Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Jacques Christian Mbongue | -- | 2395 | 2022-08-24 17:15:06 | | | |
| 2 | Rita Xu | -21 word(s) | 2374 | 2022-08-25 07:17:36 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Mbongue, J.C.; Vanterpool, E.; Firek, A.; Langridge, W.H.R. Bacterial Lipopolysaccharides. Encyclopedia. Available online: https://encyclopedia.pub/entry/26453 (accessed on 28 July 2026).
Mbongue JC, Vanterpool E, Firek A, Langridge WHR. Bacterial Lipopolysaccharides. Encyclopedia. Available at: https://encyclopedia.pub/entry/26453. Accessed July 28, 2026.
Mbongue, Jacques C., Elaine Vanterpool, Anthony Firek, William H. R. Langridge. "Bacterial Lipopolysaccharides" Encyclopedia, https://encyclopedia.pub/entry/26453 (accessed July 28, 2026).
Mbongue, J.C., Vanterpool, E., Firek, A., & Langridge, W.H.R. (2022, August 24). Bacterial Lipopolysaccharides. In Encyclopedia. https://encyclopedia.pub/entry/26453
Mbongue, Jacques C., et al. "Bacterial Lipopolysaccharides." Encyclopedia. Web. 24 August, 2022.
Copy Citation
Bacterial lipopolysaccharides (LPS), also referred to as endotoxins, are major outer surface membrane components present on almost all Gram-negative bacteria and are major determinants of sepsis-related clinical complications including septic shock. LPS acts as a strong stimulator of innate or natural immunity in a wide variety of eukaryotic species ranging from insects to humans including specific effects on the adaptive immune system.
immunological tolerance
LPS
cholera toxin B
Indoleamine 2,3 dioxygenase
1. Introduction
Understanding the role of LPS in both the toxicity of bacterial infections and effects on immune regulation has emerged as a critical objective due to the global importance of sepsis. The LPS bacterial surface molecule is produced by most Gram-negative bacteria. The attention LPS received in the early 20th century was due to its ability to stimulate the immune system, and was known as endotoxin glycolipids [1]. It was subsequently discovered that LPS created a permeable barrier on the cell surface and was the main driver of the innate resistance of Gram-negative bacteria to many antibacterial agents [1][2][3][4]. Unsurprisingly, these important properties of LPS have provided a vast and extensive literature for over 100 years.
The innate immune system’s detection of microorganisms or microbial components is mediated by a special set of proteins called pattern recognition receptors (PRRs). One of the best studied PRRs is the bacterial LPS receptor, Toll-like receptor 4 (TLR4) [5][6]. TLR4 is an important driver of the immune response to bacterial infections and its dysregulation is thought to promote abnormal cytokine production, leading to bacterial sepsis [3][4][7][8]. Because sepsis remains one of the major conditions leading to acute morbidity and mortality, understanding the nature of TLR4 signaling will direct efforts towards understanding the basic mechanisms underlying inflammation and can lead to improved clinical outcome. Bacterial LPS is widely used in inflammation models because it induces many inflammatory effects by promoting the production and release of pro-inflammatory cytokines such as TNF-α, IL-1β, and IL-6 [8][9][10].
LPS signaling by PRR leads to the activation of intracellular signaling networks that promote the expression of inflammatory genes which stimulate the acute and sustained defense of the host [6][7]. TLR4 first encounters LPS in extracellular space when interacting with intact bacteria or when exposed to soluble LPS aggregates. Upon binding to LPS, TLR4 rapidly induces the assembly of supramolecular organization centers (SMOC) called myddosomes [11].
The intermediate consists of the MyD88 adapter protein and several serine-threonine kinases from the TIRAP and IRAK families [5][6][7][8]. This hub-like organizing center is a major subcellular site where TLR4 signaling activates the NF-κB and AP-1 pathways to promote the expression of inflammatory genes [12][13][14]. Subsequently, TLR4 is taken up by endosomes and promotes the production of IRF3-dependent interferon type I (IFN) via TRAM and TRIF adaptor proteins [15][16][17].
LPS acts as a proto-endotoxin and contributes to the inflammatory cascade because it binds to the CD14/TLR4/MD2 receptor complex in many cell types, but primarily to monocytes, dendritic cells, macrophages, and B cells. In these cell types, LPS stimulates the secretion of proinflammatory cytokines, eicosanoids, and nitric oxide [1][7]. Due to its role in the activation of several transcription factors, LPS activity has been experimentally investigated for many years [1][2][7][18].
Humans are more sensitive to LPS than other animals (e.g., mice). An LPS dose of 1 mcg/kg causes shock in humans, but mice can tolerate doses up to 1000 times that amount [19]. For this reason, LPS levels in pharmaceutical products and medical devices must be strictly monitored using the limulus amebocyte lysate (LAL) assay. This requirement may be due to differences in the amount of circulating natural anti-LPS antibodies between the two species [20][21].
Multiple pathways may be activated on engagement with LPS because it has been shown that LPS has both an immunostimulatory as well as immunosuppressive roles to play in immune activation of immune cells [22][23]. Consequently, despite LPS being recognized as a classical immune stimulating factor best characterized in bacterial infection, there is clear evidence of a far more complex role for LPS in the immune cascade.
