Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | NKULU KABANGE ROLLY | -- | 3208 | 2022-08-24 11:00:09 | | | |
| 2 | NKULU KABANGE ROLLY | Meta information modification | 3208 | 2022-08-24 15:24:47 | | | | |
| 3 | Jessie Wu | Meta information modification | 3208 | 2022-08-25 03:13:29 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Kabange, N.R.; Lee, S.; Shin, D.; Lee, J.; Kwon, Y.; Kang, J.; Cha, J.; Park, H.; Alibu, S.; Lee, J. Multiple Facets of Nitrogen. Encyclopedia. Available online: https://encyclopedia.pub/entry/26431 (accessed on 08 August 2026).
Kabange NR, Lee S, Shin D, Lee J, Kwon Y, Kang J, et al. Multiple Facets of Nitrogen. Encyclopedia. Available at: https://encyclopedia.pub/entry/26431. Accessed August 08, 2026.
Kabange, Nkulu Rolly, So-Myeong Lee, Dongjin Shin, Ji-Yoon Lee, Youngho Kwon, Ju-Won Kang, Jin-Kyung Cha, Hyeonjin Park, Simon Alibu, Jong-Hee Lee. "Multiple Facets of Nitrogen" Encyclopedia, https://encyclopedia.pub/entry/26431 (accessed August 08, 2026).
Kabange, N.R., Lee, S., Shin, D., Lee, J., Kwon, Y., Kang, J., Cha, J., Park, H., Alibu, S., & Lee, J. (2022, August 24). Multiple Facets of Nitrogen. In Encyclopedia. https://encyclopedia.pub/entry/26431
Kabange, Nkulu Rolly, et al. "Multiple Facets of Nitrogen." Encyclopedia. Web. 24 August, 2022.
Copy Citation
Nitrogen is a gas present in the air with an atomic mass of 14.007, a boiling and melting points of 77.36 K and 63.15 K, respectively, and a density of 0.0012506 g·cm–3. N was first discovered in 1772 by the Scottish physician and chemist Daniel Rutherford. The multiple facets of N are described. A paradigm shift is important to shape to the future use of N-rich fertilizers in crop production and their contribution to the current global greenhouse gases (GHGs) budget would help tackle current global environmental challenges toward a sustainable agriculture.
nitrogen
agriculture
environment
1. Basic Properties of Nitrogen and N-Containing Compounds
Nitrogen is the fifth most abundant element in the universe after hydrogen (H, first), helium (He, second), oxygen (O, third), and neon (Ne, fourth), and it makes up approximately 78.1% of the earth’s atmosphere. Reports indicate that an estimate of 4000 trillion tons of N can be found in the atmosphere in the form of N2 (https://pubchem.ncbi.gov/element/Nitrogen, accessed on 31 May 2022). The most popular use of N is for the production of ammonia (NH3) when combined with hydrogen (H), in a process called the “Haber process” [1]. Then, large amounts of NH3 are used to produce mineral fertilizers in a process known as the “Ostwald process”, among other uses [2]. N is found in all living systems as part of the makeup of biological compounds. During the decomposition of organic matter (OM), sodium nitrate (NaNO3) and potassium nitrate (KNO3) are formed. Other inorganic N compounds include HNO3, NH3, the oxides (nitric oxide (NO), nitrogen dioxide (NO2), N2O4, and nitrous oxide (N2O)), and cyanides (CN). NO2, NO, nitrous acid (HONO), and nitric acid (HNO3) belong to a group of highly reactive gases known as oxides of nitrogen or nitrogen oxides (NOx) [3].
NO2 is formed during nitrification and denitrification processes, in which N2O and NO are released, with N2O reported to be formed from NO3-dependent NO formation [4]. N2O is a potent greenhouse gas (GHG), with a global warming potential (GWP) 300 times greater than the mass of carbon dioxide (CO2) in the atmosphere, right before methane (CH4) and CO2 [5][6]. As for NO, reports indicate that this molecule is versatile and bioactive, with the potential to diffuse through biological membranes owing to its physiological properties. NO can act as a signaling molecule, which may involve a very wide web with reactive oxygen species (ROS). However, excessive accumulation of NO induces stressful conditions in plants [7][8]. NO and its derived molecules are reported to be involved in abiotic and biotic stress response mechanisms.
2. Historical Use of Nitrogen and N-Rich Fertilizers in Agriculture
The history of agriculture revealed that a number of plant species were domesticated from wild ancestors [9][10][11][12][13][14] during the Early Pre-Pottery Neolithic period, at various locations and different times between 10,500 and 10,100 years before common era, BCE [15][16][17]. During this period, crops exhibited a low productivity, poor yields, and poor quality, mainly attributed to their genetic makeup [16][18][19][20][21][22][23]. Since then, significant progress has been made using plant breeding and the establishment of plant nutrition schemes. According to Pennazio [24], the development of mineral nutrition of plants began between the 17th and 18th centuries. The patterns of mineral fertilizer applications in different cropping systems, as well as their impact on the environment, continue to nourish the debate globally [25][26][27][28][29][30][31][32][33][34][35][36][37][38]. However, the increase in food demands due to the rapid increase in the global population has shown the necessity to enhance the productivity of food crops. To achieve that, the common strategies used are crop improvement and the use of mineral fertilizers during crop cultivation, which have increased over the years [39][40]. Despite the recorded progress in plant breeding, N remains an indispensable macronutrient, among the 14 mineral elements (macro- and micronutrients) required by plants for optimum growth and development, high productivity, quality, fitness, and resistance toward environmental and biotic stresses [41][42].
