Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Jong Heon Kim | -- | 2589 | 2022-09-21 08:43:37 | | | |
| 2 | Catherine Yang | Meta information modification | 2589 | 2022-09-21 08:49:37 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Rhim, J.; Baek, W.; Seo, Y.; Kim, J.H. MicroRNA-21 in Cancer. Encyclopedia. Available online: https://encyclopedia.pub/entry/27414 (accessed on 24 July 2026).
Rhim J, Baek W, Seo Y, Kim JH. MicroRNA-21 in Cancer. Encyclopedia. Available at: https://encyclopedia.pub/entry/27414. Accessed July 24, 2026.
Rhim, Jiho, Woosun Baek, Yoona Seo, Jong Heon Kim. "MicroRNA-21 in Cancer" Encyclopedia, https://encyclopedia.pub/entry/27414 (accessed July 24, 2026).
Rhim, J., Baek, W., Seo, Y., & Kim, J.H. (2022, September 21). MicroRNA-21 in Cancer. In Encyclopedia. https://encyclopedia.pub/entry/27414
Rhim, Jiho, et al. "MicroRNA-21 in Cancer." Encyclopedia. Web. 21 September, 2022.
Copy Citation
MicroRNAs (miRNAs) are small noncoding RNAs that play an important role in regulating gene expression at a posttranscriptional level. As one of the first discovered oncogenic miRNAs, microRNA-21 (miR-21) has been highlighted for its critical role in cancers, such as glioblastoma, pancreatic adenocarcinoma, non-small cell lung cancer, and many others. MiR-21 targets many vital components in a wide range of cancers and acts on various cellular processes ranging from cancer stemness to cell death. Expression of miR-21 is elevated within cancer tissues and circulating miR-21 is readily detectable in biofluids, making it valuable as a cancer biomarker with significant potential for use in diagnosis and prognosis.
noncoding RNA
microRNA-21
cancer
biomarker
RNA therapeutics
1. MicroRNA-21 and Its Role in Cancer
Among the many miRNAs that have been associated with cancer progression, microRNA-21 (miR-21) was among the first to be identified as an oncogenic miRNA or “oncomiR”. Located on chromosome 17q23.2 in the intron of the transmembrane protein 49 (TMEM49)/vacuole membrane protein 1 (VMP1) gene, miR-21 was observed to have a unique, highly conserved promoter region that is activated by activation protein 1 (AP-1) in conjugation with the switch/sucrose non-fermentable (SWI/SNF) complex and Ets-related protein PU.1, CCAAT/enhancer binding protein (C/EBP), nuclear factor I (NFI), serum response factor (SRF), p53, and signal transducer and activator of transcription 3 (STAT3) [1][2]. Alteration of miR-21 expression has been linked to epigenetic factors and the dysregulation of transcriptional and posttranscriptional regulators, whether during biogenesis or through repression, resulting in oncogenic phenotypes (Figure 1).
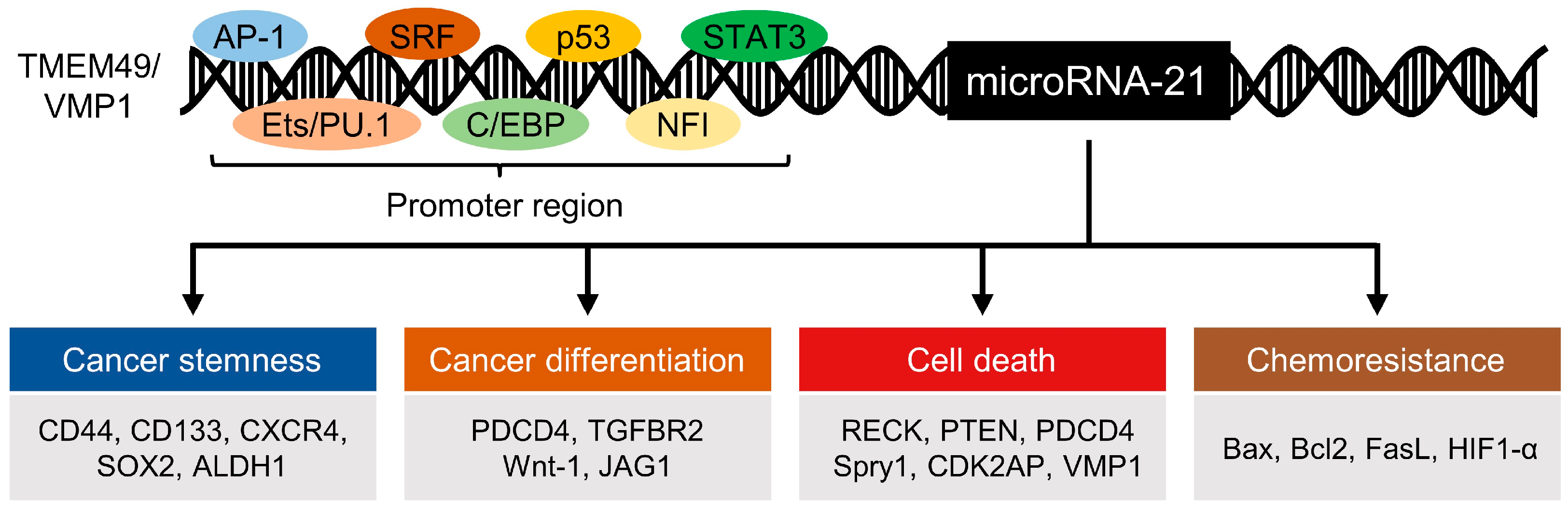
Figure 1. Epigenetic regulators of miR-21 and downstream targets of miR-21 in regards to cancer stemness, differentiation, cell death, and chemoresistance. Representative regulators of each cellular process are listed. TMEM49—transmembrane protein 49, VMP1—vacuole membrane protein 1, AP-1—activator protein 1, Ets/PU.1—Ets family transcription factor PU.1, SRF—serum response factor, C/EBP—CCAAT/enhancer binding protein, NFI—nuclear factor I, STAT3—signal transducer and activator of transcription 3, CXCR4—C-X-C chemokine receptor type 4, SOX2—sex-determining region Y-box, ALDH1—aldehyde dehydrogenase 1, PDCD4—programmed cell death 4, TGFBR2—transforming growth factor beta receptor 2, Wnt-1—wingless-integrated family member 1, JAG1—jagged canonical Notch ligand 1, RECK—reversion-inducing cysteine-rich protein with Kazal motifs, PTEN—phosphatase and tensin homolog, Spry1—sprouty RTK signaling antagonist 1, CDK2AP—cyclin-dependent kinase 2 associated protein, Bax—Bcl-associated X protein, Bcl2—B-cell lymphoma 2, FasL—Fas ligand, and HIF1-α—hypoxia-inducible factor 1-α.
