 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Xinghua Qin | -- | 3650 | 2022-08-14 10:03:10 | | | |
| 2 | Catherine Yang | + 2 word(s) | 3652 | 2022-08-15 03:25:44 | | | | |
| 3 | Catherine Yang | Meta information modification | 3652 | 2022-08-15 08:28:40 | | |
Video Upload Options
Atrial fibrillation (AF), the most commonly encountered arrhythmia in clinical practice, is characterized by irregular contractions of atrial cardiomyocytes. AF causes substantial disability and morbidity with a high risk of heart failure (HF) and ischemic stroke, and has exerted a tremendous burden on society, the health care system, and the economy. Metabolic flexibility is a novel concept that aptly describes switches in substrate metabolism depending on availability and requirements, thus coping with the dramatic fluctuations in energy supply and demand under physiological and pathological stimuli. Metabolic flexibility is critical for normal heart function, as it provides sufficient energy when the rapid and irregular contraction of atrial cardiomyocytes occurs during AF.
1. Metabolic Flexibility in the Normal Heart
1.1. Substrate Availability
1.2. Metabolic Regulatory Network
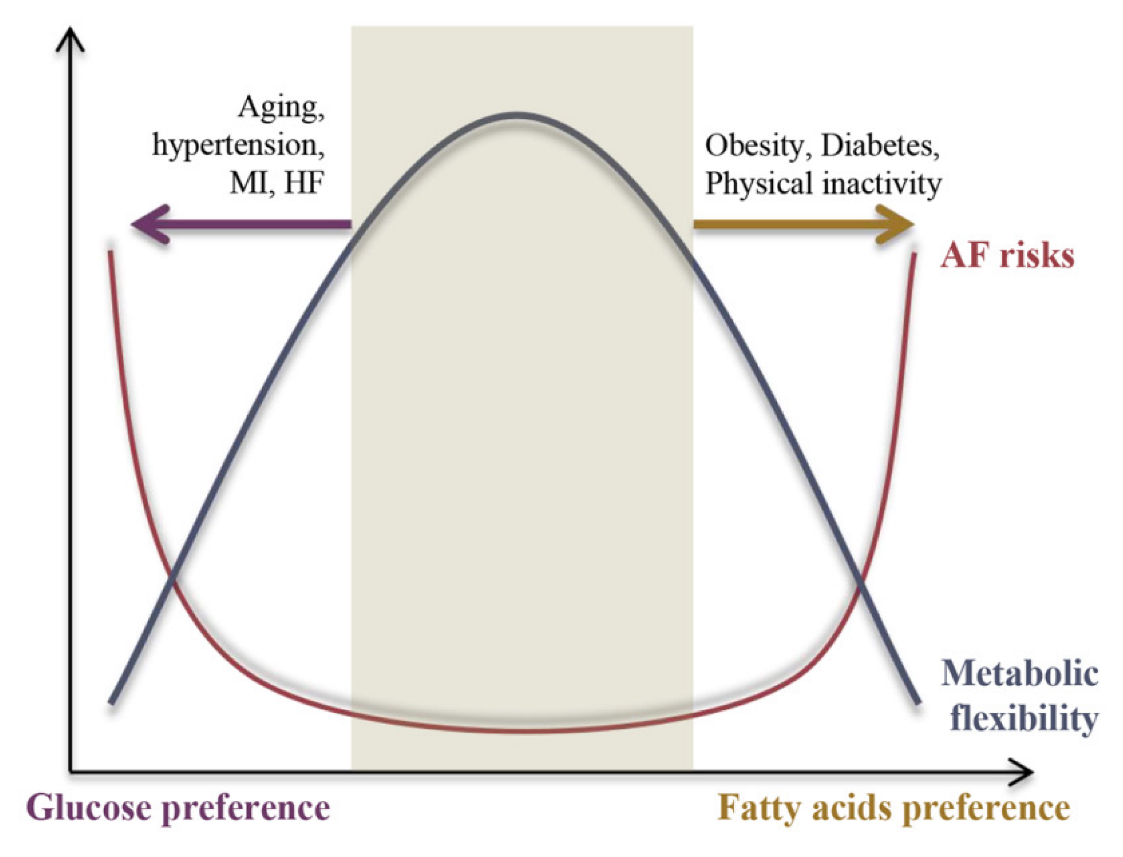
2. Metabolic Inflexibility as the Basis of Pathogenesis among AF Stressors
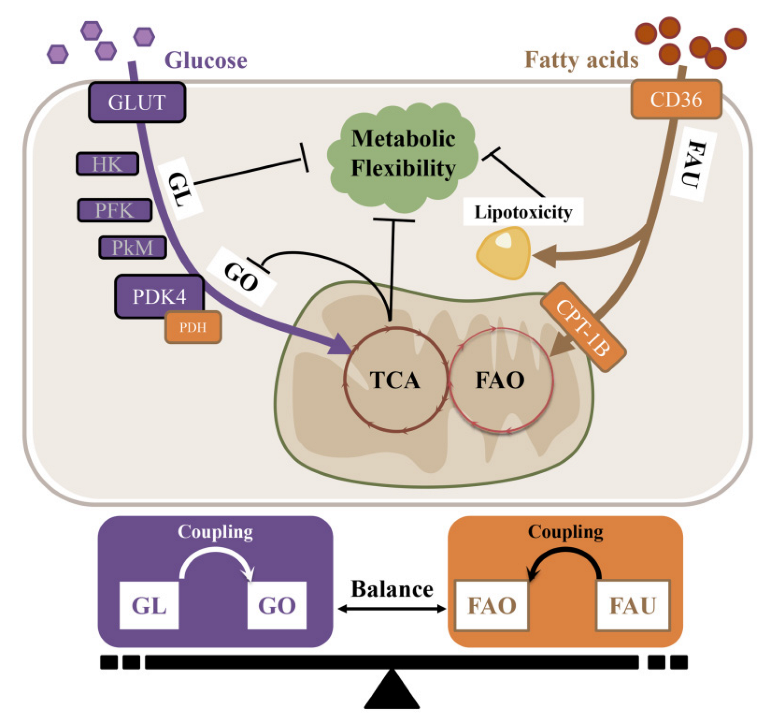
3. The Substrate-Metabolism Mechanism Underlying Metabolic Inflexibility
3.1. Substrate Metabolic Flexibility
3.1.1. Glucose Metabolic Inflexibility Underlying AF
-
Glucose metabolic abnormalities and pathogenesis
-
Insulin resistance and AF pathogenesis
-