Data indicate that the application of N-rich fertilizers increased over the years concomitant with the expansion of crop cultivation areas and the use of improved or high-yielding crop varieties that are demanding of nutrients, particularly observed in large-scale farming systems. This trend could be partially attributed to the increase in food demands subsequent to the increase in the world’s population. An estimate of the global population growth shows that the number of people on Earth increased by 219.1% in 72 years, from about 2,499,322,157 in 1950 to nearly 7,975,105,156 in 2022 (https://www.macrotrends.net/countries/WLD/world/population, accessed on 15 July 2022), in contrast to the decreasing pattern of the annual population growth rate during the same period (nearly 1.75% in 1951 down to 0.83% in 2022, a decrease by 47.4%). Available data on land coverage, as reported by the Food and Agriculture Organization of the United Nations (FAO) statistics (https://www.fao.org/faostat/en/#data/LC/visualize, accessed on 15 July 2022), suggest that herbaceous crops occupied about 1,877,418.8 ha of cultivated lands in 1992 compared to 1,904,136.4 ha in 2020 globally. In addition, the report on the use of nutrients for agricultural production during the last six decades indicates that nearly 4,155,951,874 metric tons of N was used between 1961 and 2020 (11,455,804.3 mt in 1961 and 113,291,696.7 mt in 2020) (https://www.fao.org/faostat/en/#compare, accessed on 15 July 2022). When compared with other macronutrients (phosphorus (P) and potassium (K)), N is by far the most abundantly used plant nutrient element in agriculture (nearly 4.15 billion tons (BT) within 59 years). For instance, a report by FAO showed that during 1961–2020, more than 1.9 BT of phosphate (P2O5) and 1.4 BT of potash (K2O) were used for agricultural production globally (Figure 1).
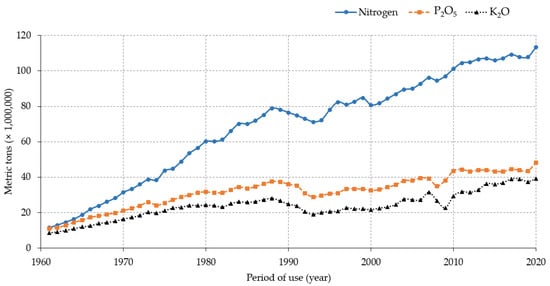
Figure 1. Pattern of global macronutrients use in agriculture from 1961 to 2020.
3. Essentiality of Nitrogen, Sources, and Availability
Arnon and Stout [43] proposed three criteria for the essentiality of a plant nutrient as follows: (1) a deficiency of the element makes it impossible for a plant to complete its lifecycle; (2) the deficiency is specific to the element in question; (3) the element is directly involved in the nutrition of the plant, such as a constituent of an essential metabolite or required for the action of an enzyme system. Other reports suggested that a more inclusive definition of the essentiality of a plant nutrient should be considered, which is not limited to those elements when they are deficient in the soil or unavailable, when they may cause symptoms of deficiency, and when their correction may involve an external supply through fertilization [44][45][46]. Therefore, on the basis of the abovementioned criteria, N and 15 other elements, namely, carbon (C), hydrogen (H), oxygen (O), phosphorus (P), potassium (K), calcium (Ca), magnesium (Mg), sulfur (S), iron (Fe), manganese (Mn), copper (Cu), zinc (Zn), molybdenum (Mo), boron (B), and chlorine (Cl), are considered essential for the growth of higher plants. Of this list, C, H, and O are acquired by the plant directly from the air and soil water, while the remaining 13 are supplied by the soil [47][48]. On the basis of the amounts in which the essential nutrients are acquired by the plant (except C, H, and O), they are categorized as primary (N, P, and K) and secondary macronutrients (Ca, Mg, and S), and micronutrients (Fe, Mn, Zn, Cu, B, Mo, and Cl).
Plants are sessile living organisms, which do not have the ability to move from one environment to another looking for food in the case of nutrient deficiency in their immediate environment [49][50]. Soil is the principal source of nutrients (including N) necessary for plants to complete their life cycle [51][52]. The availability of N is affected by various factors that may be extrinsic or intrinsic to the plant [53][54][55][56][57][58]. Crop productivity relies heavily on N fertilization [59][60][61]. N is found abundantly in the air but in a form that plants cannot absorb. The major sources of N are atmospheric nitrogen gas N2, exogenous N supply, and OM through fertilization. Atmospheric N2 gas (plentiful in the air but cannot be absorbed by plants in this form) is acquired through the nitrogen fixation cycle mediated by plant species belonging to Fabaceae (leguminous, along with a few non-leguminous plants containing nodules in their roots) in a symbiotic association with soil microorganisms known as nitrogen-fixing bacteria belonging to the genus Rhizobium. The latter mediate the conversion of atmospheric N2 to ammonium (NH4) in the soil, which in turn is converted to nitrate (NO3) during the nitrification process. According to Maier [62], the growth of many bacteria, either free-living in the environment or in symbiosis with plants, is promoted during N fixation in areas where fixed N is deficient in the soil. The root nodule bacteria have many O2-binding terminal oxidases, with a high O2 affinity, which are associated with N2 fixation and help maintain a steady O2 supply, coupled with ATP supply for high energy-demanding N2 fixation. The fertility of soil depends on several factors, including the quantity and quality of nutrients present in the soil, soil physical, biological, and chemical properties [63].