Pivotal driver genes and their associated mutations, which commonly lie at the hub of many cellular pathways, can be found across different cancer types and may lead to dysregulation of or be affected by miR-21. Bailey and colleagues characterized common mutations across at least 15 or more cancer types which include TP53, PIK3CA, KRAS, phosphatase and tensin homolog (PTEN), and ARID1A [3]. Some of these mutations have been closely associated with miR-21 and its target proteins. In non-small cell lung cancer (NSCLC) containing R175H- and R248Q-mutant p53, miR-21 was observed to be upregulated, and the patients with elevated expression of p53 mutations and miR-21 had a lower overall survival rate. In addition, the knockdown of miR-21 led to lower mutant p53 mRNA levels [4]. Furthermore, induction of miR-21 appeared to mediate disease progression and metastasis in p53-deficient tumor keratinocytes [5]. This was also evident in KRAS mutation-driven cancers such as NSCLC and pancreatic ductal adenocarcinoma (PDAC). Inhibition of miR-21 in transgenic KRAS (G12D)/Trp53 null/Pdx1-cre (KPC) cell lines appeared to reduce proliferation, migration, and invasion compared to wild-type pancreatic epithelial cells [6]. Hatley and colleagues found that miR-21 overexpression in G12D-mutant KRAS mouse NSCLC models enhanced tumor formation, and subsequent deletion of miR-21 led to a significant decrease in total tumor mass in proportion to normal lung area [7]. Likewise, epidermal growth factor receptor (EGFR)-mutated lung cancer patients had considerably increased miR-21 expression compared to those without mutations [8]. In particular, EGFR is known to affect miRNA maturation through the posttranslational modification of Ago2, thus highlighting the important relationship between a cancer mutation and miR-21 status [9].
Various independent studies determined that miR-21 is overexpressed in pancreatic adenocarcinoma, breast, and colorectal cancer (CRC) [10][11][12]. In a large-scale study using 540 human samples, miR-21 was found to be commonly overexpressed in lung, breast, stomach, prostate, colon, and pancreatic cancers [13]. MiR-21 was also found to be upregulated in glioblastoma [14][15]. Analysis of TCGA pan-cancer patient databases also reveals a similar trend across various cancers [16] (Figure 2).
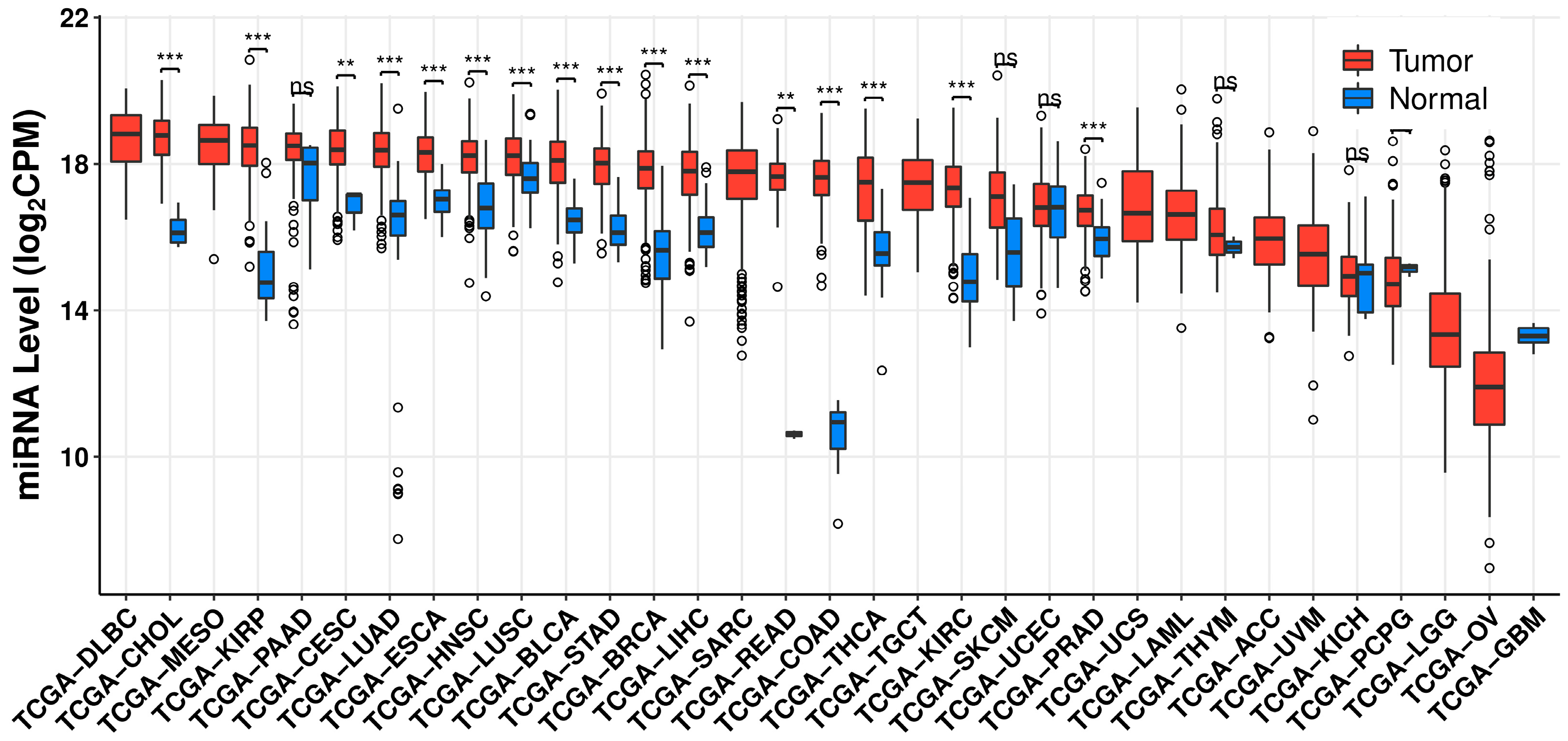
Figure 2. CancerMIRNome analysis of hsa-miR-21-5p expression in TCGA database demonstrating aberrant miR-21 expression across various cancer. Open circles indicate outliers. Some cancers (DLBC, MESO, SARC, TGCT, UCS, LAML, ACC, UVM, LGG, OV, and GBM) are incomparable in the dataset due to the lack of comparable tumor or normal samples. Significance was evaluated by CancerMIRNome via Wilcoxon rank-sum test. ** p < 0.01, *** p < 0.001, ns; not significant. DLBC—diffuse large B-cell lymphoma, CHOL—cholangiocarcinoma, MESO—mesothelioma, KIRP—kidney renal papillary cell carcinoma, PAAD—pancreatic adenocarcinoma, CESC—cervical squamous cell carcinoma and endocervical adenocarcinoma, LUAD—lung adenocarcinoma, ESCA—esophageal carcinoma, HNSC—head and neck squamous cell carcinoma, LUSC—lung squamous cell carcinoma, BLCA—bladder urothelial carcinoma, STAD—stomach adenocarcinoma, BRCA—breast invasive carcinoma, LIHC—liver hepatocellular carcinoma, SARC—sarcoma, READ—rectum adenocarcinoma, COAD—colon adenocarcinoma, THCA—thyroid carcinoma, TGCT—testicular germ cell tumor, KIRC—kidney renal clear cell carcinoma, SKCM—skin cutaneous melanoma, UCEC—uterine corpus endometrial carcinoma, PRAD—prostate adenocarcinoma, UCS—uterine carcinosarcoma, LAML—acute myeloid leukemia, THYM—thymoma, ACC—adrenocortical carcinoma, UVM—uveal melanoma, KICH—kidney chromophobe, PCPG—pheochromocytoma and paraganglioma, LGG—brain lower grade glioma, OV—ovarian serous cystadenocarcinoma, and GBM—glioblastoma multiforme.