Dendritic cells (DCs), considered the most potent APCs, are critical gateways of the immune system and have the unique ability to synthesize a wide range of input signals and transmit them to naive lymphocytes, thus directing immunization, or suppression of pathogenic microorganisms and tumors [24]. When confronted with pathogen-associated molecular structures, DCs “mature” by upregulating the expression of MHC class II receptors that exhibit antigen, cofactors, and processed cytokines and chemokines [25]. DCs contain TLRs which are major pattern recognition receptors that initiate and regulate immune responses via various signaling pathways [26]. Thus, the application and targeted regulation of DCs to control cancer and infectious diseases is being pursued in the development of clinical therapeutics [27]. However, DCs are also involved in the pathogenesis of diseases caused by immune cell dysfunction, such as chronic inflammation, autoimmunity, and cancer development and progression [28][29]. Uptake processing and presentation of self-antigens as foreign proteins is considered fundamental to the development of autoimmune conditions such as type 1 diabetes. Thus, targeting the downregulation of DC activation may be a useful strategy for treatment of these diseases.
Monocytes are also important in the early acute inflammatory phase of the immune response to an infectious agent because they can stimulate and modulate the adaptive immune system by inducing cytokine secretion and antigen presentation to T cells [30][31]. The adaptive immune response is complimentary to the innate immune response and these two processes can simultaneously eliminate pathogens. In most cases, monocytes initiate and enhance the immune response. However, LPS activation of monocytes has been shown to suppress the T cell immune response and induce the expression of FOXP3 regulatory T cell function modulators in resting CD4 + CD25 T cells via a PGE2-dependent mechanism [32].
LPS also plays an immunosuppressive role in autoimmunity [22]. For example, repeated exposure to LPS causes a state of endotoxin tolerance that, in part, contributes to the well-recognized state of immunosuppression seen in sepsis [33][34]. This effect is particularly important in antigen presenting cells such as monocytes [35] and dendritic cells [27][36][37].
Furthermore, exposure of bone marrow-derived dendritic cells (BMDC) to high doses of pure lipopolysaccharide for 24 h (LPS-primed BMDC) increases their potency in the prevention of inter-photoreceptor retinoid binding protein in Freund’s adjuvant-induced experimental autoimmune uveoretinitis (EAU) [38].
The concept that exposure to LPS is important for interacting with the immune system to prevent allergic and autoimmune diseases has a long history. Strachan proposed the hygiene hypothesis in 1989, and since then there have been many studies indicating differences in endotoxin levels in different habitats. These studies showed an association between LPS in house dust and the incidence of asthma. Children in rural areas have significantly fewer autoimmune diseases, such as type 1 diabetes, asthma, allergies, and generalized atopy, compared to children in urban environments, but these observations are largely descriptive [39]. Children who grew up on farms had lower rates of allergies and asthma, and dust-contaminated bedding and mattresses in their homes contained higher levels of LPS [39], a result suggesting that chronic environmental LPS exposure can promote immunotolerance to environmental antigens.
Early studies in animal models showed that the increased sensitivity of C3H/HeJ mice to food allergens was due to an inability to signal through TLR4 [40]. Neonatal administration of a cocktail of broad-spectrum antibiotics induced a food allergy response in TLR4-sufficient mice—similar to those seen in TLR4-deficient mice—which identifies the intestinal microbiota as a source of TLR4 ligand [40]. The authors of this new study confirm that several mouse model studies have shown that NOD mice that are sensitive to T1D are protected against the disease by oral or intraperitoneal administration of LPS [41]. Recent developments have revealed the mechanisms underlying adjuvant stimulated fusion protein vaccines such as the cholera toxin B subunit adjuvant linked to autoantigens like proinsulin (CTB-INS) for the protection against autoimmunity. It was shown that CTB-INS impedes human monocyte-derived DC (moDC) activation through stimulation of indoleamine 2,3 dioxygenase (IDO1) biosynthesis [42][43]. The resulting state of DC tolerance was enhanced by the residual presence of E. coli lipopolysaccharide (LPS) in partially purified CTB-INS preparations [37]. This adjuvant-like action for LPS is now recognized in vaccine development. However, the toxicity of LPS may limit its use [44][45].
2. Function of LPS
2.1. Virulence and Toxicity
Lipid A, which is the toxic component of LPS, and polysaccharide side chains, which are considered the non-toxic but immunogenic part of LPS, act as virulence determinants in Gram-negative bacteria [46][47][48]. O antigens have adhesive properties, phagocyte resistance, antigen protection, and antigen mutation properties [47][49]. Lipid A acts as an immunostimulant that induces biological responses to a specific organism [50][51][52].
2.2. Biological Activity of Lipopolysaccharide
An animal’s biological immune responses can be analyzed using various parameters, such as an injection of live or killed Gram-negative cells or purified LPS in laboratory animals, which causes a broad spectrum of pathophysiological responses, such as fever, changes in blood counts, disseminated intravascular coagulation white blood cells, hypotension, and shock resulting in death. Injecting very small doses of endotoxin can cause death in most mammals. The sequence of events follows a regular pattern: (1) latency period; (2) physiological stress (diarrhea, exhaustion, shock); and (3) death. The rate at which death occurs depends on the dose of the endotoxin, the route of administration of the toxin, and the animal species.