NO3 and NH4 are the major forms of N taken up by the plant, with NO3 being the most abundant [61]. However, roughly half of N (all N sources and forms considered) is used by the plant, while the remainder is either lost to groundwater (percolation or leaching), consumed by soil microorganisms (bacteria) involved in the decomposition of OM to humus, or converted back to atmospheric N2 via the denitrification process.
4. Nitrogen-Based Fertilizers
Nitrogen is commonly applied using commercial N-containing fertilizers or OM (composts, liquid organic fertilizers, and animal feces) [64][65][66][67][68]. One of the most abundant forms of N commercially available is urea 46% N ((CO(NH2)2) or CH4N2O) also known as carbamide, or it can be found together with P in the form of diammonium phosphate (DAP, (NH4)2HPO4). The latter is also referred to as ammonium monohydrogen phosphate, ammonium hydrogen phosphate, or ammonium phosphate dibasic, with the typical formulation of 18% N, 46% P2O5, 0% K2O) or triple superphosphate (TSP) referred to as calcium dihydrogen phosphate or monocalcium phosphate ((Ca(H2PO4)2·H2O), containing 45% phosphate (P2O5) (0–45–0), 15% Ca), whereas, K is supplied as potash (K2O) [58]. Together, they make up the trio widely known as NPK. In the soil, N is available to the plant and transported in the form of NO3 and NH4, with NO3 transport being predominant over NH4 and the major source N [69][70]. DAP is known as the world’s most widely used phosphorus fertilizer and one of the known water-soluble ammonium phosphate salts that can be produced when ammonia (NH3) reacts with phosphoric acid (H3PO4) ((NH4)2HPO4 (s) ⇌ NH3 (g) + (NH4)H2PO4 (s)) [71]. Reports indicate that, when applied as a source of N and P, DAP temporarily increases the soil pH but becomes more acidic over the long term upon nitrification of the NH4, and it is said to be incompatible with alkaline chemicals due to the high potential for the NH4 ion to convert to NH3 in a high-pH environment (pH 7.5–8.0).
5. Tracking Nitrogen Uptake and Assimilation in Plants
Fertilizers labeled with radioactive isotopes, such as phosphorus and nitrogen-15 have been used to investigate fertilizer uptake, retention, and utilization [72][73][74]. The N-15 (15N) isotopic technique, which may help identify the source of N2O generation and reduce the emission of this potent GHG during nitrification and denitrification processes, offers comparative advantages over conventional techniques for measuring the impact of climate change [74]. Stable isotope enrichment approaches have long been established to trace the source of N2O following the application of 15N-labeled fertilizers, such as 15N-labeled NH4 and NO3 [75]. This technique enables the determination of the source of fertilizer-derived 15N-N2O.
In general, nitrification derived N2O is quantified upon the supply of 15NH4, while that mediated by denitrification is measured following the supply of 15NO3. In the same way, considering that multiple pathways mediating N2O formation and consumption occur simultaneously in various microenvironments in soils, nitrification inhibitors and isotope signature techniques are commonly utilized to separate N2O-producing and -reducing pathways [76]. Stevens and Laughlin [77] suggested that the reduction of N2O to N2 could be quantified by determining 15N in N2 following the supply of 15NO3. Similarly, Baggs et al. [78] demonstrated that application of 14NH4/15NO3 and 15NH4/14NO3 helped determine the relative contributions of nitrification and denitrification to N2O production. Furthermore, He et al. [79] proposed the use of the stable 15N isotope as a means for quantifying N transfer between mycorrhizal plants, considering that plants acquire nutrients from soil, and mycorrhizae play vital roles in plant nutrient acquisition, performance, and productivity. In the same way, the use of the carbon-13 (13C) stable isotopic technique helps evaluate the source of carbon sequestered in the soil.
Isotopic fractionation [80][81][82][83] can cause the isotope amount ratio n(15N)/n(14N) to increase systematically through food chains via assimilation of N compounds in biomolecules such as proteins. Isotopic fractionation occurs because of assimilation, storage, and excretion of proteins and other N compounds. Isotope amount ratio n(15N)/n(14N) measurements have been widely used to test hypotheses about predator–prey relations and detect disruptions to the trophic structure of ecosystems that might be caused by toxic contaminants, invasive species, harvesting, or organisms. Similar principles are used to detect differences in diets among animals, including humans [84][85][86].
Artificially enriched 15N tracers are used to study movement and transformation of N in biological and environmental systems, such as the uptake and loss of N fertilizers by crops. A common experiment involves introducing an isotopically labeled compound into the environment and then analyzing various samples taken from the environment for the presence of the enriched isotope to determine where the labeled compound moved and whether it transformed into another compound. Artificially enriched 15N has also been employed to study uptake and dispersal of N in feed supplies used in food production industries such as aquaculture [87].