Although the mechanisms linking miR-21 upregulation with other oncogenic factors have not yet been elucidated, various reports have indicated that miR-21 could drive cancer progression. In glioma, the β-catenin pathway was found to regulate miR-21 via STAT3, which is a well-known oncogenic transcription factor that promotes tumor growth and invasion [17][18]. MiR-21 also appears to be regulated by EGFR and the activation of β-catenin and AP-1. Upon upregulation, miR-21 regulates EGFR/Akt signaling by targeting the von Hippel–Lindau (VHL) and peroxisome proliferator-activated receptor alpha (PPAR-α) axes [19]. Furthermore, miR-21 upregulation appears to disrupt Ras/mitogen-activated protein kinase (MAPK) signaling via downregulation of Sprouty 2 (Spry2) leading to increased glioma malignancy [20]. MiR-21 also targets Spry2 in PDAC, resulting in the enhancement of epidermal growth factor-induced cell proliferation [21].
In NSCLC, miR-21 was found to repress the tumor suppressor PTEN, and promote growth and invasion phenotypes [22]. In breast cancer, programmed cell death 4 (PDCD4) was found to be directly regulated and repressed by miR-21 [23]. Additionally, miR-21 appeared to target and suppress the expression of tumor suppressor tropomyosin 1 (TPM1), mammary serine protease inhibitor Maspin, and PDCD4 in metastatic breast cancer, thereby affecting tumorigenicity, invasion, and metastasis [24].
Interestingly, Medina and colleagues reported that some cancers depend on one to several oncogenic genes including miR-21 called an “oncomiR addiction”. By using Tet-off and Cre recombinase technology to modulate miR-21 expression in a cross-bred mouse model, the authors showed that miR-21 upregulation led to and was necessary for the malignant phenotype of pre-B-cell lymphoma, which demonstrated this hypothesis. Importantly, the tumor regressed upon inactivation of miR-21, demonstrating the therapeutic validity of targeting miR-21 [25].
2. MicroRNA-21 as a Biomarker in Cancer
MiRNAs are stably detectable in the plasma and serum, and thus may be developed as molecular biomarkers for minimally invasive tool in cancer diagnosis and prognosis [26][27]. MiRNAs can exist in biofluids as either a part of an extracellular vesicle (EV) or a non-vesicle-associated ribonucleoprotein complex and can be used to distinguish patho-logies at various stages of disease progression [28]. Given that miR-21 is highly expressed in a wide variety of cancers and linked to several oncogenic characteristics, it has been evaluated as a diagnostic and prognostic cancer biomarker [29][30][31][32][33][34].
Toiyama and colleagues found that serum miR-21 levels were significantly higher in patients with adenomas and CRC. Furthermore, high miR-21 expression in the serum and tissues could be associated with tumor size, metastasis, and patient survival [35]. Exosomal miRNA profiling of ovarian cancer patients showed that miR-21, miR-141, miR-200a, miR-200b, miR-200c, miR-203, miR-205, and miR-214 were significantly elevated and could be used to distinguish patients from healthy individuals [36]. MiR-21 may even be useful in rare cancers with few available biomarkers. In leptomeningeal metastasis (LM) patients, miR-21 expression was monitored using EVs extracted from cerebrospinal fluid (CSF) and found that aberrant miR-21 expression correlated with patient survival [37][38][39].
The potential use of miRNA as diagnostic or prognostic factors has become increasingly important as accessibility to big data continues to improve. Databases and tools such as Circulating MicroRNA Expression Profiling (CMEP) and CancerMIRNome can help researchers identify miRNAs of interest in a variety of cancers and give a preliminary picture of their clinical significance [16][40].
The use of high-throughput visualization tools, such as BioGRID, could help researchers identify interactions between miR-21 and cellular protein partners, providing further insight into the role of miR-21 (Figure 3) [41][42]. In addition, predictive tools such as miRcode can be used to identify potential lncRNA-miRNA interactions, although it should be noted that these tools require more experimental validation [43][44]. It is still evident that there may be an underlying intricate network of mRNA-miRNA-lncRNA in disease progression that has yet to be explored. Improvements in a sensitive and live detection system for particular miRNAs such as miRDREL could also help illustrate a clearer picture of the action mechanism of oncomiRs such as miR-21 [45].
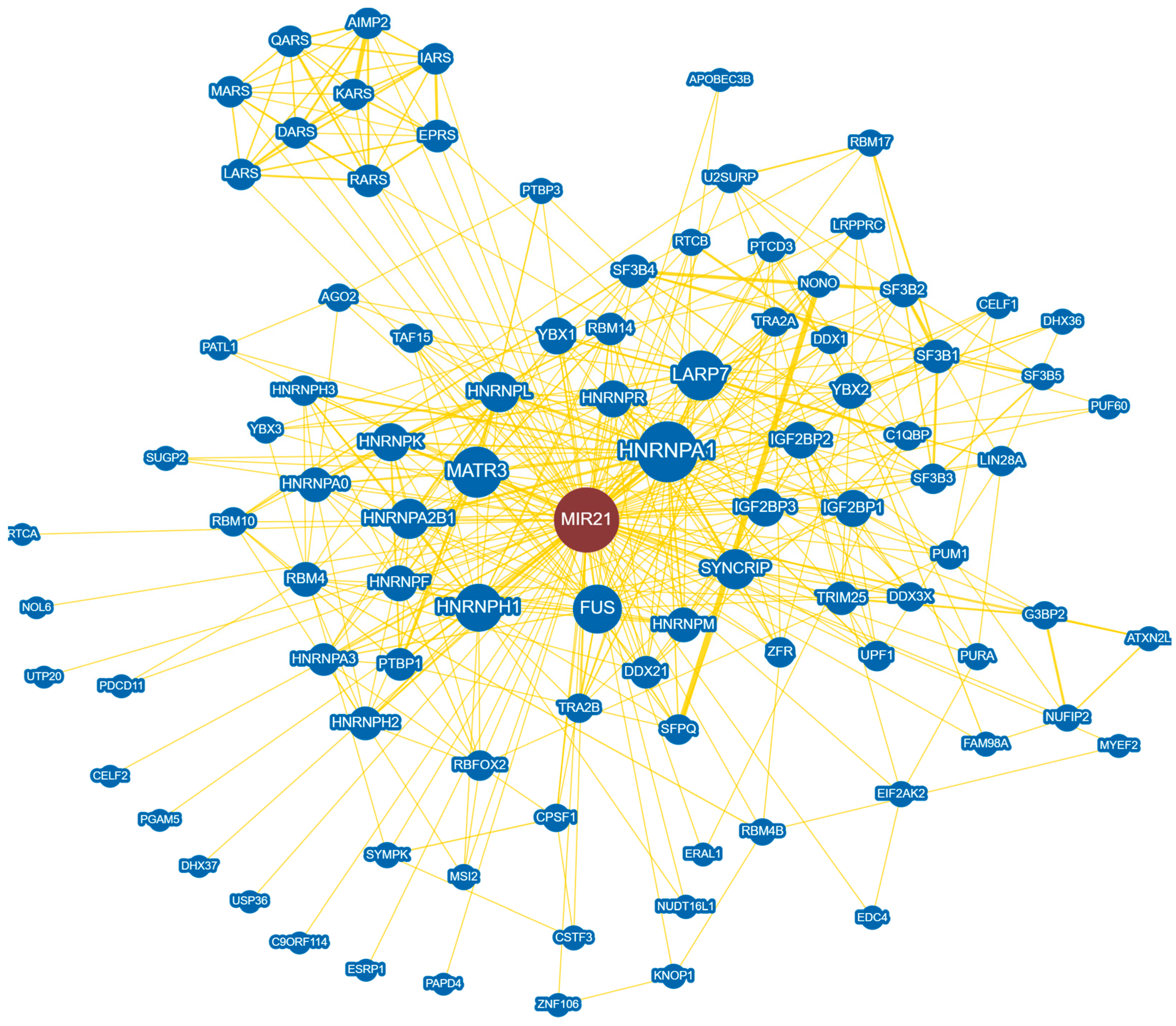
