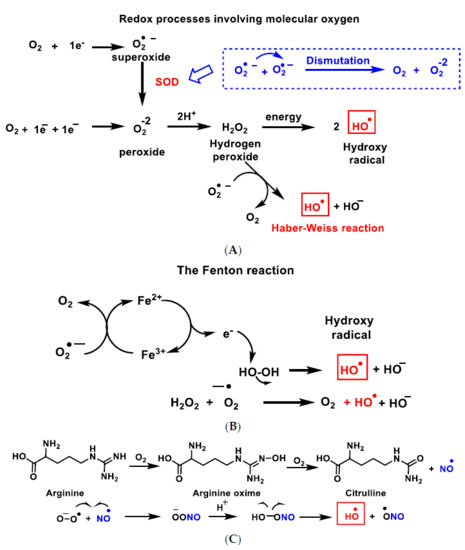
3. Dual Role of ROS: Physiological and Pathological
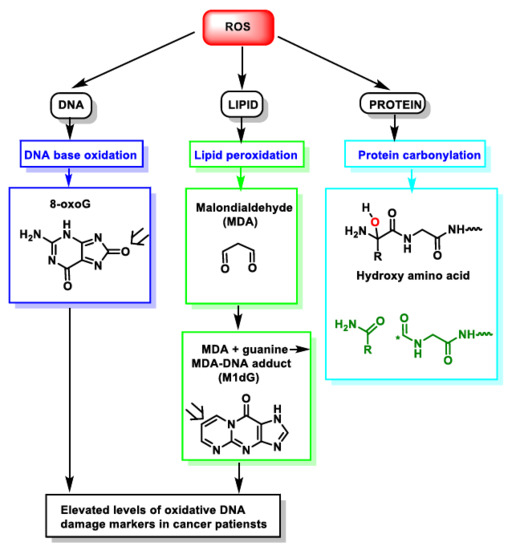
References
- Mailloux, R.J. An update on mitochondrial reactive oxygen species production. Antioxidants 2020, 9, 472.
- Magnani, F.; Mattevi, A. Structure and mechanisms of ROS generation by NADPH oxidases. Curr. Opin. Struct. Biol. 2019, 59, 91–97.
- Collin, F. Chemical basis of reactive oxygen species reactivity and involvement in neurodegenerative diseases. Int. J. Mol. Sci. 2019, 20, 2407.
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383.
- Heid, M.E.; Keyel, P.A.; Kamga, C.; Shiva, S.; Watkins, S.C.; Salter, R.D. Mitochondrial reactive oxygen species induces NLRP3-dependent lysosomal damage and inflammasome activation. J. Immunol. 2013, 191, 5230–5238.
- Sousa, J.S.; D’Imprima, E.; Vonck, J. Mitochondrial Respiratory Chain Complexes. In Membrane Protein Complexes: Structure and Function; Harris, J.R., Boekema, E.J., Eds.; Springer: Singapore, 2018; pp. 167–227.
- Valenta, H.; Erard, M.; Dupré-Crochet, S.; Nüβe, O. The NADPH Oxidase and the Phagosome. In Molecular and Cellular Biology of Phagocytosis; Hallett, M.B., Ed.; Springer International Publishing: Cham, Switzerland, 2020; pp. 153–177.
- Kehrer, J.P. The haber–weiss reaction and mechanisms of toxicity. Toxicology 2000, 149, 43–50.
- Chen, H.-Y. Why the reactive oxygen species of the fenton reaction switches from oxoiron(IV) species to hydroxyl radical in phosphate buffer solutions? A computational rationale. ACS Omega 2019, 4, 14105–14113.
- Fujiwara, O.; Fukuda, S.; Lopez, E.; Zeng, Y.; Niimi, Y.; DeWitt, D.S.; Herndon, D.N.; Prough, D.S.; Enkhbaatar, P. Peroxynitrite decomposition catalyst reduces vasopressin requirement in ovine MRSA sepsis. Intensive Care Med. Exp. 2019, 7, 12.
- Jiang, H.; Xue, A.; Wang, Z.; Xia, R.; Wang, L.; Tang, Y.; Wan, P.; Chen, Y. Electrochemical degradation of lignin by ROS. Sustain. Chem. 2020, 1, 345–360.
- Fisher, A.B.; Zhang, Q. Nadph and Nadph Oxidase. In Encyclopedia of Respiratory Medicine; Laurent, G.J., Shapiro, S.D., Eds.; Academic Press: Oxford, UK, 2006; pp. 77–84.
- Wen, J.J.; Garg, N.J. Manganese superoxide dismutase deficiency exacerbates the mitochondrial ROS production and oxidative damage in Chagas disease. PLoS Negl. Trop. Dis. 2018, 12, e0006687.
- Davalli, P.; Marverti, G.; Lauriola, A.; D’Arca, D. Targeting oxidatively induced DNA damage response in cancer: Opportunities for novel cancer therapies. Oxidative Med. Cell. Longev. 2018, 2018, 2389523.
- Yan, L.L.; Zaher, H.S. How do cells cope with RNA damage and its consequences? J. Biol. Chem. 2019, 294, 15158–15171.
- Ito, F.; Sono, Y.; Ito, T. Measurement and clinical significance of lipid peroxidation as a biomarker of oxidative stress: Oxidative stress in diabetes, atherosclerosis, and chronic inflammation. Antioxidants 2019, 8, 72.
- Hawkins, C.L.; Davies, M.J. Detection, identification, and quantification of oxidative protein modifications. J. Biol. Chem. 2019, 294, 19683–19708.