The stable isotopes of N are subject to isotopic fractionation via physical, chemical, and biological processes. Variations in the isotope amount ratio n(15N)/n(14N) are commonly used to study Earth system processes, especially those related to biology, because N is a major nutrient for growth [88]. To illustrate this, isotope fractionation occurs when dissolved solutes, such as nitrate (NO3), are transformed to more reduced compounds (i.e., N2) because NO3 with higher 14N abundance tends to be more readily broken down. This leaves the residual unreacted NO3 with a higher n(15N)/n(14N) ratio than the initial ratio prior to reaction. Changes in the isotopic composition of biologically reactive compounds can be used to detect such reactions in aquatic environments, which are important mechanisms for removing reactive contaminants such as NO3 [89].
Variation in N stable isotopes has been used to track fertilizer N accumulation into plants, soils, and infiltrating groundwater to improve efficiency and reduce impact on the environment, such as experimental agricultural fields where various amounts of excess N from fertilizers and plant residues can be found in groundwater.
6. Paradigm Shift to a Sustainable Agricultural Production System
Agriculture is a major source of food. Food production is required to double within the next 8–10 decades or so from the perspective of the rapid population growth. Agriculture has been identified as a sink and source of GHGs, as well as the economic sector that suffers the most from climate change. As a source, reports support that the application of N-rich fertilizers is regarded as one of the factors contributing significantly to enhancing GHGs formation during crops cultivation. Rodale [90] proposed paths to transition to a sustainable agricultural production system on the basis of the following observations: (i) because renewable resources are the basis of operation and productivity of modern agriculture, in the event of depletion of nonrenewable resources, either food will become extremely expensive or productivity will decline; (ii) the current food production system contributes to the environmental degradation (soil erosion, soil degradation, and deforestation); (iii) lines of evidence indicate that a number of agricultural practices, including the pattern of N-rich fertilizer application, contribute to the escalation of pollution problems; (iv) there is a strong agreement that the natural resources are limited and should be used in a sustainable manner; (v) conventional technologies and secular agricultural production systems are likely to be unsustainable in the future in the event that agricultural production becomes the major source of energy and feed stocks; (vi) a major concern exists over whether the good life in rural areas can be maintained if family farms are replaced by large-scale industrialized farms, which produce all the food.
Owing to the above, sustainable agriculture, as proposed by diverse sources and condensed by the International Food and Agriculture Development (IFAD) task force report [91], could be portrayed as follows: (i) the successful management of resources to satisfy changing human needs, while maintaining or enhancing the natural resources base and avoiding environmental degradation; (ii) the ability of an agricultural system to maintain production over time in the face of social and economic pressures; (iii) one that should conserve and protect natural resources and allow for long-term economic growth by managing all exploited resources for sustainable yields. Other sources argue that stainability can only be achieved when resources, inputs, and technologies are within the capabilities of the farmers to own, hire, and manage with increasing efficiency to achieve desirable levels of productivity with minimal effects on the resources base, human life, and environmental quality. In this regard, sustainable agricultural production system is referred to as “one that maintains an acceptable and increasing level of productivity that satisfies prevailing needs and is continuously adapted to meet the future needs for increasing the carrying of the resource base and other worthwhile human needs” [92][93][94][95].
Nevertheless, the major challenge remains the reduction in GHG emissions from agriculture during crop cultivation. The last two decades have been marked by a hunt for sustainable and effective strategies toward achieving a sustainable agriculture, here referring to agricultural practices that help reduce, in a sustainable manner, the emission of GHGs. Among them, improving N use efficiency (NUE, from uptake to assimilation and remobilization) is considered the most promising and effective approach, because it allows significantly reducing the application of N-rich fertilizers, which helps cut down the production cost, with a low impact on the environment. Reports have demonstrated that NUE is controlled genetically, and this complex trait allows optimizing the use of N available to the plant. NUE is equally important when plants experience nutrients shortage or unavailability caused by environmental stresses, such as drought [96][97][98], salinity [99], heat stress [100], or heavy-metal toxicity, whereby the acquisition of N is either restricted or impaired. In addition to the well-characterized NO3 or NH4 transport- and assimilation-related genes [101], N remobilization (one of the components of NUE) from NO3 stored in the vacuole regulated through a process known as autophagy carries the potential to salvage poor N supply or transport within the plant, and it offers an alternative for balanced plant productivity and reduced GHG emissions [100][102][103][104]. Likewise, NO, an ancient molecule with multiple roles in the plant, previously suggested to play an important role in N acquisition and assimilation events through NR activity [105][106], could serve as a potential target for improving NUE in plants.
Other alternative approaches may include intermittent drainage, especially in flooded or irrigated cultivation systems, the use of nitrification inhibitors, and the application of biochar. Unlike in the industrial sector, it is important to indicate that net zero GHG emission may not be a realistic target from agriculture with regard to the biological nature of CH4 and N2O production, involving soil microorganisms such as methanotrophs and methanogens, and the requirement of CO2 in plant metabolic processes.
References
- Clark, J.J. The Haber Process for the Manufacture of Ammonia. 2017. Available online: https://www.chemguide.co.uk (accessed on 10 July 2022).
- Wang, Y.; Yu, Y.; Jia, R.; Zhang, C.; Zhang, B. Electrochemical synthesis of nitric acid from air and ammonia through waste utilization. Nat. Sci. Rev. 2019, 6, 730–738.
- World Health Organization. Who Guidelines for Indoor Air Quality: Selected Pollutants; World Health Organization, Regional Office for Europe: Geneva, Switzerland, 2010.
- Timilsina, A.; Zhang, C.; Pandey, B.; Bizimana, F.; Dong, W.; Hu, C. Potential pathway of nitrous oxide formation in plants. Front. Plant Sci. 2020, 11, 1177.