3. Lipopolysaccharide Signaling and Immune Activation Mechanisms in Higher Organisms
3.1. Lipopolysaccharide Detoxification Mechanisms in Higher Animals
The defense against infection in vertebrates is mediated by two interdependent arms of the immune system, known as innate and adaptive portions of the immune system. The innate immune system, consisting of antigen presenting cells, recognizes a diverse array of non-self-antigens and if overwhelmed, can signal and activate the adaptive immune system through well-established signaling pathways to stimulate an array of T-cells and B-cells to overcome the pathogen [53]. As LPS can have significant adverse effects on animals and humans, a process to detoxify LPS has been developed [54]. The detoxification mechanism of LPS occurs through enzymatic degradation or through complement-mediated detoxification, which leads to the breakdown of LPS.
3.2. Host-Microbe Interactions (Lipopolysaccharide Activity) in Invertebrates—Insects
The innate immune system of insects plays an important role in the development of immunity [55]. In recent years, arthropods and insects have become the most useful models for describing the molecular regulation of the innate immune response [56]. Insects have highly effective defense mechanisms against invasive microorganisms, which include Gram-negative and Gram-positive molecules, LPS, and peptidoglycan [55][56][57].
These insect defense mechanisms include cellular and humoral responses. Cellular responses include phagocytosis and/or encapsulation of large parasites by bacterial nodules and blood cells [58]. In addition, the humoral response uses various antimicrobial peptides which are synthesized in the adipose body and some hemocytes after induction by septic lesions and which are then secreted into the hemolymph [59][60][61]. The insect defense system against LPS pathogens results in a transient increase in antimicrobial activity in the acellular hemolymph, including phagocytosis and encapsulation of invaders by blood cells and subsequent production of antimicrobial proteins (mainly in the insect’s adipose tissue) [62]. Strong immunoreactivity was found in the interaction between Galleria mellonella (large wax moth) and LPS. The high tolerance of LPS to insects can be explained by an extremely effective detoxification mechanism involving the binding of LPS to hemolymph lipophorins [63]. This observation suggests that LPS has the potential to induce immune activation.
Activation of the proteolytic cascade and coagulation cascade using LPS triggers the limulus hemocyte to act as a signaling mechanism [64][65][66]. In addition, a blood cell membrane receptor for LPS has been isolated from Bombyx mori silkworm that can transmit an activation signal for the synthesis of the antibacterial peptide cecropin B [67][68][69].
3.3. Expression of Genes and Signaling Action Induced by Lipopolysaccharide in Vertebrates and Invertebrates
It is a general phenomenon that antibacterial protein gene expression culminates a few hours after bacterial infection and decreases over time in vertebrates. This reduction in antibacterial protein gene expression has been shown to correlate with LPS deprivation [70][71]. TLRs, a class of pattern recognition receptors (PRRs) found in vertebrates, play an important role not only in initiating innate immunity, but also in activating adaptive immunity (Figure 1).
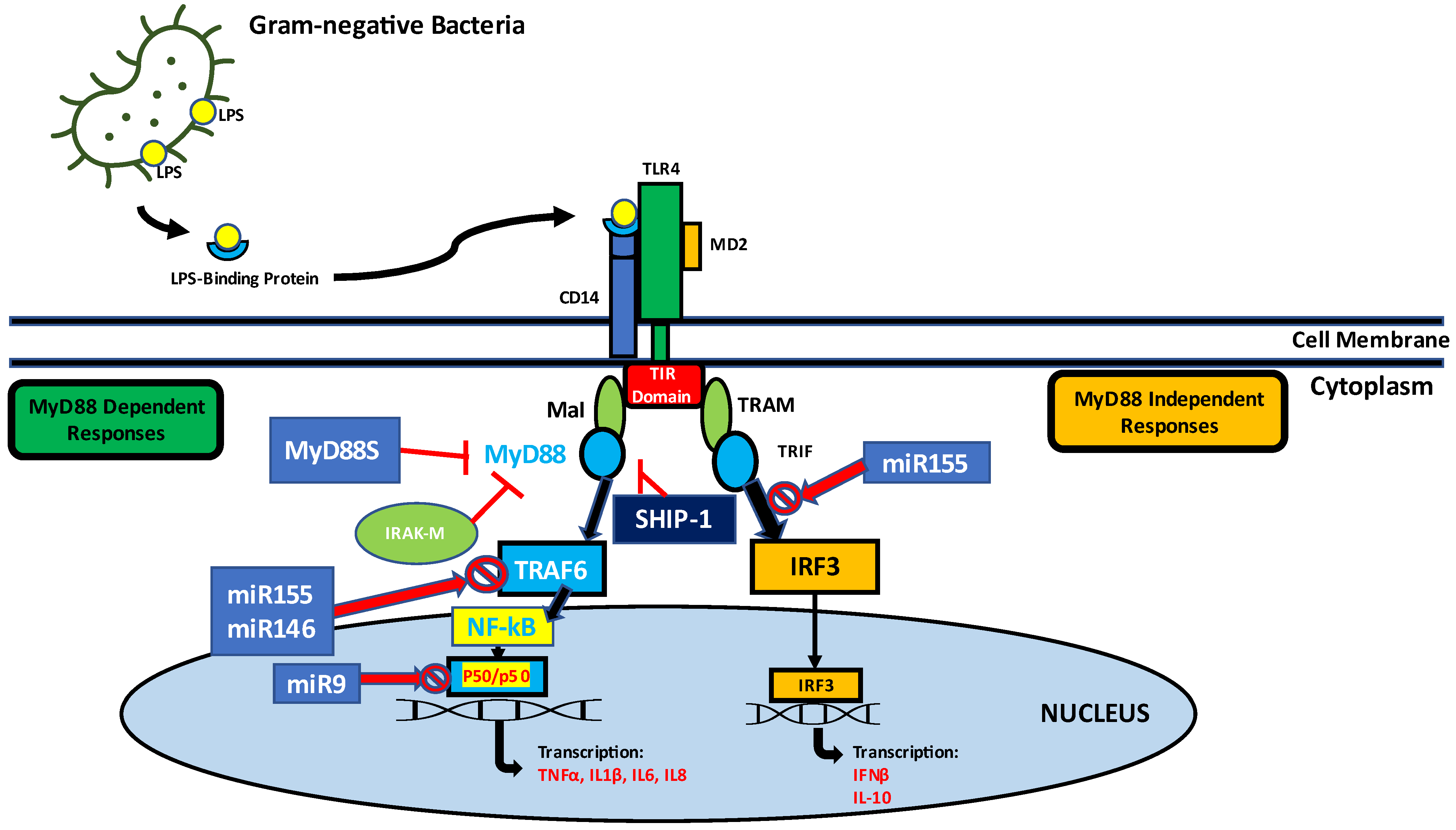