Figure 3. BioGRID protein interaction analysis of hsa-miR-21 interactome. In total, 96 protein interactions are displayed with an arbor layout in which related nodes are spaced closer together. Of the 96 protein interactors, 94 were found through high-throughput evidence, while 2 were found through low-throughput. AGO2—argonaute RISC catalytic component 2, AIMP2—aminoacyl tRNA synthetase complex-interacting multifunctional protein 2, APOBEC3B—apolipoprotein B mRNA editing enzyme catalytic subunit 3B, ATXN2L—ataxin-2-like protein, C1QBP—complement C1q binding protein, C9orf114—putative methyltransferase, CELF1/2—CUGBP Elav-like family member 1/2, CPSF1—cleavage and polyadenylation specific factor 1, CSTF3—cleavage stimulation factor subunit 3, DARS—aspartate-tRNA synthetase, DDX1/21/3X—DEAD-box helicase 1/21/3X, DHX36/37—DEAH-box helicase 36/37, EDC4—enhancer of mRNA decapping 4, EIF2AK2—eukaryotic translation initiation factor 2 alpha kinase 2, EPRS—glutamyl-prolyl-tRNA synthetase, ERAL1—Era-like 12S mitochondrial rRNA chaperone 1, ESRP1—epithelial splicing regulatory protein 1, FAM98A—family with sequence similarity 98 member A, FUS—FUS RNA binding protein, G3BP2—G3BP stress granule assembly factor 2, HNRNPA0/A1/A2B1/A3/F/H1/H2/H3/K/L/M/R—heterogeneous nuclear ribonucleoprotein A0/A1/A2B1/A3/F/H1/H2/H3/K/L/M/R, IARS—isoleucyl-tRNA synthetase, IGF2BP1/2/3—insulin-like growth factor 2 mRNA binding protein 1/2/3, KARS—lysyl-tRNA synthetase, KNOP1—lysine-rich nucleolar protein 1, LARP7—La ribonucleoprotein 7, LARS—leucyl-tRNA synthetase, LIN28A—lin-28 homologue A, LRPPRC—leucine-rich pentatricopeptide repeat containing, MARS—methionyl-tRNA synthetase, MATR3—matrin 3, MSI2—musashi RNA binding protein 2, MYEF2—myelin expression factor 2, NOL6—nucleolar protein 6, NONO—non-POU domain containing octamer binding, NUDT16L1—nudix hydrolase 16 like 1, NUFIP2—nuclear FMR1 interacting protein 2, PAPD4—PAP-associated domain-containing protein 4, PATL1—PAT1 homolog 1, PDCD11—programmed cell death 11, PGAM5—PGAM family member 5 mitochondrial serine/threonine protein phosphatase, PTBP1/3—polypyrimidine tract binding protein 1/3, PTCD3—pentatricopeptide repeat domain 3, PUF60—poly(U) binding splicing factor 60, PUM1—pumilio RNA binding family member 1, PURA—purine-rich element binding protein A, QARS—glutaminyl-tRNA synthetase, RARS—arginyl-tRNA synthetase, RBFOX2—RNA binding Fox-1 homolog 2, RBM10/14/17/4/4B—RNA binding motif protein 10/14/17/4/4B, RTCA/B—RNA 3′-terminal phosphate cyclase A/B, SF3B1/2/3/4/5—splicing factor 3b subunit 1/2/3/4/5, SFPQ—splicing factor proline/glutamine-rich, SUGP2—SURP and G-patch domain containing 2, SYMPK—symplekin, SYNCRIP—synaptotagmin binding cytoplasmic RNA interacting protein, TAF15—TATA-box binding protein associated factor 15, TRA2A/2B—transformer 2 alpha/2 beta homolog, TRIM25—tripartite motif containing 25, U2SURP—U2 snRNP-associated SURP domain containing, UPF1—UPF1 RNA helicase and ATPase, USP36—ubiquitin specific peptidase 36, UTP20—UTP20 small subunit processome component, YBX1/2/3—Y-box binding protein 1/2/3, ZFR—zinc finger RNA binding protein, and ZNF106—zinc finger protein 106.
3. MicroRNA-21-Targeted RNA Therapeutics
Various studies have explored the possibility of targeting miR-21 with RNA-mediated therapeutics such as antisense oligonucleotides (ASO), locked nucleic acid (LNA)-anti-miRNAs, and most recently synthetic or natural circRNA, which act as miRNA sponges (Table 1).
Table 1. Therapeutic approaches of miR-21 targeting in cancers.
| Type | Target Cancers | Cell Lines & Experimental Models |
Phenotypes | References |
|---|---|---|---|---|
| LNA | Colorectal adenocarcinoma |
LS174T | Proliferation inhibited; apoptosis enhanced |
[46] |
| Melanoma | B16F10; C57BL/6 mice |
Tumor growth and volume inhibited; apoptosis enhanced |
[47] | |
| Glioblastoma | U87MG; Orthotopic xenograft in athymic nude mice |
Tumor growth inhibited; apoptosis enhanced |
[48][49] | |
| Non-small cell lung cancer | A549; Female nude mice |
Drug sensitization; tumor growth inhibited; apoptosis enhanced |
[50] | |
| Breast cancer | MCF-7 | Proliferation inhibited | [23] | |
| ASO | Laryngeal squamous cell carcinoma |
Hep-2; BALB/c nude mice |
Tumor growth and proliferation inhibited; invasiveness decreased; cell cycle arrest; apoptosis enhanced |
[51] |
| Non-small cell lung cancer | PC9; Female BALB/c nude mice |
Proliferation inhibited; apoptosis enhanced; tumor growth inhibited |
[52] | |
| Glioblastoma multiforme |
LN229; U251MG; U373MG; T98G |
Drug sensitization; cell viability decreased |
[53][54] | |
| Hepatocellular carcinoma |
Huh7; HepG2 | Migration and invasiveness decreased |
[55] | |
| Breast phyllode tumor |
Patient-derived breast stromal cells; Female nude mice |
Proliferation inhibited and invasion decreased |
[56] | |
| CircRNA | Gastric carcinoma | NCI-N87; AGS; MKN28 | Proliferation inhibited | [57] |
| Lung cancer | L132, A549; LL2; 3D multicellular spheroids |
Proliferation inhibited; migration decreased; apoptosis enhanced |
[58] |
ASOs are single-stranded RNAs that are complementary to a specific target RNA, especially miRNAs [59]. In pancreatic cancer cells, delivery of ASO-miR-21 suppressed the epithelial-mesenchymal transition and hindered proliferation. Furthermore, co-delivery of ASO-miR-21 with gemcitabine, a first-line drug for the treatment of locally advanced or metastatic pancreatic cancer acted synergistically in inducing apoptosis and growth inhibition [60][61]. Various in vivo studies utilizing miR-21 ASOs have yielded tumor growth inhibition in myeloma, HCC, glioblastoma multiforme (GBM), and breast cancer, demonstrating its therapeutic potential [62][63][64][65].