- Davidson, E.A. The contribution of manure and fertilizer nitrogen to atmospheric nitrous oxide since 1860. Nat. Geosci. 2009, 2, 659–662.
- Syakila, A.; Kroeze, C. The global nitrous oxide budget revisited. Greenh. Gas Meas. Manag. 2011, 1, 17–26.
- Corpas, F.J.; Barroso, J.B. Nitro-oxidative stress vs oxidative or nitrosative stress in higher plants. New Phytol. 2013, 199, 633–635.
- Oz, M.T.; Eyidogan, F.; Yucel, M.; Öktem, H.A. Functional role of nitric oxide under abiotic stress conditions. In Nitric Oxide Action in Abiotic Stress Responses in Plants; Springer: Basel, Switzerland, 2015; pp. 21–41.
- Purugganan, M.D. Evolutionary insights into the nature of plant domestication. Curr. Biol. 2019, 29, R705–R714.
- Sang, T.; Ge, S. Genetics and phylogenetics of rice domestication. Curr. Opin. Genet. Dev. 2007, 17, 533–538.
- Heslop-Harrison, J.S.; Schwarzacher, T. Domestication, genomics and the future for banana. Ann. Bot. 2007, 100, 1073–1084.
- Schaal, B. Plants and people: Our shared history and future. New Phytol. 2019, 1, 14–19.
- Larson, G.; Piperno, D.R.; Allaby, R.G.; Purugganan, M.D.; Andersson, L.; Arroyo-Kalin, M.; Barton, L.; Vigueira, C.C.; Denham, T.; Dobney, K. Current perspectives and the future of domestication studies. Proc. Nat. Acad. Sci. USA 2014, 111, 6139–6146.
- Zeder, M.A.; Bradley, D.G.; Smith, B.D.; Emshwiller, E. Documenting Domestication: New Genetic and Archaeological Paradigms; University of California Press: Berkeley, CA, USA, 2006.
- Zohary, D.; Hopf, M.; Weiss, E. Domestication of Plants in the Old World: The Origin and Spread of Domesticated Plants in Southwest Asia, Europe, and the Mediterranean Basin; Oxford University Press: London, UK, 2012.
- Hancock, J.F. Plant Evolution and the Origin of Crop Species; CABI: Wallingford, UK, 2012.
- Stetter, M.G.; Gates, D.J.; Mei, W.; Ross-Ibarra, J. How to make a domesticate. Curr. Biol. 2017, 27, R896–R900.
- Doebley, J.; Stec, A.; Wendel, J.; Edwards, M. Genetic and morphological analysis of a maize-teosinte F2 population: Implications for the origin of maize. Proc. Nat. Acad. Sci. USA 1990, 87, 9888–9892.
- Harlan, J.R.; Zohary, D. Distribution of wild wheats and barley: The present distribution of wild forms may provide clues to the regions of early cereal domestication. Science 1966, 153, 1074–1080.
- Baker, H.G. Plants and Civilization; Macmillan International Higher Education: London, UK, 1970.
- Zohary, M. Flora Palaestina: Platanaceae to Umbelliferae, 2nd ed.; Israel Academy of Sciences and Humanities: Jerusalem, Israel, 1972.
- Langer, R.H.M.; Langer, R.; Hill, G. Agricultural Plants; Cambridge University Press: Cambridge, UK, 1991.
- Martin, G.B.; Adams, M.W. Landraces of Phaseolus vulgaris (fabaceae) in northern malawi. I. Regional variation. Econ. Bot. 1987, 41, 190–203.
- Pennazio, S. Mineral nutrition of plants: A short history of plant physiology. Biol. Forum/Riv. Biol. 2005, 98, 215–236.
- Garcia-Oliveira, A.L.; Chander, S.; Ortiz, R.; Menkir, A.; Gedil, M. Genetic basis and breeding perspectives of grain iron and zinc enrichment in cereals. Front. Plant Sci. 2018, 9, 937.
- Abdullah, M.; Hossain, K.L. Effects of urea-n fertilizer dosage supplemented with ipil-ipil tree litter on yield of rice and insect prevalence. J. For. Res. 2006, 17, 335–338.
- Paungfoo-Lonhienne, C.; Yeoh, Y.K.; Kasinadhuni, N.R.P.; Lonhienne, T.G.; Robinson, N.; Hugenholtz, P.; Ragan, M.A.; Schmidt, S. Nitrogen fertilizer dose alters fungal communities in sugarcane soil and rhizosphere. Sci. Rep. 2015, 5, 8678.
- Sari, Y.; Alkaff, M. Classification of rice leaf using fuzzy logic and hue saturation value (hsv) to determine fertilizer dosage. In Proceedings of the 2020 Fifth International Conference on Informatics and Computing (ICIC), Gorontalo, Indonesia, 3–4 November 2020; pp. 1–6.
- Rasaq, S.; Shittu, T.; Fadimu, G.; Abass, A.; Omoniyi, O. Effect of cassava variety, fertilizer type and dosage on the physicochemical, functional and pasting properties of high quality cassava flour (hqcf). Qual. Assur. Saf. Crop. Food. 2020, 12, 18–27.
- Ayoub, A.T. Fertilizers and the environment. Nutr. Cycl. Agroecosyst. 1999, 55, 117–121.