Figure 1. Endotoxin tolerance (ET) is defined as the reduced ability of cells to respond after exposure to endotoxin. The body becomes tolerant to subsequent attacks by lethal doses of endotoxin, and cytokine release and cell/tissue damage due to inflammatory responses are greatly reduced in endotoxin-resistant conditions. Key features of endotoxin tolerance are down-regulation of inflammatory mediators such as tumor necrosis factor (TNF-α) and interleukin-1β (IL-1β), upregulation of pro-inflammatory cytokines such as IL-10 and increased transforming growth factor (TGF-β). Alternative variants of MyD88S lacking an intermediate domain (ID) show a predominantly negative effect of suppressing the immune response by suppressing the MyD88 pathway. Tolerance mediators such as MyD88s, IRAK-M, and SHIP-1 are upregulated. In the endotoxin resistance phase, three LPS-inducible miRNAs (miR-155, miR-146α, and miR-9), were shown to mediate gene colocalization and transcription factor binding by blocking the binding of NF-kB p50 dimers and IRF3 as well as TRAF6-dependent NF-kB pathway, which contributes to the regulation of tolerance to endotoxins.
LPS non-specific recognition is well documented and reported in insects [72][73]. The signaling and triggering mechanism was identified in Bombyx mori [74][75]. LPS-increased levels of mRNAs were observed in A. gambiae mosquitoes containing Plasmodium berguei-infected abdomen contents [76][77]. LPS and β-1,3-glucan binding protein (LGBP) were isolated from Litopenaeus vannamei in which mRNA expression was induced by the challenge of the bacterium Vibrio alginolyticus [78][79].
Vertebrates have been shown to have acquired immunity with ‘immunological memory’, while invertebrates seem to lack this part of the adaptive immune system [80]. However, they have innate immunity, which is characterized by specific immune reactions against foreign antigens. Invertebrates are the most effective defense mechanisms of the cellular and humoral response against bacterial infections [57]. Mainly humoral reactions produce antimicrobial peptides to destroy pathogens [59][60] and this is followed by phagocytosis and nodule formation reactions as immediate defense responses to infection [81].
In addition, cellular defense reactions have been reported in invertebrates [82][83][84]. However, compared to cellular responses, humoral responses play an important role in immune defense. Insects do not appear to have an adaptive immune response that acts similarly to the well-documented antibody or the histocompatibility adaptive immune response of vertebrates [55]. In invertebrate immunity, LPS plays a role in the early stages of signaling that activates acute phase protein genes. In particular, future research on immune surveillance and purification of pathogens in vertebrates and invertebrates could demonstrate the efficacy of innate immune systems based on bacterial endotoxins.
References
- Bertani, B.; Ruiz, N. Function and Biogenesis of Lipopolysaccharides. EcoSal Plus 2018, 8.
- Alexander, T.E.; Smith, I.M.; Lipsky, Z.W.; Lozeau, L.D.; Camesano, T.A. Role of lipopolysaccharides and lipoteichoic acids on C-Chrysophsin-1 interactions with model Gram-positive and Gram-negative bacterial membranes. Biointerphases 2020, 15, 031007.
- Brandenburg, K.; Schromm, A.B.; Weindl, G.; Heinbockel, L.; Correa, W.; Mauss, K.; de Tejada, G.M.; Garidel, P. An update on endotoxin neutralization strategies in Gram-negative bacterial infections. Expert Rev. Anti-Infect. Ther. 2020, 19, 495–517.
- Giordano, N.P.; Cian, M.B.; Dalebroux, Z.D. Outer Membrane Lipid Secretion and the Innate Immune Response to Gram-Negative Bacteria. Infect. Immun. 2020, 88, e00920-19.
- Poltorak, A.; He, X.; Smirnova, I.; Liu, M.-Y.; Van Huffel, C.; Du, X.; Birdwell, D.; Alejos, E.; Silva, M.; Galanos, C.; et al. Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice: Mutations in Tlr4 gene. Science 1998, 282, 2085–2088.
- Iwasaki, A.; Medzhitov, R. Toll-like receptor control of the adaptive immune responses. Nat. Immunol. 2004, 5, 987–995.
- Rosadini, C.V.; Kagan, J.C. Early innate immune responses to bacterial LPS. Curr. Opin. Immunol. 2016, 44, 14–19.