Mechanisms of glucose metabolic inflexibility underlying AF
3.1.2. FAs Metabolic Inflexibility Underlying AF Pathogenesis
-
FAO and AF pathogenesis
-
Mechanisms of FAs’ metabolic inflexibility underlying AF
3.1.3. Amino Acids’ Metabolic Flexibility
-
BCAA and AF pathogenesis
-
Mechanisms of BCAA metabolic inflexibility underlying AF
3.1.4. Ketones and Metabolic Flexibility
-
Ketones and AF pathogenesis
-
Mechanisms of ketone metabolic inflexibility underlying AF
3.2. Metabolism Regulatory Signaling and Metabolic Flexibility
3.3. The Substrate-Metabolism Mechanism Underlying Metabolic Inflexibility and AF Pathogenesis
4. Anti-AF Strategies Targeting Metabolic Inflexibility
References
- Kelley, D.E.; He, J.; Menshikova, E.V.; Ritov, V.B. Dysfunction of Mitochondria in Human Skeletal Muscle in Type 2 Diabetes. Diabetes 2002, 51, 2944–2950.
- Smith, R.L.; Soeters, M.R.; Wust, R.; Houtkooper, R.H. Metabolic Flexibility as an Adaptation to Energy Resources and Requirements in Health and Disease. Endocr. Rev. 2018, 39, 489–517.
- Abe, J.; Abriel, H.; Accili, E.A.; Acosta, D.; Allen, L.F.; Allen, T.J.; Antzelevitch, C.; Aoki, H.; Ardell, J.L.; Arita, M.; et al. Contributors. In Heart Physiology and Pathophysiology, 4th ed.; Sperelakis, N., Kurachi, Y., Terzic, A., Cohen, M.V., Eds.; Academic Press: San Diego, CA, USA, 2001; pp. xiii–xvii. ISBN 978-0-12-656975-9.
- Suga, H. Ventricular Energetics. Physiol. Rev. 1990, 70, 247–277.
- Kolwicz, S.J.; Purohit, S.; Tian, R. Cardiac Metabolism and Its Interactions with Contraction, Growth, and Survival of Cardiomyocytes. Circ. Res. 2013, 113, 603–616.