- Kim, S.; Dale, B.E. Effects of nitrogen fertilizer application on greenhouse gas emissions and economics of corn production. Environ. Sci. Technol. 2008, 42, 6028–6033.
- Xie, Z.; Xu, Y.; Liu, G.; Liu, Q.; Zhu, J.; Tu, C.; Amonette, J.E.; Cadisch, G.; Yong, J.W.; Hu, S.; et al. Impact of biochar application on nitrogen nutrition of rice, greenhouse-gas emissions and soil organic carbon dynamics in two paddy soils of china. Plant Soil 2013, 370, 527–540.
- Zhong, Y.; Wang, X.; Yang, J.; Zhao, X.; Ye, X. Exploring a suitable nitrogen fertilizer rate to reduce greenhouse gas emissions and ensure rice yields in paddy fields. Sci. Total Environ. 2016, 565, 420–426.
- Paterson, A.H. What has QTL mapping taught us about plant domestication? New Phytol. 2002, 154, 591–608.
- Purugganan, M.D.; Fuller, D.Q.J.N. The nature of selection during plant domestication. Nature 2009, 457, 843–848.
- Chaudhary, B. Plant domestication and resistance to herbivory. Int. J. Plant Genom. 2013, 2013, 1–14.
- Vaughan, D.A.; Lu, B.-R.; Tomooka, N. The evolving story of rice evolution. Plant Sci. 2008, 174, 394–408.
- Tan, L.; Li, X.; Liu, F.; Sun, X.; Li, C.; Zhu, Z.; Fu, Y.; Cai, H.; Wang, X.; Xie, D. Control of a key transition from prostrate to erect growth in rice domestication. Nat. Genet. 2008, 40, 1360–1364.
- van Dijk, M.; Morley, T.; Rau, M.L.; Saghai, Y. A meta-analysis of projected global food demand and population at risk of hunger for the period 2010–2050. Nat. Food 2021, 2, 494–501.
- Sadik, N. Population growth and the food crisis. Food Nutr. Agric. 1991, 1, 3–6.
- Six, J. Plant nutrition for sustainable development and global health. Plant Soil 2011, 339, 1–2.
- White, P.J.; Brown, P. Plant nutrition for sustainable development and global health. Ann. Biol. 2010, 105, 1073–1080.
- Arnon, D.I.; Stout, P. The essentiality of certain elements in minute quantity for plants with special reference to copper. Plant Physiol. 1939, 14, 371.
- Mengel, K.; Kirkby, E. Principles of Plant Nutrition; International Potash Institute: Worblaufen-Bern, Switzerland, 1987; pp. 687–695.
- Tisdale, S.L.; Nelson, W.L.; Beaton, J.D. Soil Fertility and Fertilizers; Collier Macmillan Publishers: Springfield, OH, USA, 1985.
- Nicholas, D. Minor mineral nutrients. Ann. Rev. 1961, 12, 63–90.
- Epstein, E. Mineral Nutrition of Plants: Principles and Perspectives; John Wiley Sons, Inc.: New York, NY, USA, 1972.
- Grundon, N.J.; Edwards, D.; Takkar, P.; Asher, C.J.; Clark, R. Nutritional Disorders of Grain Sorghum; Australian Center for International Agricultural Research (ACIAR): Canberra, Australia, 1987.
- Atkinson, N.J.; Urwin, P.E. The interaction of plant biotic and abiotic stresses: From genes to the field. J. Exp. Bot. 2012, 63, 3523–3543.
- Verma, S.; Nizam, S.; Verma, P.K. Biotic and abiotic stress signaling in plants. In Stress Signaling in Plants: Genomics and Proteomics Perspective, Volume 1; Springer: New York, NY, USA, 2013; pp. 25–49.
- Fageria, N.; Filho, M.B.; Moreira, A.; Guimarães, C.J. Foliar fertilization of crop plants. J. Plant Nutr. 2009, 32, 1044–1064.
- Lambers, H.; Raven, J.A.; Shaver, G.R.; Smith, S.E. Plant nutrient-acquisition strategies change with soil age. Trend. Ecol. Evol. 2008, 23, 95–103.
- Miransari, M. Soil microbes and the availability of soil nutrients. Acta Physiol. Plant. 2013, 35, 3075–3084.
- Binkley, D.; Vitousek, P. Soil nutrient availability. In Plant Physiological Ecology; Springer: Dordrecht, The Netherlands, 1989; pp. 75–96.
- Nemeth, K. The availability of nutrients in the soil as determined by electro-ultrafiltration (EUF). Adv. Agron. 1980, 31, 155–188.
- Nair, K.P. The buffering power of plant nutrients and effects on availability. Adv. Agron. 1996, 57, 237–287.
- Greger, M.; Landberg, T.; Vaculík, M. Silicon influences soil availability and accumulation of mineral nutrients in various plant species. Plants 2018, 7, 41.
- López-Arredondo, D.L.; Sánchez-Calderón, L.; Yong-Villalobos, L. Molecular and genetic basis of plant macronutrient use efficiency: Concepts, opportunities, and challenges. In Plant Macronutrient Use Efficiency; Elsevier: Amsterdam, The Netherlands, 2017; pp. 1–29.
- Xu, G.; Fan, X.; Miller, A.J. Plant nitrogen assimilation and use efficiency. Ann. Rev. Plant Biol. 2012, 63, 153–182.