- Tan, Y.; Kagan, J.C. A Cross-Disciplinary perspective on the innate immune responses to bacterial lipopolysaccharide. Mol. Cell 2014, 54, 212–223.
- Kent, L.W.; Rahemtulla, F.; Hockett, R.D.; Gilleland, R.C.; Michalek, S.M. Effect of lipopolysaccharide and inflammatory cytokines on Interleukin-6 production by healthy human gingival fibroblasts. Infect. Immun. 1998, 66, 608–614.
- Choi, J.; Moon, S.; Bae, H.; Kim, Y.-W.; Lee, D.; Kim, S.; Seo, Y.; Wang, H.S.; Choi, Y.W.; Lee, M.W.; et al. Alnus Sibirica Extracts Suppress the Expression of Inflammatory Cytokines Induced by Lipopolysaccharides, Tumor Necrosis Factor-α, and Interferon-γ in Human Dermal Fibroblasts. Molecules 2019, 24, 2883.
- Bonham, K.; Orzalli, M.H.; Hayashi, K.; Wolf, A.I.; Glanemann, C.; Weninger, W.; Iwasaki, A.; Knipe, D.M.; Kagan, J.C. A promiscuous lipid-binding protein diversifies the subcellular sites of toll-like receptor signal transduction. Cell 2014, 156, 705–716.
- Lai, J.; Ge, M.; Shen, S.; Yang, L.; Jin, T.; Cao, D.; Xu, H.; Zheng, X.; Qiu, S.; Wang, K.; et al. Activation of NFKB-JMJD3 signaling promotes bladder fibrosis via boosting bladder smooth muscle cell proliferation and collagen accumulation. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2019, 1865, 2403–2410.
- Min, Y.; Kim, M.-J.; Lee, S.; Chun, E.; Lee, K.-Y. Inhibition of TRAF6 ubiquitin-ligase activity by PRDX1 leads to inhibition of NFKB activation and autophagy activation. Autophagy 2018, 14, 1347–1358.
- Wang, P.; Zhou, S.; Ge, Y.; Lu, M.; Liu, Z.; Gong, R. Valproate hampers podocyte acquisition of immune phenotypes via intercepting the GSK3β facilitated NFkB activation. Oncotarget 2017, 8, 88332–88344.
- Van Acker, T.; Eyckerman, S.; Walle, L.V.; Gerlo, S.; Goethals, M.; Lamkanfi, M.; Bovijn, C.; Tavernier, J.; Peelman, F. The small GTPase Arf6 is essential for the Tram/Trif pathway in TLR4 signaling. J. Biol. Chem. 2014, 289, 1364–1376.
- Wang, Y.; Yang, Y.; Liu, X.; Wang, N.; Cao, H.; Lu, Y.; Zhou, H.; Zheng, J. Inhibition of clathrin/dynamin-dependent internalization interferes with LPS-mediated TRAM–TRIF-dependent signaling pathway. Cell. Immunol. 2012, 274, 121–129.
- Zhang, S.; Yuquan, W.; Guo, Q.; Li, R.; Li, G.-B.; Tan, S.; Li, X.; Wei, Y.; Wu, M. Annexin A2 binds to endosomes and negatively regulates TLR4-triggered inflammatory responses via the TRAM-TRIF pathway. Sci. Rep. 2015, 5, 15859.
- Hilliard, A.; Mendonca, P.; Soliman, K.F. Involvement of NFƙB and MAPK signaling pathways in the preventive effects of Ganoderma lucidum on the inflammation of BV-2 microglial cells induced by LPS. J. Neuroimmunol. 2020, 345, 577269.
- Warren, H.S.; Fitting, C.; Hoff, E.; Adib-Conquy, M.; Beasley-Topliffe, L.; Tesini, B.; Liang, X.; Valentine, C.; Hellman, J.; Hayden, D.; et al. Resilience to bacterial infection: Difference between species could be due to proteins in serum. J. Infect. Dis. 2010, 201, 223–232.
- Reid, R.R.; Prodeus, A.P.; Khan, W.; Hsu, T.; Rosen, F.S.; Carroll, M.C. Endotoxin shock in antibody-deficient mice: Unraveling the role of natural antibody and complement in the clearance of lipopolysaccharide. J. Immunol. 1997, 159, 970–975.
- Boes, M.; Prodeus, A.P.; Schmidt, T.; Carroll, M.C.; Chen, J. A critical role of natural immunoglobulin M in immediate defense against systemic bacterial infection. J. Exp. Med. 1998, 188, 2381–2386.
- Zhou, F.; Zhang, G.-X.; Rostami, A. LPS-treated bone marrow-derived dendritic cells induce immune tolerance through modulating differentiation of CD4+ regulatory T cell subpopulations mediated by 3G11 and CD127. Immunol. Res. 2016, 65, 630–638.
- Hayashi, T.; Gray, C.S.; Chan, M.; Tawatao, R.I.; Ronacher, L.; McGargill, M.A.; Datta, S.K.; Carson, D.A.; Corr, M. Prevention of autoimmune disease by induction of tolerance to Toll-like receptor 7. Proc. Natl. Acad. Sci. USA 2009, 106, 2764–2769.
- Mbongue, J.; Nicholas, D.; Firek, A.; Langridge, W. The role of dendritic cells in tissue-specific autoimmunity. J. Immunol. Res. 2014, 2014.