LNA technology “bridges” the 2′-O and 4′-C atoms in the ribose structure of RNA nucleotides, and is thus capable of improving the hybridization stability and specificity of anti-miRNAs [66][67]. In CRC, the application of LNA-anti-miR-21 was shown to effectively inhibit growth and induce apoptosis, demonstrating the clinical potential of using LNA for miRNA-targeting therapeutics [46]. Furthermore, fully LNA-modified phosphorothioate oligonucleotides or “tiny LNAs” (8-mer-long) were reported to target the seed regions of Ago2-bound miR-21 [68].
Chu and colleagues demonstrated that this technology could be applied in vivo through the systemic administration of LNA-miR-21 inhibitors to KPC mice expected to have low-grade pancreatic intraepithelial neoplasia 1 (PanIN-1) lesions. Mice that received LNA-miR-21 did not show disease progression, which demonstrates that LNA-miR-21 inhibitors could effectively interfere with malignant progression [6].
CircRNAs have been observed to play a variety of roles such as transcriptional regulation through interaction with U1 snRNP, or functional regulation through protein interactions as in the case of circFOXO3 [69][70]. Although other linear RNAs can also act as competing endogenous RNA (ceRNA), circRNAs are advantageous in structural stability and conservation which have led to growing interest in circRNAs in therapeutic applications [71]. In addition, circRNAs are capable of acting as ceRNA through miRNA sponging [72]. MiRNA sponges have also been shown to effectively regulate miR-21 in vitro [73][74]. Recently, both synthetic and natural circRNAs were found to act as effective miRNA sponges against miR-21 [75].
In NSCLC, natural circRNA (c0001287) sponged miR-21 to upregulate PTEN and thereby inhibit proliferation and metastasis [76]. Liu and colleagues created a synthetic circRNA to sponge miR-21 and found that the inhibition of miR-21 through this method could suppress gastric carcinoma cell proliferation by downregulating death domain-associated protein DAXX [57]. Interestingly, Muller and colleagues delivered polyethyleneimine (PEI) nanoparticles containing circular miR-21-5p decoys through intraperitoneal injection in subcutaneous xenograft mice and found significant potency in inhibiting tumor growth in vivo [77]. Thus, the importance of circRNA can be shown in its therapeutic potential in cancer treatment.
Recently, Wang and colleagues developed a method to simultaneously sponge miR-21 and deliver a chemotherapeutic drug. In this system, an oligonucleotide shell of a spherical nucleic acid of gold nanoparticles loaded with doxorubicin was shown to capture miR-21/miR-155, and thereby trigger the release of doxorubicin to enable tumor-specific chemotherapy [78].
References
- Fujita, S.; Ito, T.; Mizutani, T.; Minoguchi, S.; Yamamichi, N.; Sakurai, K.; Iba, H. miR-21 Gene expression triggered by AP-1 is sustained through a double-negative feedback mechanism. J. Mol. Biol. 2008, 378, 492–504.
- Feng, Y.H.; Tsao, C.J. Emerging role of microRNA-21 in cancer. Biomed. Rep. 2016, 5, 395–402.
- Bailey, M.H.; Tokheim, C.; Porta-Pardo, E.; Sengupta, S.; Bertrand, D.; Weerasinghe, A.; Colaprico, A.; Wendl, M.C.; Kim, J.; Reardon, B.; et al. Comprehensive Characterization of Cancer Driver Genes and Mutations. Cell 2018, 173, 371–385.
- Zhou, Y.; Guo, D.; Zhang, Y. Association of MicroRNA-21 with p53 at Mutant Sites R175H and R248Q, Clinicopathological Features, and Prognosis of NSCLC. Mol. Ther. Oncolytics 2020, 19, 208–217.
- Bornachea, O.; Santos, M.; Martínez-Cruz, A.B.; García-Escudero, R.; Dueñas, M.; Costa, C.; Segrelles, C.; Lorz, C.; Buitrago, A.; Saiz-Ladera, C.; et al. EMT and induction of miR-21 mediate metastasis development in Trp53-deficient tumours. Sci. Rep. 2012, 2, 434.
- Chu, N.J.; Anders, R.A.; Fertig, E.J.; Cao, M.; Hopkins, A.C.; Keenan, B.P. Inhibition of miR-21 Regulates Mutant KRAS Effector Pathways and Intercepts Pancreatic Ductal Adenocarcinoma Development. Cancer Prev. Res. 2020, 13, 569–582.
- Hatley, M.E.; Patrick, D.M.; Garcia, M.R.; Richardson, J.A.; Bassel-Duby, R.; van Rooij, E.; Olson, E.N. Modulation of K-Ras-dependent lung tumorigenesis by MicroRNA-21. Cancer Cell 2010, 18, 282–293.
- Seike, M.; Goto, A.; Okano, T.; Bowman, E.D.; Schetter, A.J.; Horikawa, I.; Mathe, E.A.; Jen, J.; Yang, P.; Sugimura, H.; et al. MiR-21 is an EGFR-regulated anti-apoptotic factor in lung cancer in never-smokers. Proc. Natl. Acad. Sci. USA 2009, 106, 12085–12090.
- Shen, J.; Xia, W.; Khotskaya, Y.B.; Huo, L.; Nakanishi, K.; Lim, S.O.; Du, Y.; Wang, Y.; Chang, W.C.; Chen, C.H.; et al. EGFR modulates microRNA maturation in response to hypoxia through phosphorylation of AGO2. Nature 2013, 497, 383–387.
- Bloomston, M.; Frankel, W.L.; Petrocca, F.; Volinia, S.; Alder, H.; Hagan, J.P.; Liu, C.G.; Bhatt, D.; Taccioli, C.; Croce, C.M. MicroRNA expression patterns to differentiate pancreatic adenocarcinoma from normal pancreas and chronic pancreatitis. JAMA 2007, 297, 1901–1908.