- Masclaux-Daubresse, C.; Daniel-Vedele, F.; Dechorgnat, J.; Chardon, F.; Gaufichon, L.; Suzuki, A. Nitrogen uptake, assimilation and remobilization in plants: Challenges for sustainable and productive agriculture. Ann. Bot. 2010, 105, 1141–1157.
- Krapp, A. Plant nitrogen assimilation and its regulation: A complex puzzle with missing pieces. Curr. Opin. Plant Biol. 2015, 25, 115–122.
- Maier, R.J. Nitrogen fixation and respiration: Two processes linked by the energetic demands of nitrogenase. In Respiration in Archaea and Bacteria; Springer: Dordrecht, The Netherlands, 2004; pp. 101–120.
- Ellis, B.; Foth, H. Soil Fertility; CRC Press: Boca Raton, FL, USA, 1996.
- Burns, R.C.; Hardy, R.W. Nitrogen Fixation in Bacteria and Higher Plants; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012.
- Postgate, J. Nitrogen Fixation; Cambridge University Press: Cambridge, UK, 1998.
- Boddey, R.; Urquiaga, S.; Reis, V.; Döbereiner, J. Biological nitrogen fixation associated with sugar cane. In Nitrogen Fixation; Springer: Dordrecht, The Netherlands, 1991; pp. 105–111.
- Schubert, K.R. Products of biological nitrogen fixation in higher plants: Synthesis, transport, and metabolism. Ann. Rev. Plant Physiol. 1986, 37, 539–574.
- Bothe, H.; Ferguson, S.; Newton, W.E. Biology of the Nitrogen Cycle; Elsevier: London, UK, 2006.
- Mustapha, S.; Voncir, N.; Umar, S. Content and distribution of nitrogen forms in some black cotton soils in akko lga, gombe state, nigeria. Int. J. Soil Sci. 2011, 6, 275.
- Harmsen, G.; Van Schreven, D. Mineralization of organic nitrogen in soil. Adv. Agron. 1955, 7, 299–398.
- Lewu, F.; Volova, T.; Thomas, S.; Rakhimol, K. Controlled Release Fertilizers for Sustainable Agriculture; Academic Press: Cambridge, MA, USA, 2020.
- Singh, B.; Singh, J.; Kaur, A. Applications of radioisotopes in agriculture. Int. J. Biotchnol. Bioeng. 2013, 4, 167–174.
- Gaspar, M. Stable nitrogen isotope helps scientists optimize water, fertilizer use. IAEA Bull. 2017, 58, 18–19.
- Liu, R.; Hu, H.; Suter, H.; Hayden, H.L.; He, J.; Mele, P.; Chen, D. Nitrification is a primary driver of nitrous oxide production in laboratory microcosms from different land-use soils. Front. Microbiol. 2016, 7, 1373.
- Baggs, E. A review of stable isotope techniques for N2O source partitioning in soils: Recent progress, remaining challenges and future considerations. Rap. Comm. Mass Spectrom. 2008, 22, 1664–1672.
- Zhang, J.; Cai, Z.; Cheng, Y.; Zhu, T. Denitrification and total nitrogen gas production from forest soils of eastern China. Soil Biol. Biochem. 2009, 41, 2551–2557.
- Stevens, R.; Laughlin, R. Measurement of nitrous oxide and di-nitrogen emissions from agricultural soils. Nutr. Cycl. Agroecosyst. 1998, 52, 131–139.
- Baggs, E.; Blum, H. CH4 oxidation and emissions of CH4 and N2O from lolium perenne swards under elevated atmospheric CO2. Soil Biol. Biochem. 2004, 36, 713–723.
- He, X.; Xu, M.; Qiu, G.Y.; Zhou, J. Use of 15N stable isotope to quantify nitrogen transfer between mycorrhizal plants. J. Plant Ecol. 2009, 2, 107–118.
- Kotajima, S.; Koba, K.; Ikeda, D.; Terada, A.; Isaka, K.; Nishina, K.; Kimura, Y.; Makabe, A.; Yano, M.; Fujitani, H.; et al. Nitrogen and oxygen isotope signatures of nitrogen compounds during anammox in the laboratory and a wastewater treatment plant. Microb. Environ. 2020, 35, ME20031.
- Sánchez-Carrillo, S.; Álvarez-Cobelas, M. Stable isotopes as tracers in aquatic ecosystems. Environ. Rev. 2018, 26, 69–81.
- Trimmer, M.; Grey, J.; Heppell, C.M.; Hildrew, A.G.; Lansdown, K.; Stahl, H.; Yvon-Durocher, G. River bed carbon and nitrogen cycling: State of play and some new directions. Sci. Total Environ. 2012, 434, 143–158.
- Dabundo, R. Nitrogen Isotopes in the Measurement of N2-Fixation and the Estimation of Denitrification in the Global Ocean; UCONN Library: Storrs, CT, USA, 2014; Available online: https://opencommons.uconn.edu/ (accessed on 10 July 2022).
- Hopkins, J.B., III; Ferguson, J.M. Estimating the diets of animals using stable isotopes and a comprehensive Bayesian mixing model. PLoS ONE 2012, 7, e28478.
- Montoya, J.P. Nitrogen isotope fractionation in the modern ocean: Implications for the sedimentary record. In Carbon Cycling in the Glacial Ocean: Constraints on the Ocean’s Role in Global Change; Springer: Berlin/Heidelberg, Germany, 1994; pp. 259–279.