- Joffre, O.; Nolte, M.A.; Spörri, R.; Sousa, C.R.E. Inflammatory signals in dendritic cell activation and the induction of adaptive immunity. Immunol. Rev. 2009, 227, 234–247.
- Kumar, H.; Kawai, T.; Akira, S. Toll-like receptors and innate immunity. Biochem. Biophys. Res. Commun. 2009, 388, 621–625.
- Steinman, R.M.; Banchereau, J. Taking dendritic cells into medicine. Nature 2007, 449, 419–426.
- Galkina, E.; Ley, K. Immune and Inflammatory Mechanisms of Atherosclerosis. Annu. Rev. Immunol. 2009, 27, 165–197.
- Oyoshi, M.K.; He, R.; Kumar, L.; Yoon, J.; Geha, R.S. Cellular and molecular mechanisms in atopic dermatitis. Adv. Immunol. 2009, 102, 135–226.
- Said, E.A.; Dupuy, F.P.; Trautmann, L.; Zhang, Y.; Shi, Y.; El-Far, M.; Hill, B.J.; Noto, A.; Ancuta, P.; Peretz, Y.; et al. Programmed death-1-induced interleukin-10 production by monocytes impairs CD4+ T cell activation during HIV infection. Nat. Med. 2010, 16, 452–459.
- Agarwal, S.; Piesco, N.; Johns, L.; Riccelli, A. Differential Expression of IL-1β, TNF-α, IL-6, and IL-8 in Human Monocytes in Response to Lipopolysaccharides from Different Microbes. J. Dent. Res. 1995, 74, 1057–1065.
- Bryn, T.; Yaqub, S.; Mahic, M.; Henjum, K.; Aandahl, E.M.; Taskén, K. LPS-activated monocytes suppress T-cell immune responses and induce FOXP3+ T cells through a COX-2-PGE2-dependent mechanism. Int. Immunol. 2008, 20, 235–245.
- Adib-Conquy, M.; Adrie, C.; Moine, P.; Asehnoune, K.; Fitting, C.; Pinsky, M.R.; Dhainaut, J.F.; Cavaillon, J.M. NF-kappaB expression in mononuclear cells of patients with sepsis resembles that observed in lipopolysaccharide tolerance. Am. J. Respir. Crit. Care Med. 2000, 162, 1877–1883.
- López-Collazo, E.; del Fresno, C. Pathophysiology of endotoxin tolerance: Mechanisms and clinical consequences. Crit. Care 2013, 17, 242.
- Randolph, G.J.; Jakubzick, C.; Qu, C. Antigen presentation by monocytes and monocyte-derived cells. Curr. Opin. Immunol. 2008, 20, 52–60.
- Morin, J.; Faideau, B.; Gagnerault, M.; Lepault, F.; Boitard, C.; Boudaly, S. Passive transfer of flt-3L-derived dendritic cells delays diabetes development in NOD mice and associates with early production of interleukin (IL)-4 and IL-10 in the spleen of recipient mice. Clin. Exp. Immunol. 2003, 134, 388–395.
- Kim, N.-S.; Torrez, T.; Langridge, W. LPS enhances CTB-INSULIN induction of IDO1 and IL-10 synthesis in human dendritic cells. Cell. Immunol. 2019, 338, 32–42.
- Klaska, I.P.; Muckersie, E.; Martin-Granados, C.; Christofi, M.; Forrester, J.V. Lipopolysaccharide-primed heterotolerant dendritic cells suppress experimental autoimmune uveoretinitis by multiple mechanisms. Immunology 2016, 150, 364–377.
- Braun-Fahrländer, C.; Riedler, J.; Herz, U.; Eder, W.; Waser, M.; Grize, L.; Maisch, S.; Carr, D.; Gerlach, F.; Bufe, A.; et al. Environmental exposure to endotoxin and its relation to asthma in school-age children. N. Engl. J. Med. 2002, 347, 869–877.
- Bashir, M.E.H.; Louie, S.; Shi, H.N.; Nagler-Anderson, C. Toll-Like receptor 4 signaling by intestinal microbes influences susceptibility to food allergy. J. Immunol. 2004, 172, 6978–6987.
- Vatanen, T.; Kostic, A.D.; d’Hennezel, E.; Siljander, H.; Franzosa, E.A.; Yassour, M.; Kolde, R.; Vlamakis, H.; Arthur, T.D.; Hämäläinen, A.M.; et al. Variation in Microbiome LPS Immunogenicity Contributes to Autoimmunity in Humans. Cell 2016, 165, 1551.
- Mbongue, J.C.; Nicholas, D.A.; Zhang, K.; Kim, N.S.; Hamilton, B.N.; Larios, M.; Zhang, G.; Umezawa, K.; Firek, A.F.; Langridge, W.H. Induction of indoleamine 2, 3-dioxygenase in human dendritic cells by a cholera toxin B subunit-proinsulin vaccine. PLoS ONE 2015, 10, e0118562.
- Kim, N.-S.; Mbongue, J.C.; Nicholas, D.A.; Esebanmen, G.E.; Unternaehrer, J.J.; Firek, A.F.; Langridge, W.H.R. Chimeric Vaccine Stimulation of Human Dendritic Cell Indoleamine 2, 3-Dioxygenase Occurs via the Non-Canonical NF-κB Pathway. PLoS ONE 2016, 11, e0147509.
- Zariri, A.; van der Ley, P. Biosynthetically engineered lipopolysaccharide as vaccine adjuvant. Expert Rev. Vaccines 2015, 14, 861–876.