- Iorio, M.V.; Ferracin, M.; Liu, C.G.; Veronese, A.; Spizzo, R.; Sabbioni, S.; Magri, E.; Pedriali, M.; Fabbri, M.; Campiglio, M.; et al. MicroRNA gene expression deregulation in human breast cancer. Cancer Res. 2005, 65, 7065–7070.
- Schetter, A.J.; Leung, S.Y.; Sohn, J.J.; Zanetti, K.A.; Bowman, E.D.; Yanaihara, N.; Yuen, S.T.; Chan, T.L.; Kwong, D.L.; Au, G.K.; et al. MicroRNA expression profiles associated with prognosis and therapeutic outcome in colon adenocarcinoma. JAMA 2008, 299, 425–436.
- Volinia, S.; Calin, G.A.; Liu, C.G.; Ambs, S.; Cimmino, A.; Petrocca, F.; Visone, R.; Iorio, M.; Roldo, C.; Ferracin, M.; et al. A microRNA expression signature of human solid tumors defines cancer gene targets. Proc. Natl. Acad. Sci. USA 2006, 103, 2257–2261.
- Landgraf, P.; Rusu, M.; Sheridan, R.; Sewer, A.; Iovino, N.; Aravin, A.; Pfeffer, S.; Rice, A.; Kamphorst, A.O.; Landthaler, M.; et al. A mammalian microRNA expression atlas based on small RNA library sequencing. Cell 2007, 129, 1401–1414.
- Ciafrè, S.A.; Galardi, S.; Mangiola, A.; Ferracin, M.; Liu, C.G.; Sabatino, G.; Negrini, M.; Maira, G.; Croce, C.M.; Farace, M.G. Extensive modulation of a set of microRNAs in primary glioblastoma. Biochem. Biophys. Res. Commun. 2005, 334, 1351–1358.
- Li, R.; Qu, H.; Wang, S. CancerMIRNome: An interactive analysis and visualization database for miRNome profiles of human cancer. Nucleic Acids Res. 2022, 50, D1139–D1146.
- Ren, Y.; Zhou, X.; Mei, M.; Yuan, X.B.; Han, L.; Wang, G.X.; Jia, Z.F.; Xu, P.; Pu, P.Y.; Kang, C.S. MicroRNA-21 inhibitor sensitizes human glioblastoma cells U251 (PTEN-mutant) and LN229 (PTEN-wild type) to taxol. BMC Cancer 2010, 10, 27.
- Han, L.; Yue, X.; Zhou, X.; Lan, F.M.; You, G.; Zhang, W.; Zhang, K.L.; Zhang, C.Z.; Cheng, J.Q.; Yu, S.Z.; et al. MicroRNA-21 expression is regulated by β-catenin/STAT3 pathway and promotes glioma cell invasion by direct targeting RECK. CNS Neurosci. Ther. 2012, 18, 573–583.
- Zhang, K.L.; Han, L.; Chen, L.Y.; Shi, Z.D.; Yang, M.; Ren, Y.; Chen, L.C.; Zhang, J.X.; Pu, P.Y.; Kang, C.S. Blockage of a miR-21/EGFR regulatory feedback loop augments anti-EGFR therapy in glioblastomas. Cancer Lett. 2014, 342, 139–149.
- Kwak, H.J.; Kim, Y.J.; Chun, K.R.; Woo, Y.M.; Park, S.J.; Jeong, J.A.; Jo, S.H.; Kim, T.H.; Min, H.S.; Chae, J.S.; et al. Downregulation of Spry2 by miR-21 triggers malignancy in human gliomas. Oncogene 2011, 30, 2433–2442.
- Zhao, Q.; Chen, S.; Zhu, Z.; Yu, L.; Ren, Y.; Jiang, M.; Weng, J. miR-21 promotes EGF-induced pancreatic cancer cell proliferation by targeting Spry2. Cell Death Dis. 2018, 9, 1157.
- Zhang, J.G.; Wang, J.J.; Zhao, F.; Liu, Q.; Jiang, K.; Yang, G.H. MicroRNA-21 (miR-21) represses tumor suppressor PTEN and promotes growth and invasion in non-small cell lung cancer (NSCLC). Clin. Chim. Acta 2010, 411, 846–852.
- Frankel, L.B.; Christoffersen, N.R.; Jacobsen, A.; Lindow, M.; Krogh, A.; Lund, A.H. Programmed cell death 4 (PDCD4) is an important functional target of the microRNA miR-21 in breast cancer cells. J. Biol. Chem. 2008, 283, 1026–1033.
- Zhu, S.; Wu, H.; Wu, F.; Nie, D.; Sheng, S.; Mo, Y.Y. MicroRNA-21 targets tumor suppressor genes in invasion and metastasis. Cell Res. 2008, 18, 350–359.
- Medina, P.P.; Nolde, M.; Slack, F.J. OncomiR addiction in an in vivo model of microRNA-21-induced pre-B-cell lymphoma. Nature 2010, 467, 86–90.
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297.
- Cortez, M.A.; Bueso-Ramos, C.; Ferdin, J.; Lopez-Berestein, G.; Sood, A.K.; Calin, G.A. MicroRNAs in body fluids--the mix of hormones and biomarkers. Nat. Rev. Clin. Oncol. 2011, 8, 467–477.
- Creemers, E.E.; Tijsen, A.J.; Pinto, Y.M. Circulating microRNAs: Novel biomarkers and extracellular communicators in cardiovascular disease? Circ. Res. 2012, 110, 483–495.
- Tomimaru, Y.; Eguchi, H.; Nagano, H.; Wada, H.; Kobayashi, S.; Marubashi, S.; Tanemura, M.; Tomokuni, A.; Takemasa, I.; Umeshita, K.; et al. Circulating microRNA-21 as a novel biomarker for hepatocellular carcinoma. J. Hepatol. 2012, 56, 167–175.
- Sierzega, M.; Kaczor, M.; Kolodziejczyk, P.; Kulig, J.; Sanak, M.; Richter, P. Evaluation of serum microRNA biomarkers for gastric cancer based on blood and tissue pools profiling: The importance of miR-21 and miR-331. Br. J. Cancer 2017, 117, 266–273.
- Guo, X.; Lv, X.; Lv, X.; Ma, Y.; Chen, L.; Chen, Y. Circulating miR-21 serves as a serum biomarker for hepatocellular carcinoma and correlated with distant metastasis. Oncotarget 2017, 8, 44050–44058.
- Qu, K.Z.; Zhang, K.; Li, H.; Afdhal, N.H.; Albitar, M. Circulating microRNAs as biomarkers for hepatocellular carcinoma. J. Clin. Gastroenterol. 2011, 45, 355–360.
- Lee, K.-Y.; Seo, Y.; Im, J.H.; Rhim, J.; Baek, W.; Kim, S.; Kwon, J.-W.; Yoo, B.C.; Shin, S.H.; Yoo, H.; et al. Molecular Signature of Extracellular Vesicular Small Non-Coding RNAs Derived from Cerebrospinal Fluid of Leptomeningeal Metastasis Patients: Functional Implication of miR-21 and Other Small RNAs in Cancer Malignancy. Cancers 2021, 13, 209.