- Neely, J.R.; Morgan, H.E. Relationship between Carbohydrate and Lipid Metabolism and the Energy Balance of Heart Muscle. Annu. Rev. Physiol. 1974, 36, 413–459.
- Lopaschuk, G.D.; Jaswal, J.S. Energy Metabolic Phenotype of the Cardiomyocyte during Development, Differentiation, and Postnatal Maturation. J. Cardiovasc. Pharmacol. 2010, 56, 130–140.
- Stanley, W.C.; Recchia, F.A.; Lopaschuk, G.D. Myocardial Substrate Metabolism in the Normal and Failing Heart. Physiol. Rev. 2005, 85, 1093–1129.
- Wentz, A.E.; D’Avignon, D.A.; Weber, M.L.; Cotter, D.G.; Doherty, J.M.; Kerns, R.; Nagarajan, R.; Reddy, N.; Sambandam, N.; Crawford, P.A. Adaptation of Myocardial Substrate Metabolism to a Ketogenic Nutrient Environment. J. Biol. Chem. 2010, 285, 24447–24456.
- Kolb, H.; Kempf, K.; Röhling, M.; Lenzen-Schulte, M.; Schloot, N.C.; Martin, S. Ketone Bodies: From Enemy to Friend and Guardian Angel. BMC Med. 2021, 19, 313.
- McGarry, J.D.; Foster, D.W. Regulation of Hepatic Fatty Acid Oxidation and Ketone Body Production. Annu. Rev. Biochem. 1980, 49, 395–420.
- Zaha, V.G.; Young, L.H. AMP-Activated Protein Kinase Regulation and Biological Actions in the Heart. Circ. Res. 2012, 111, 800–814.
- Lehman, J.J.; Barger, P.M.; Kovacs, A.; Saffitz, J.E.; Medeiros, D.M.; Kelly, D.P. Peroxisome Proliferator-Activated Receptor Gamma Coactivator-1 Promotes Cardiac Mitochondrial Biogenesis. J. Clin. Invest. 2000, 106, 847–856.
- Francis, G.A.; Annicotte, J.S.; Auwerx, J. PPAR-Alpha Effects on the Heart and Other Vascular Tissues. Am. J. Physiol. Heart Circ. Physiol. 2003, 285, H1–H9.
- Miyamoto, S.; Murphy, A.N.; Brown, J.H. Akt Mediated Mitochondrial Protection in the Heart: Metabolic and Survival Pathways to the Rescue. J. Bioenerg. Biomembr. 2009, 41, 169–180.
- Cerychova, R.; Pavlinkova, G. HIF-1, Metabolism, and Diabetes in the Embryonic and Adult Heart. Front. Endocrinol. 2018, 9, 460.
- Semenza, G.L. Hypoxia-Inducible Factor 1: Regulator of Mitochondrial Metabolism and Mediator of Ischemic Preconditioning. Biochim. Biophys. Acta 2011, 1813, 1263–1268.
- Aucouturier, J.; Duché, P.; Timmons, B.W. Metabolic Flexibility and Obesity in Children and Youth: Metabolic Flexibility in Youth. Obes. Rev. 2011, 12, e44–e53.
- Stull, A.J.; Galgani, J.E.; Johnson, W.D.; Cefalu, W.T. The Contribution of Race and Diabetes Status to Metabolic Flexibility in Humans. Metabolism 2010, 59, 1358–1364.
- Karwi, Q.G.; Uddin, G.M.; Ho, K.L.; Lopaschuk, G.D. Loss of Metabolic Flexibility in the Failing Heart. Front. Cardiovasc. Med. 2018, 5, 68.
- Huxley, R.R.; Filion, K.B.; Konety, S.; Alonso, A. Meta-Analysis of Cohort and Case-Control Studies of Type 2 Diabetes Mellitus and Risk of Atrial Fibrillation. Am. J. Cardiol. 2011, 108, 56–62.
- Dublin, S.; Glazer, N.L.; Smith, N.L.; Psaty, B.M.; Lumley, T.; Wiggins, K.L.; Page, R.L.; Heckbert, S.R. Diabetes Mellitus, Glycemic Control, and Risk of Atrial Fibrillation. J. Gen. Intern. Med. 2010, 25, 853–858.