- Chilton, P.M.; Hadel, D.M.; To, T.T.; Mitchell, T.C.; Darveau, R.P. Adjuvant activity of naturally occurring monophosphoryl lipopolysaccharide preparations from mucosa-associated bacteria. Infect. Immun. 2013, 81, 3317–3325.
- Raetz, C.R.; Whitfield, C. Lipopolysaccharide endotoxins. Annu. Rev. Biochem. 2002, 71, 635–700.
- Gaspar, J.A.; Thomas, J.A.; Marolda, C.L.; Valvano, M.A. Surface expression of O-specific lipopolysaccharide in Escherichia coli requires the function of the TolA protein. Mol. Microbiol. 2000, 38, 262–275.
- Whitfield, C. Biosynthesis and Assembly of Capsular Polysaccharides in Escherichia coli. Annu. Rev. Biochem. 2006, 75, 39–68.
- Vinés, E.D.; Marolda, C.L.; Balachandran, A.; Valvano, M.A. Defective O-Antigen polymerization in tolA and pal mutants of Escherichia coli in response to Extracytoplasmic stress. J. Bacteriol. 2005, 187, 3359–3368.
- Hancock, R.E.; Diamond, G. The role of cationic antimicrobial peptides in innate host defences. Trends Microbiol. 2000, 8, 402–410.
- Papo, N.; Shai, Y. A molecular mechanism for lipopolysaccharide protection of gram-negative bacteria from antimicrobial peptides. J. Biol. Chem. 2005, 280, 10378–10387.
- Surapaneni, K.M.; Vishnu Priya, V.; Mallika, J. Effect of pioglitazone, quercetin, and hydroxy citric acid on vascular endothelial growth factor messenger RNA (VEGF mRNA) expression in experimentally induced nonalcoholic steatohepatitis (NASH). Turk. J. Med. Sci. 2015, 45, 542–546.
- Nitkin, C.R.; Xia, S.; Menden, H.; Yu, W.; Xiong, M.; Heruth, D.P.; Ye, S.Q.; Sampath, V. FOSL1 is a novel mediator of endotoxin/lipopolysaccharide-induced pulmonary angiogenic signaling. Sci. Rep. 2020, 10, 1–14.
- Mamat, U.; Wilke, K.; Bramhill, D.; Schromm, A.B.; Lindner, B.; Kohl, T.A.; Corchero, J.L.; Villaverde, A.; Schaffer, L.; Head, S.R.; et al. Detoxifying Escherichia coli for endotoxin-free production of recombinant proteins. Microb. Cell Factories 2015, 14, 57.
- Hoffmann, J.A. The immune response of Drosophila. Nature 2003, 426, 33–38.
- Royet, J. Drosophila melanogaster innate immunity: An emerging role for peptidoglycan recognition proteins in bacteria detection. Experientia 2004, 61, 537–546.
- Hultmark, D. Immune reactions in Drosophila and other insects: A model for innate immunity. Trends Genet. 1993, 9, 178–183.
- Lackie, A. Immune mechanisms in insects. Parasitol. Today 1988, 4, 98–105.
- Cociancich, S.; Bulet, P.; Hetru, C.; Hoffmann, J. The inducible antibacterial peptides of insects. Parasitol. Today 1994, 10, 132–139.
- Cociancich, S.; Dupont, A.; Hegy, G.; Lanot, R.; Holder, F.; Hetru, C.; Hoffmann, J.A.; Bulet, P. Novel inducible antibacterial peptides from a hemipteran insect, the sap-sucking bug Pyrrhocoris apterus. Biochem. J. 1994, 300, 567–575.
- Koizumi, N.; Imamura, M.; Kadotani, T.; Yaoi, K.; Iwahana, H.; Sato, R. The lipopolysaccharide-binding protein participating in hemocyte nodule formation in the silkworm Bombyx mori is a novel member of the C-type lectin superfamily with two different tandem carbohydrate-recognition domains. FEBS Lett. 1999, 443, 139–143.
- Wittwer, D.; Weise, C.; Götz, P.; Wiesner, A. LPS (Lipopolysaccharide)-activated immune responses in a hemocyte cell line from Estigmene acraea (Lepidoptera). Dev. Comp. Immunol. 1997, 21, 323–336.
- Kato, Y.; Motoi, Y.; Taniai, K.; Kadono-Okuda, K.; Yamamoto, M.; Higashino, Y.; Shimabukuro, M.; Chowdhury, S.; Xu, J.; Sugiyama, M.; et al. Lipopolysaccharide-lipophorin complex formation in insect hemolymph: A common pathway of lipopolysaccharide detoxification both in insects and in mammals. Insect Biochem. Mol. Biol. 1994, 24, 547–555.
- Kawabata, S.-I.; Nagayama, R.; Hirata, M.; Shigenaga, T.; Agarwala, K.L.; Saito, T.; Cho, J.; Nakajima, H.; Takagi, T.; Iwanaga, S. Tachycitin, a Small Granular Component in Horseshoe Crab Hemocytes, Is an Antimicrobial Protein with Chitin-Binding Activity. J. Biochem. 1996, 120, 1253–1260.
- Kawabata, S.-I.; Saeki, K.; Iwanaga, S. Limulus kexin: A new type of Kex2-like endoprotease specifically expressed in hemocytes of the horseshoe crab. FEBS Lett. 1996, 386, 201–204.