- Im, J.H.; Yoo, B.C.; Lee, J.H.; Lee, K.-Y.; Kim, K.-H.; Kim, J.H.; Park, H.J.; Park, M.; Lee, S.H.; Kwon, J.-W.; et al. Experimental Assessment of Leptomeningeal Metastasis Diagnosis in Medulloblastoma Using Cerebrospinal Fluid Metabolomic Profiles. Metabolites 2021, 11, 851.
- Toiyama, Y.; Takahashi, M.; Hur, K.; Nagasaka, T.; Tanaka, K.; Inoue, Y.; Kusunoki, M.; Boland, C.R.; Goel, A. Serum miR-21 as a diagnostic and prognostic biomarker in colorectal cancer. J. Natl. Cancer Inst. 2013, 105, 849–859.
- Taylor, D.D.; Gercel-Taylor, C. MicroRNA signatures of tumor-derived exosomes as diagnostic biomarkers of ovarian cancer. Gynecol. Oncol. 2008, 110, 13–21.
- Lee, K.Y.; Im, J.H.; Lin, W.; Gwak, H.S. Nanoparticles in 472 Human Cerebrospinal Fluid: Changes in Extracellular Vesicle Concentration and miR-21 Expression as a Biomarker for Leptomeningeal Metastasis. Cancers 2020, 12, 2745.
- Im, J.H.; Kim, T.H.; Lee, K.-Y.; Gwak, H.-S.; Lin, W.; Park, J.B.; Kim, J.H.; Yoo, B.C.; Park, S.-M.; Kwon, J.-W.; et al. Exploratory Profiling of Extracellular MicroRNAs in Cerebrospinal Fluid Comparing Leptomeningeal Metastasis with Other Central Nervous System Tumor Statuses. J. Clin. Med. 2021, 10, 4860.
- Im, J.H.; Yoo, B.C.; Lee, J.H.; Kim, K.H.; Kim, T.H.; Lee, K.Y.; Kim, J.H.; Park, J.B.; Kwon, J.W.; Shin, S.H.; et al. Comparative cerebrospinal fluid metabolites profiling in glioma patients to predict malignant transformation and leptomeningeal metastasis with a potential for preventive personalized medicine. EPMA J. 2020, 11, 469–484.
- Li, J.R.; Tong, C.Y.; Sung, T.J.; Kang, T.Y.; Zhou, X.J.; Liu, C.C. CMEP: A database for circulating microRNA expression profiling. Bioinformatics 2019, 35, 3127–3132.
- Oughtred, R.; Rust, J.; Chang, C.; Breitkreutz, B.J.; Stark, C.; Willems, A.; Boucher, L.; Leung, G.; Kolas, N.; Zhang, F.; et al. The BioGRID database: A comprehensive biomedical resource of curated protein, genetic, and chemical interactions. Protein Sci. 2021, 30, 187–200.
- Stark, C.; Breitkreutz, B.J.; Reguly, T.; Boucher, L.; Breitkreutz, A.; Tyers, M. BioGRID: A general repository for interaction datasets. Nucleic Acids Res. 2006, 34, D535–D539.
- López-Urrutia, E.; Bustamante Montes, L.P.; Ladrón de Guevara Cervantes, D.; Pérez-Plasencia, C.; Campos-Parra, A.D. Crosstalk Between Long Non-coding RNAs, Micro-RNAs and mRNAs: Deciphering Molecular Mechanisms of Master Regulators in Cancer. Front. Oncol. 2019, 9, 669.
- Jeggari, A.; Marks, D.S.; Larsson, E. miRcode: A map of putative microRNA target sites in the long non-coding transcriptome. Bioinformatics 2012, 28, 2062–2063.
- Seo, Y.; Kim, S.S.; Kim, N.; Cho, S. Development of a miRNA-controlled dual-sensing system and its application for targeting miR-21 signaling in tumorigenesis. Exp. Mol. Med. 2020, 52, 1989–2004.
- Nedaeinia, R.; Sharifi, M. Locked nucleic acid anti-miR-21 inhibits cell growth and invasive behaviors of a colorectal adenocarcinoma cell line: LNA-anti-miR as a novel approach. Cancer Gene Ther. 2016, 23, 246–253.
- Javanmard, S.H.; Vaseghi, G.; Ghasemi, A.; Rafiee, L.; Ferns, G.A.; Esfahani, H.N.; Nedaeinia, R. Therapeutic inhibition of microRNA-21 (miR-21) using locked-nucleic acid (LNA)-anti-miR and its effects on the biological behaviors of melanoma cancer cells in preclinical studies. Cancer Cell Int. 2020, 20, 384.
- Lee, T.J.; Yoo, J.Y.; Shu, D.; Li, H.; Zhang, J.; Yu, J.G.; Jaime-Ramirez, A.C.; Acunzo, M.; Romano, G.; Cui, R.; et al. RNA Nanoparticle-Based Targeted Therapy for Glioblastoma through Inhibition of Oncogenic miR-21. Mol. Ther. 2017, 25, 1544–1555.
- Griveau, A.; Bejaud, J.; Anthiya, S.; Avril, S.; Autret, D.; Garcion, E. Silencing of miR-21 by locked nucleic acid-lipid nanocapsule complexes sensitize human glioblastoma cells to radiation-induced cell death. Int. J. Pharm. 2013, 454, 765–774.
- Xu, L.; Huang, Y.; Chen, D.; He, J.; Zhu, W.; Zhang, Y.; Liu, X. Downregulation of miR-21 increases cisplatin sensitivity of non-small-cell lung cancer. Cancer Genet. 2014, 207, 214–220.
- Ren, J.; Zhu, D.; Liu, M.; Sun, Y.; Tian, L. Downregulation of miR-21 modulates Ras expression to promote apoptosis and suppress invasion of Laryngeal squamous cell carcinoma. Eur. J. Cancer 2010, 46, 3409–3416.
- Li, B.; Ren, S.; Li, X.; Wang, Y.; Garfield, D.; Zhou, S.; Chen, X.; Su, C.; Chen, M.; Kuang, P.; et al. MiR-21 overexpression is associated with acquired resistance of EGFR-TKI in non-small cell lung cancer. Lung Cancer 2014, 83, 146–153.
- Li, Y.; Li, W.; Yang, Y.; Lu, Y.; He, C.; Hu, G.; Liu, H.; Chen, J.; He, J.; Yu, H. MicroRNA-21 targets LRRFIP1 and contributes to VM-26 resistance in glioblastoma multiforme. Brain Res. 2009, 1286, 13–18.
- Giunti, L.; da Ros, M.; Vinci, S.; Gelmini, S.; Iorio, A.L.; Buccoliero, A.M.; Cardellicchio, S.; Castiglione, F.; Genitori, L.; de Martino, M.; et al. Anti-miR21 oligonucleotide enhances chemosensitivity of T98G cell line to doxorubicin by inducing apoptosis. Am. J. Cancer Res. 2015, 5, 231–242.
- Connolly, E.C.; Van Doorslaer, K.; Rogler, L.E.; Rogler, C.E. Overexpression of miR-21 promotes an in vitro metastatic phenotype by targeting the tumor suppressor RHOB. Mol. Cancer Res. 2010, 8, 691–700.