- Qi, W.; Zhang, N.; Korantzopoulos, P.; Letsas, K.P.; Cheng, M.; Di, F.; Tse, G.; Liu, T.; Li, G. Serum Glycated Hemoglobin Level as a Predictor of Atrial Fibrillation: A Systematic Review with Meta-Analysis and Meta-Regression. PLoS ONE 2017, 12, e0170955.
- Saito, S.; Teshima, Y.; Fukui, A.; Kondo, H.; Nishio, S.; Nakagawa, M.; Saikawa, T.; Takahashi, N. Glucose Fluctuations Increase the Incidence of Atrial Fibrillation in Diabetic Rats. Cardiovasc. Res. 2014, 104, 5–14.
- Gu, J. Impact of Long-Term Glycemic Variability on Development of Atrial Fibrillation in Type 2 Diabetic Patients. Anatol. J. Cardiol. 2017, 18, 410–416.
- Chao, T.-F.; Suenari, K.; Chang, S.-L.; Lin, Y.-J.; Lo, L.-W.; Hu, Y.-F.; Tuan, T.-C.; Tai, C.-T.; Tsao, H.-M.; Li, C.-H.; et al. Atrial Substrate Properties and Outcome of Catheter Ablation in Patients With Paroxysmal Atrial Fibrillation Associated With Diabetes Mellitus or Impaired Fasting Glucose. Am. J. Cardiol. 2010, 106, 1615–1620.
- Lee, Y.; Cha, S.J.; Park, J.H.; Shin, J.H.; Lim, Y.H.; Park, H.C.; Shin, J.; Kim, C.K.; Park, J.K. Association between Insulin Resistance and Risk of Atrial Fibrillation in Non-Diabetics. Eur. J. Prev. Cardiol. 2020, 27, 1934–1941.
- Johnson, L.S.; Juhlin, T.; Engström, G.; Nilsson, P.M. Low Fasting Plasma Insulin Is Associated with Atrial Fibrillation in Men from a Cohort Study—The Malmö Preventive Project. BMC Cardiovasc. Disord. 2014, 14, 107.
- Fontes, J.D.; Lyass, A.; Massaro, J.M.; Rienstra, M.; Dallmeier, D.; Schnabel, R.B.; Wang, T.J.; Vasan, R.S.; Lubitz, S.A.; Magnani, J.W.; et al. Insulin Resistance and Atrial Fibrillation (from the Framingham Heart Study). Am. J. Cardiol. 2012, 109, 87–90.
- Garg, P.K.; Biggs, M.L.; Kaplan, R.; Kizer, J.R.; Heckbert, S.R.; Mukamal, K.J. Fasting and Post-Glucose Load Measures of Insulin Resistance and Risk of Incident Atrial Fibrillation: The Cardiovascular Health Study. Nutr. Metab. Cardiovasc. 2018, 28, 716–721.
- Chan, Y.-H.; Chang, G.-J.; Lai, Y.-J.; Chen, W.-J.; Chang, S.-H.; Hung, L.-M.; Kuo, C.-T.; Yeh, Y.-H. Atrial Fibrillation and Its Arrhythmogenesis Associated with Insulin Resistance. Cardiovasc. Diabetol. 2019, 18, 125.
- Polina, I.; Jansen, H.J.; Li, T.; Moghtadaei, M.; Bohne, L.J.; Liu, Y.; Krishnaswamy, P.; Egom, E.E.; Belke, D.D.; Rafferty, S.A.; et al. Loss of Insulin Signaling May Contribute to Atrial Fibrillation and Atrial Electrical Remodeling in Type 1 Diabetes. Proc. Natl. Acad. Sci. USA 2020, 117, 7990–8000.
- Maria, Z.; Campolo, A.R.; Scherlag, B.J.; Ritchey, J.W.; Lacombe, V.A. Insulin Treatment Reduces Susceptibility to Atrial Fibrillation in Type 1 Diabetic Mice. Front. Cardiovasc. Med. 2020, 7, 134.
- Maria, Z.; Campolo, A.R.; Scherlag, B.J.; Ritchey, J.W.; Lacombe, V.A. Dysregulation of Insulin-Sensitive Glucose Transporters During Insulin Resistance-Induced Atrial Fibrillation. BBA Mol. Basis Dis. 2018, 1864, 987–996.