- Hedges, R.E.; Reynard, L.M. Nitrogen isotopes and the trophic level of humans in archaeology. J. Archeol. Sci. 2007, 34, 1240–1251.
- Burford, M.; Preston, N.; Glibert, P.; Dennison, W. Tracing the fate of 15n-enriched feed in an intensive shrimp system. Aquaculture 2002, 206, 199–216.
- Michener, R.; Lajtha, K. Stable Isotopes in Ecology and Environmental Science; John Wiley & Sons: Hoboken, NJ, USA, 2008.
- Granger, J.; Sigman, D.M.; Lehmann, M.F.; Tortell, P.D. Nitrogen and oxygen isotope fractionation during dissimilatory nitrate reduction by denitrifying bacteria. Limnol. Ocean. 2008, 53, 2533–2545.
- Rodale, R. Agricultural Systems: The Importance of Sustainability, National Forum; Honor Society of Phi Kappa Phi: Baton Rouge, LA, USA, 1988; p. 2.
- BIFAD. Environment and Natural Resources: Strategies for Sustainable Agriculture Task Force Report; US Agency for International Development: Washington, DC, USA, 1988.
- Okigbo, B. Development of Sustainable Agricultural Production Systems in Africa: Roles of International Agricultural Research Centers and National Agricultural Research Systems; Institute of Tropical Agriculture: Ibadan, Nigeria, 1991; Available online: https://agris.fao.org/agris-search/index.do (accessed on 24 June 2022).
- Dover, M.J.; Talbot, L.M. To Feed the Earth: Agro-Ecology for Sustainable Development; World Resources Institute: Washington, DC, USA, 1987.
- Knezek, B.D.; Hesterman, O.B.; Wink, L. Exploring a New Vision of Agriculture, National Forum; Honor Society of Phi Kappa Phi: Baton Rouge, LA, USA, 1988; p. 10.
- Lynam, J.K.; Herdt, R.W. Sense and sustainability: Sustainability as an objective in international agricultural research. Agric. Econom. 1989, 3, 381–398.
- Ding, L.; Lu, Z.; Gao, L.; Guo, S.; Shen, Q. Is nitrogen a key determinant of water transport and photosynthesis in higher plants upon drought stress? Front. Plant Sci. 2018, 9, 1143.
- Ramamoorthy, P.; Bheemanahalli, R.; Meyers, S.L.; Shankle, M.W.; Reddy, K.R. Drought, low nitrogen stress, and ultraviolet-b radiation effects on growth, development, and physiology of sweetpotato cultivars during early season. Genes 2022, 13, 156.
- Araus, V.; Swift, J.; Alvarez, J.M.; Henry, A.; Coruzzi, G.M. A balancing act: How plants integrate nitrogen and water signals. J. Exp. Bot. 2020, 71, 4442–4451.
- Tian, J.; Pang, Y.; Zhao, Z. Drought, salinity, and low nitrogen differentially affect the growth and nitrogen metabolism of sophora japonica (L.) in a semi-hydroponic phenotyping platform. Front. Plant Sci. 2021, 12, 715456.
- Zhen, F.; Liu, Y.; Ali, I.; Liu, B.; Liu, L.; Cao, W.; Tang, L.; Zhu, Y. Short-term heat stress at booting stage inhibited nitrogen remobilization to grain in rice. J. Agric. Food Res. 2020, 2, 100066.
- Ye, J.Y.; Tian, W.H.; Jin, C.W. Nitrogen in plants: From nutrition to the modulation of abiotic stress adaptation. Stress Biol. 2022, 2, 1–14.
- Fan, T.; Yang, W.; Zeng, X.; Xu, X.; Xu, Y.; Fan, X.; Luo, M.; Tian, C.; Xia, K.; Zhang, M. A rice autophagy gene OsATG8B is involved in nitrogen remobilization and control of grain quality. Front. Plant Sci. 2020, 11, 588.
- Zhen, X.; Zheng, N.; Yu, J.; Bi, C.; Xu, F. Autophagy mediates grain yield and nitrogen stress resistance by modulating nitrogen remobilization in rice. PLoS ONE 2021, 16, e0244996.
- Yu, J.; Zhen, X.; Li, X.; Li, N.; Xu, F. Increased autophagy of rice can increase yield and nitrogen use efficiency (NUE). Front. Plant Sci. 2019, 10, 584.
- Berger, A.; Boscari, A.; Horta Araujo, N.; Maucourt, M.; Hanchi, M.; Bernillon, S.; Rolin, D.; Puppo, A.; Brouquisse, R. Plant nitrate reductases regulate nitric oxide production and nitrogen-fixing metabolism during the medicago truncatula–sinorhizobium meliloti symbiosis. Front. Plant Sci. 2020, 11, 1313.
- Pan, Q.-N.; Geng, C.-C.; Li, D.-D.; Xu, S.-W.; Mao, D.-D.; Umbreen, S.; Loake, G.J.; Cui, B.-M. Nitrate reductase-mediated nitric oxide regulates the leaf shape in Arabidopsis by mediating the homeostasis of reactive oxygen species. Int. J. Mol. Sci. 2019, 20, 2235.
More
Information
Subjects:
Agronomy
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.1K
Entry Collection:
Environmental Sciences
Revisions:
3 times
(View History)
Update Date:
25 Aug 2022
Table of Contents



Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No