- Gong, C.; Nie, Y.; Qu, S.; Liao, J.Y.; Cui, X.; Yao, H.; Zeng, Y.; Su, F.; Song, E.; Liu, Q. miR-21 induces myofibroblast differentiation and promotes the malignant progression of breast phyllodes tumors. Cancer Res. 2014, 74, 4341–4352.
- Liu, X.; Abraham, J.M.; Cheng, Y.; Wang, Z.; Wang, Z.; Zhang, G.; Ashktorab, H.; Smoot, D.T.; Cole, R.N.; Boronina, T.N.; et al. Synthetic Circular RNA Functions as a miR-21 Sponge to Suppress Gastric Carcinoma Cell Proliferation. Mol. Ther. Nucleic Acids 2018, 13, 312–321.
- Rama, A.R.; Quiñonero, F. Synthetic Circular miR-21 Sponge as Tool for Lung Cancer Treatment. Int. J. Mol. Sci. 2022, 23, 2963.
- Davis, S.; Lollo, B.; Freier, S.; Esau, C. Improved targeting of miRNA with antisense oligonucleotides. Nucleic Acids Res. 2006, 34, 2294–2304.
- Burris, H.A., 3rd; Moore, M.J.; Andersen, J.; Green, M.R.; Rothenberg, M.L.; Modiano, M.R.; Cripps, M.C.; Portenoy, R.K.; Storniolo, A.M.; Tarassoff, P.; et al. Improvements in survival and clinical benefit with gemcitabine as first-line therapy for patients with advanced pancreas cancer: A randomized trial. J. Clin. Oncol. 1997, 15, 2403–2413.
- Li, Y.; Chen, Y.; Li, J.; Zhang, Z.; Huang, C.; Lian, G.; Yang, K.; Chen, S.; Lin, Y.; Wang, L.; et al. Co-delivery of microRNA-21 antisense oligonucleotides and gemcitabine using nanomedicine for pancreatic cancer therapy. Cancer Sci. 2017, 108, 1493–1503.
- Si, M.L.; Zhu, S.; Wu, H.; Lu, Z.; Wu, F.; Mo, Y.Y. miR-21-mediated tumor growth. Oncogene 2007, 26, 2799–2803.
- Leone, E.; Morelli, E.; Di Martino, M.T.; Amodio, N.; Foresta, U.; Gullà, A.; Rossi, M.; Neri, A.; Giordano, A.; Munshi, N.C.; et al. Targeting miR-21 inhibits in vitro and in vivo multiple myeloma cell growth. Clin. Cancer Res. 2013, 19, 2096–2106.
- Wagenaar, T.R.; Tolstykh, T.; Shi, C.; Jiang, L.; Zhang, J.; Li, Z.; Yu, Q.; Qu, H.; Sun, F.; Cao, H.; et al. Identification of the endosomal sorting complex required for transport-I (ESCRT-I) as an important modulator of anti-miR uptake by cancer cells. Nucleic Acids Res. 2015, 43, 1204–1215.
- Zhou, X.; Zhang, J.; Jia, Q.; Ren, Y.; Wang, Y.; Shi, L.; Liu, N.; Wang, G.; Pu, P.; You, Y.; et al. Reduction of miR-21 induces glioma cell apoptosis via activating caspase 9 and 3. Oncol. Rep. 2010, 24, 195–201.
- Koshkin, A.A.; Singh, S.K.; Nielsen, P.; Rajwanshi, V.K.; Kumar, R.; Meldgaard, M.; Olsen, C.E.; Wengel, J. LNA (Locked Nucleic Acids): Synthesis of the adenine, cytosine, guanine, 5-methylcytosine, thymine and uracil bicyclonucleoside monomers, oligomerisation, and unprecedented nucleic acid recognition. Tetrahedron 1998, 54, 3607–3630.
- Lima, J.F.; Cerqueira, L. Anti-miRNA oligonucleotides: A comprehensive guide for design. RNA Biol. 2018, 15, 338–352.
- Obad, S.; dos Santos, C.O.; Petri, A.; Heidenblad, M.; Broom, O.; Ruse, C.; Fu, C.; Lindow, M.; Stenvang, J.; Straarup, E.M.; et al. Silencing of microRNA families by seed-targeting tiny LNAs. Nat. Genet. 2011, 43, 371–378.
- Du, W.W.; Zhang, C.; Yang, W.; Yong, T.; Awan, F.M.; Yang, B.B. Identifying and Characterizing circRNA-Protein Interaction. Theranostics 2017, 7, 4183–4191.
- Li, Z.; Huang, C.; Bao, C.; Chen, L.; Lin, M.; Wang, X.; Zhong, G.; Yu, B.; Hu, W.; Dai, L.; et al. Exon-intron circular RNAs regulate transcription in the nucleus. Nat. Struct. Mol. Biol. 2015, 22, 256–264.
- Zhong, Y.; Du, Y.; Yang, X.; Mo, Y.; Fan, C.; Xiong, F.; Ren, D.; Ye, X.; Li, C.; Wang, Y.; et al. Circular RNAs function as ceRNAs to regulate and control human cancer progression. Mol. Cancer 2018, 17, 79.
- Anastasiadou, E.; Jacob, L.S.; Slack, F.J. Non-coding RNA networks in cancer. Nat. Rev. Cancer 2018, 18, 5–18.
- Ebert, M.S.; Neilson, J.R.; Sharp, P.A. MicroRNA sponges: Competitive inhibitors of small RNAs in mammalian cells. Nat. Methods 2007, 4, 721–726.
- Jung, J.; Yeom, C.; Choi, Y.S.; Kim, S.; Lee, E.; Park, M.J.; Kang, S.W.; Kim, S.B.; Chang, S. Simultaneous inhibition of multiple oncogenic miRNAs by a multi-potent microRNA sponge. Oncotarget 2015, 6, 20370–20387.
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388.
- Zhang, C.C.; Li, Y.; Feng, X.Z.; Li, D.B. Circular RNA circ_0001287 inhibits the proliferation, metastasis, and radiosensitivity of non-small cell lung cancer cells by sponging microRNA miR-21 and up-regulating phosphatase and tensin homolog expression. Bioengineered 2021, 12, 414–425.
- Müller, S.; Wedler, A.; Breuer, J.; Glaß, M.; Bley, N.; Lederer, M.; Haase, J.; Misiak, C.; Fuchs, T.; Ottmann, A.; et al. Synthetic circular miR-21 RNA decoys enhance tumor suppressor expression and impair tumor growth in mice. NAR Cancer 2020, 2, zcaa014.
- Wang, X.; Yang, T.; Yu, Z.; Liu, T.; Jin, R.; Weng, L.; Bai, Y.; Gooding, J.J.; Zhang, Y.; Chen, X. Intelligent Gold Nanoparticles with Oncogenic MicroRNA-Dependent Activities to Manipulate Tumorigenic Environments for Synergistic Tumor Therapy. Adv. Mater. 2022, 34, e2110219.
More
Information
Subjects:
Biochemistry & Molecular Biology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
2.1K
Revisions:
2 times
(View History)
Update Date:
21 Sep 2022
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No