- Liu, Y.; Bai, F.; Liu, N.; Ouyang, F.; Liu, Q. The Warburg Effect: A New Insight into Atrial Fibrillation. Clin. Chim. Acta 2019, 499, 4–12.
- Zhang, S.; Hulver, M.W.; McMillan, R.P.; Cline, M.A.; Gilbert, E.R. The Pivotal Role of Pyruvate Dehydrogenase Kinases in Metabolic Flexibility. Nutr. Metab. 2014, 11, 10.
- Tao, R.; Xiong, X.; Harris, R.A.; White, M.F.; Dong, X.C. Genetic Inactivation of Pyruvate Dehydrogenase Kinases Improves Hepatic Insulin Resistance Induced Diabetes. PLoS ONE 2013, 8, e71997.
- Hu, H.J.; Zhang, C.; Tang, Z.H.; Qu, S.L.; Jiang, Z.S. Regulating the Warburg Effect on Metabolic Stress and Myocardial Fibrosis Remodeling and Atrial Intracardiac Waveform Activity Induced by Atrial Fibrillation. Biochem. Biophys. Res. Commun. 2019, 516, 653–660.
- Simoneau, J.-A.; Kelley, D.E. Altered Glycolytic and Oxidative Capacities of Skeletal Muscle Contribute to Insulin Resistance in NIDDM. J. Appl. Physiol. 1997, 83, 166–171.
- Forouhi, N.G.; Koulman, A.; Sharp, S.J.; Imamura, F.; Kroger, J.; Schulze, M.B.; Crowe, F.L.; Huerta, J.M.; Guevara, M.; Beulens, J.W.; et al. Differences in the Prospective Association between Individual Plasma Phospholipid Saturated Fatty Acids and Incident Type 2 Diabetes: The EPIC-InterAct Case-Cohort Study. Lancet Diabetes Endocrinol. 2014, 2, 810–818.
- Zhao, G.; Cheng, D.; Wang, Y.; Cao, Y.; Xiang, S.; Yu, Q. A Metabolomic Study for Chronic Heart Failure Patients Based on a Dried Blood Spot Mass Spectrometry Approach. RSC Adv. 2020, 10, 19621–19628.
- Pararasa, C.; Ikwuobe, J.; Shigdar, S.; Boukouvalas, A.; Nabney, I.T.; Brown, J.E.; Devitt, A.; Bailey, C.J.; Bennett, S.J.; Griffiths, H.R. Age-Associated Changes in Long-Chain Fatty Acid Profile during Healthy Aging Promote pro-Inflammatory Monocyte Polarization via PPARgamma. Aging Cell 2016, 15, 128–139.
- Fretts, A.M.; Mozaffarian, D.; Siscovick, D.S.; Djousse, L.; Heckbert, S.R.; King, I.B.; McKnight, B.; Sitlani, C.; Sacks, F.M.; Song, X.; et al. Plasma Phospholipid Saturated Fatty Acids and Incident Atrial Fibrillation: The Cardiovascular Health Study. J. Am. Heart Assoc. 2014, 3, e000889.
- Jensen, P.N.; Fretts, A.M.; Hoofnagle, A.N.; Sitlani, C.M.; McKnight, B.; King, I.B.; Siscovick, D.S.; Psaty, B.M.; Heckbert, S.R.; Mozaffarian, D.; et al. Plasma Ceramides and Sphingomyelins in Relation to Atrial Fibrillation Risk: The Cardiovascular Health Study. J. Am. Heart Assoc. 2020, 9, e012853.
- Steinbusch, L.K.; Schwenk, R.W.; Ouwens, D.M.; Diamant, M.; Glatz, J.F.; Luiken, J.J. Subcellular Trafficking of the Substrate Transporters GLUT4 and CD36 in Cardiomyocytes. Cell Mol. Life Sci. 2011, 68, 2525–2538.
- Lenski, M.; Schleider, G.; Kohlhaas, M.; Adrian, L.; Adam, O.; Tian, Q.; Kaestner, L.; Lipp, P.; Lehrke, M.; Maack, C.; et al. Arrhythmia Causes Lipid Accumulation and Reduced Glucose Uptake. Basic Res. Cardiol. 2015, 110, 40.
- Koonen, D.P.; Febbraio, M.; Bonnet, S.; Nagendran, J.; Young, M.E.; Michelakis, E.D.; Dyck, J.R. CD36 Expression Contributes to Age-Induced Cardiomyopathy in Mice. Circulation 2007, 116, 2139–2147.