- Kawabata, S.-I.; Tokunaga, F.; Kugi, Y.; Motoyama, S.; Miura, Y.; Hirata, M.; Iwanaga, S. Limulus factor D, a 43-kDa protein isolated from horseshoe crab hemocytes, is a serine protease homologue with antimicrobial activity. FEBS Lett. 1996, 398, 146–150.
- Xu, W.-H.; Sato, Y.; Ikeda, M.; Yamashita, O. Molecular characterization of the gene encoding the precursor protein of diapause hormone and pheromone biosynthesis activating neuropeptide (DH-PBAN) of the silkworm, Bombyx mori and its distribution in some insects. Biochim. Biophys. Acta (BBA) Gene Struct. Expr. 1995, 1261, 83–89.
- Xu, W.-H.; Sato, Y.; Ikeda, M.; Yamashita, O. Stage-dependent and temperature-controlled expression of the gene encoding the precursor protein of diapause hormone and pheromone biosynthesis activating neuropeptide in the silkworm, bombyx mori. J. Biol. Chem. 1995, 270, 3804–3808.
- Sugiyama, M.; Kuniyoshi, H.; Kotani, E.; Taniai, K.; Kadono-Okuda, K.; Kato, Y.; Yamamoto, M.; Shimabukuro, M.; Chowdhury, S.; Xu, J.; et al. Characterization of a Bombyx mori cDNA encoding a novel member of the attacin family of insect antibacterial proteins. Insect Biochem. Mol. Biol. 1995, 25, 385–392.
- Takeda, K.; Akira, S. Toll-like receptors in innate immunity. Int. Immunol. 2005, 17, 1–14.
- Kang, D.; Liu, G.; Lundström, A.; Gelius, E.; Steiner, H. A peptidoglycan recognition protein in innate immunity conserved from insects to humans. Proc. Natl. Acad. Sci. USA 1998, 95, 10078–10082.
- Fabrick, J.; Baker, J.; Kanost, M. cDNA cloning, purification, properties, and function of a β-1,3-glucan recognition protein from a pyralid moth, Plodiainterpunctella. Insect Biochem. Mol. Biol. 2003, 33, 579–594.
- Ma, C.; Kanost, M. A β1,3-Glucan Recognition protein from an insect, manduca sexta, agglutinates microorganisms and activates the Phenoloxidase cascade. J. Biol. Chem. 2000, 275, 7505–7514.
- Ochiai, M.; Ashida, M. Purification of a beta-1,3-glucan recognition protein in the prophenoloxidase activating system from hemolymph of the silkworm, Bombyx mori. J. Biol. Chem. 1988, 263, 12056–12062.
- Ashida, M.; Ochiai, M.; Niki, T. Immunolocalization of prophenoloxidase among hemocytes of the silkworm, Bombyx mori. Tissue Cell 1988, 20, 599–610.
- Dimopoulos, G.; Richman, A.; Müller, H.M.; Kafatos, F.C. Molecular immune responses of the mosquito Anopheles gambiae to bacteria and malaria parasites. Proc. Natl. Acad. Sci. USA 1997, 94, 11508–11513.
- Dimopoulos, G.; Seeley, D.; Wolf, A.; Kafatos, F.C. Malaria infection of the mosquito Anopheles gambiae activates immune-responsive genes during critical transition stages of the parasite life cycle. EMBO J. 1998, 17, 6115–6123.
- Yeh, M.-S.; Lai, C.-Y.; Liu, C.-H.; Kuo, C.-M.; Cheng, W. A second proPO present in white shrimp Litopenaeus vannamei and expression of the proPOs during a Vibrio alginolyticus injection, molt stage, and oral sodium alginate ingestion. Fish Shellfish Immunol. 2009, 26, 49–55.
- Yeh, M.-S.; Liu, C.-H.; Hung, C.-W.; Cheng, W. cDNA cloning, identification, tissue localisation, and transcription profile of a transglutaminase from white shrimp, Litopenaeus vannamei, after infection by Vibrio alginolyticus. Fish Shellfish Immunol. 2009, 27, 748–756.
- Litman, G.W.; Rast, J.P.; Fugmann, S.D. The origins of vertebrate adaptive immunity. Nat. Rev. Immunol. 2010, 10, 543–553.
- Miller, J.S.; Nguyen, T.; Stanley-Samuelson, D.W. Eicosanoids mediate insect nodulation responses to bacterial infections. Proc. Natl. Acad. Sci. USA 1994, 91, 12418–12422.
- Jomori, T.; Natori, S. Function of the lipopolysaccharide-binding protein of Periplaneta americana as an opsonin. FEBS Lett 1992, 296, 283–286.
- Shigenaga, T.; Takayenoki, Y.; Kawasaki, S.; Seki, N.; Muta, T.; Toh, Y.; Ito, A.; Iwanaga, S. Separation of large and small granules from horseshoe crab (Tachypleus tridentatus) hemocytes and characterization of their components1. J. Biochem. 1993, 114, 307–316.
- Marmaras, V.J.; Charalambidis, N.D.; Zervas, C. Immune response in insects: The role of phenoloxidase in defense reactions in relation to melanization and sclerotization. Arch. Insect Biochem. Physiol. 1996, 31, 119–133.
More
Information
Subjects:
Immunology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.8K
Revisions:
2 times
(View History)
Update Date:
25 Aug 2022
Table of Contents



Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No