- Shingu, Y.; Yokota, T.; Takada, S.; Niwano, H.; Ooka, T.; Katoh, H.; Tachibana, T.; Kubota, S.; Matsui, Y. Decreased Gene Expression of Fatty Acid Binding Protein 3 in the Atrium of Patients with New Onset of Atrial Fibrillation in Cardiac Perioperative Phase. J. Cardiol. 2018, 71, 65–70.
- Barth, A.S.; Merk, S.; Arnoldi, E.; Zwermann, L.; Kloos, P.; Gebauer, M.; Steinmeyer, K.; Bleich, M.; Kääb, S.; Hinterseer, M.; et al. Reprogramming of the Human Atrial Transcriptome in Permanent Atrial Fibrillation: Expression of a Ventricular-like Genomic Signature. Circ. Res. 2005, 96, 1022–1029.
- Tu, T.; Zhou, S.; Liu, Z.; Li, X.; Liu, Q. Quantitative Proteomics of Changes in Energy Metabolism-Related Proteins in Atrial Tissue from Valvular Disease Patients with Permanent Atrial Fibrillation. Circ. J. 2014, 78, 993–1001.
- Zhang, Y.; Fu, Y.; Jiang, T.; Liu, B.; Sun, H.; Zhang, Y.; Fan, B.; Li, X.; Qin, X.; Zheng, Q. Enhancing Fatty Acids Oxidation via L-Carnitine Attenuates Obesity-Related Atrial Fibrillation and Structural Remodeling by Activating AMPK Signaling and Alleviating Cardiac Lipotoxicity. Front. Pharmacol. 2021, 12, 771940.
- D’Souza, K.; Nzirorera, C.; Kienesberger, P.C. Lipid Metabolism and Signaling in Cardiac Lipotoxicity. Biochim. Biophys. Acta 2016, 1861, 1513–1524.
- Sato, S.; Suzuki, J.; Hirose, M.; Yamada, M.; Zenimaru, Y.; Nakaya, T.; Ichikawa, M.; Imagawa, M.; Takahashi, S.; Ikuyama, S.; et al. Cardiac Overexpression of Perilipin 2 Induces Atrial Steatosis, Connexin 43 Remodeling, and Atrial Fibrillation in Aged Mice. Am. J. Physiol. Endocrinol. Metab. 2019, 317, E1193–E1204.
- Kwak, H.-B. Exercise and Obesity-Induced Insulin Resistance in Skeletal Muscle. Integr. Med. Res. 2013, 2, 131–138.
- Zhelev, Z.; Aoki, I.; Lazarova, D.; Vlaykova, T.; Higashi, T.; Bakalova, R. A “Weird” Mitochondrial Fatty Acid Oxidation as a Metabolic “Secret” of Cancer. Oxid. Med. Cell Longev. 2022, 2022, 1–38.
- Taegtmeyer, H.; Harinstein, M.E.; Gheorghiade, M. More than Bricks and Mortar: Comments on Protein and Amino Acid Metabolism in the Heart. Am. J. Cardiol. 2008, 101, 3E–7E.
- Nie, C.; He, T.; Zhang, W.; Zhang, G.; Ma, X. Branched Chain Amino Acids: Beyond Nutrition Metabolism. Int. J. Mol. Sci. 2018, 19, 954.
- Yu, L.-M.; Dong, X.; Zhao, J.-K.; Xu, Y.-L.; Xu, D.-Y.; Xue, X.-D.; Zhou, Z.-J.; Huang, Y.-T.; Zhao, Q.-S.; Luo, L.-Y.; et al. Activation of PKG-CREB-KLF15 by Melatonin Attenuates Angiotensin II-Induced Vulnerability to Atrial Fibrillation via Enhancing Branched-Chain Amino Acids Catabolism. Free Radical Biol. Med. 2022, 178, 202–214.
- Wang, W.; Zhang, F.; Xia, Y.; Zhao, S.; Yan, W.; Wang, H.; Lee, Y.; Li, C.; Zhang, L.; Lian, K.; et al. Defective Branched Chain Amino Acid Catabolism Contributes to Cardiac Dysfunction and Remodeling Following Myocardial Infarction. Am. J. Physiol. Heart C 2016, 311, H1160–H1169.
- Sun, H.; Olson, K.C.; Gao, C.; Prosdocimo, D.A.; Zhou, M.; Wang, Z.; Jeyaraj, D.; Youn, J.-Y.; Ren, S.; Liu, Y.; et al. Catabolic Defect of Branched-Chain Amino Acids Promotes Heart Failure. Circulation 2016, 133, 2038–2049.
- Yoon, M.-S. The Emerging Role of Branched-Chain Amino Acids in Insulin Resistance and Metabolism. Nutrients 2016, 8, 405.
- Lynch, C.J.; Adams, S.H. Branched-Chain Amino Acids in Metabolic Signalling and Insulin Resistance. Nat. Rev. Endocrinol. 2014, 10, 723–736.
- Bing, R.J. The Metabolism of the Heart. Harvey Lect. 1954, 50, 27–70.
- Rudolph, W.; Maas, D.; Richter, J.; Hasinger, F.; Hofmann, H.; Dohrn, P. On the Significance of Acetoacetate and Beta-Hydroxybutyrate in Human Myocardial Metabolism. Klin. Wochenschr. 1965, 43, 445–451.
- Harvey, K.L.; Holcomb, L.E.; Kolwicz, S.J. Ketogenic Diets and Exercise Performance. Nutrients 2019, 11, 2296.
- Lommi, J.; Koskinen, P.; Naveri, H.; Harkonen, M.; Kupari, M. Heart Failure Ketosis. J. Intern. Med. 1997, 242, 231–238.
- Lommi, J.; Kupari, M.; Koskinen, P.; Naveri, H.; Leinonen, H.; Pulkki, K.; Harkonen, M. Blood Ketone Bodies in Congestive Heart Failure. J. Am. Coll. Cardiol. 1996, 28, 665–672.
- Rudolph, W.; Schinz, A. Studies on Myocardial Blood Flow, Oxygen Consumption, and Myocardial Metabolism in Patients with Cardiomyopathy. Recent Adv. Stud. Cardiac. Struct. Metab. 1973, 2, 739–749.
- Mayr, M.; Yusuf, S.; Weir, G.; Chung, Y.L.; Mayr, U.; Yin, X.; Ladroue, C.; Madhu, B.; Roberts, N.; De Souza, A.; et al. Combined Metabolomic and Proteomic Analysis of Human Atrial Fibrillation. J. Am. Coll. Cardiol. 2008, 51, 585–594.
- Zou, Z.; Sasaguri, S.; Rajesh, K.G.; Suzuki, R. Dl-3-Hydroxybutyrate Administration Prevents Myocardial Damage after Coronary Occlusion in Rat Hearts. Am. J. Physiol. Heart Circ. Physiol. 2002, 283, H1968–H1974.
- Sato, K.; Kashiwaya, Y.; Keon, C.A.; Tsuchiya, N.; King, M.T.; Radda, G.K.; Chance, B.; Clarke, K.; Veech, R.L. Insulin, Ketone Bodies, and Mitochondrial Energy Transduction. FASEB J. 1995, 9, 651–658.
- Watt, M.J.; Miotto, P.M.; De Nardo, W.; Montgomery, M.K. The Liver as an Endocrine Organ—Linking NAFLD and Insulin Resistance. Endocr. Rev. 2019, 40, 1367–1393.
- Goday, A.; Bellido, D.; Sajoux, I.; Crujeiras, A.B.; Burguera, B.; García-Luna, P.P.; Oleaga, A.; Moreno, B.; Casanueva, F.F. Short-Term Safety, Tolerability and Efficacy of a Very Low-Calorie-Ketogenic Diet Interventional Weight Loss Program versus Hypocaloric Diet in Patients with Type 2 Diabetes Mellitus. Nutr. Diabetes 2016, 6, e230.
- Ellenbroek, J.H.; van Dijck, L.; Töns, H.A.; Rabelink, T.J.; Carlotti, F.; Ballieux, B.E.P.B.; de Koning, E.J.P. Long-Term Ketogenic Diet Causes Glucose Intolerance and Reduced β- and α-Cell Mass but No Weight Loss in Mice. Am. J. Physiol. Endocr. Metab. 2014, 306, E552–E558.
- Kimura, I.; Inoue, D.; Maeda, T.; Hara, T.; Ichimura, A.; Miyauchi, S.; Kobayashi, M.; Hirasawa, A.; Tsujimoto, G. Short-Chain Fatty Acids and Ketones Directly Regulate Sympathetic Nervous System via G Protein-Coupled Receptor 41 (GPR41). Proc. Natl. Acad. Sci. USA 2011, 108, 8030–8035.
- Xu, S.; Tao, H.; Cao, W.; Cao, L.; Lin, Y.; Zhao, S.M.; Xu, W.; Cao, J.; Zhao, J.Y. Ketogenic Diets Inhibit Mitochondrial Biogenesis and Induce Cardiac Fibrosis. Signal Transduct. Target Ther. 2021, 6, 54.
- Hasselbaink, D.M.; Glatz, J.F.; Luiken, J.J.; Roemen, T.H.; Van der Vusse, G.J. Ketone Bodies Disturb Fatty Acid Handling in Isolated Cardiomyocytes Derived from Control and Diabetic Rats. Biochem. J. 2003, 371, 753–760.
- Vanoverschelde, J.L.; Wijns, W.; Kolanowski, J.; Bol, A.; Decoster, P.M.; Michel, C.; Cogneau, M.; Heyndrickx, G.R.; Essamri, B.; Melin, J.A. Competition between Palmitate and Ketone Bodies as Fuels for the Heart: Study with Positron Emission Tomography. Am. J. Physiol. 1993, 264, H701–H707.
- Ozcan, C.; Battaglia, E.; Young, R.; Suzuki, G. LKB1 Knockout Mouse Develops Spontaneous Atrial Fibrillation and Provides Mechanistic Insights Into Human Disease Process. JAHA 2015, 4, e001733.
- Deshmukh, A.; Ghannam, M.; Liang, J.; Saeed, M.; Cunnane, R.; Ghanbari, H.; Latchamsetty, R.; Crawford, T.; Batul, S.A.; Chung, E.; et al. Effect of Metformin on Outcomes of Catheter Ablation for Atrial Fibrillation. J. Cardiovasc. Electrophysiol. 2021, 32, 1232–1239.
- Ostropolets, A.; Elias, P.A.; Reyes, M.V.; Wan, E.Y.; Pajvani, U.B.; Hripcsak, G.; Morrow, J.P. Metformin Is Associated With a Lower Risk of Atrial Fibrillation and Ventricular Arrhythmias Compared With Sulfonylureas: An Observational Study. Circ. Arrhythm. Electrophysiol. 2021, 14, e009115.
- Liu, G.Z.; Hou, T.T.; Yuan, Y.; Hang, P.Z.; Zhao, J.J.; Sun, L.; Zhao, G.Q.; Zhao, J.; Dong, J.M.; Wang, X.B.; et al. Fenofibrate Inhibits Atrial Metabolic Remodelling in Atrial Fibrillation through PPAR-Alpha/Sirtuin 1/PGC-1alpha Pathway. Br. J. Pharmacol. 2016, 173, 1095–1109.
- Yu, J.; Li, W.; Li, Y.; Zhao, J.; Wang, L.; Dong, D.; Pan, Z.; Yang, B. Activation of Beta(3)-Adrenoceptor Promotes Rapid Pacing-Induced Atrial Electrical Remodeling in Rabbits. Cell Physiol. Biochem. 2011, 28, 87–96.
- Bai, F.; Liu, Y.; Tu, T.; Li, B.; Xiao, Y.; Ma, Y.; Qin, F.; Xie, J.; Zhou, S.; Liu, Q. Metformin Regulates Lipid Metabolism in a Canine Model of Atrial Fibrillation through AMPK/PPAR-Alpha/VLCAD Pathway. Lipids Health Dis. 2019, 18, 109.
- Ozcan, C.; Dixit, G.; Li, Z. Activation of AMP-Activated Protein Kinases Prevents Atrial Fibrillation. J. Cardiovasc. Trans. Res. 2021, 14, 492–502.
- Goodpaster, B.H.; Sparks, L.M. Metabolic Flexibility in Health and Disease. Cell Metab. 2017, 25, 1027–1036.
- Williams, N.C.; O’Neill, L.A.J. A Role for the Krebs Cycle Intermediate Citrate in Metabolic Reprogramming in Innate Immunity and Inflammation. Front. Immunol. 2018, 9, 141.
- Wijesurendra, R.S.; Casadei, B. Mechanisms of Atrial Fibrillation. Heart 2019, 105, 1860–1867.

