Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Yonghui Dong | -- | 939 | 2022-07-29 10:13:11 | | | |
| 2 | Beatrix Zheng | + 1908 word(s) | 2847 | 2022-08-01 03:02:44 | | | | |
| 3 | Beatrix Zheng | Meta information modification | 2847 | 2022-08-01 03:15:17 | | | | |
| 4 | Beatrix Zheng | + 9 word(s) | 2856 | 2022-08-01 03:16:21 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Dong, Y.; Wu, X.; Han, L.; Bian, J.; He, C.; El-Omar, E.; Gong, L.; Wang, M. Structure and Physiological Activities of Anthocyanins. Encyclopedia. Available online: https://encyclopedia.pub/entry/25647 (accessed on 01 August 2026).
Dong Y, Wu X, Han L, Bian J, He C, El-Omar E, et al. Structure and Physiological Activities of Anthocyanins. Encyclopedia. Available at: https://encyclopedia.pub/entry/25647. Accessed August 01, 2026.
Dong, Yonghui, Xue Wu, Lin Han, Ji Bian, Caian He, Emad El-Omar, Lan Gong, Min Wang. "Structure and Physiological Activities of Anthocyanins" Encyclopedia, https://encyclopedia.pub/entry/25647 (accessed August 01, 2026).
Dong, Y., Wu, X., Han, L., Bian, J., He, C., El-Omar, E., Gong, L., & Wang, M. (2022, July 29). Structure and Physiological Activities of Anthocyanins. In Encyclopedia. https://encyclopedia.pub/entry/25647
Dong, Yonghui, et al. "Structure and Physiological Activities of Anthocyanins." Encyclopedia. Web. 29 July, 2022.
Copy Citation
Anthocyanidin is a kind of water-soluble natural pigment that widely exists in natural plants. Like other natural flavonoids, anthocyanin has a C6-C3-C6 carbon skeleton. Due to the different carbon substituents (-OH, -OCH3) on the B ring, different types of anthocyanins were derived. The six common anthocyanins were Pelargonidin (Pg), Cyanidin (Cy), Delphinidin (Dp), Peonidin (Pn), Petunidin (Pt), and Malvidin (Mv). In addition to giving food a variety of bright colors, anthocyanin also has important biological activities, such as antioxidant, anti-inflammatory, and anti-aging effects, among others. A large number of studies have shown that dietary anthocyanins have a good preventive effect on cardiovascular diseases.
anthocyanin
gut microbiota
vascular endothelial cells senescence
clearance of senescence cells
cardioprotection
1. Structure of Anthocyanin
Anthocyanin is a water-soluble flavanol compound that widely exists in fruits, vegetables, and flowers, such as blueberry, sunflower, grape, pitaya, purple sweet potato, and purple cabbage. It is an extremely important secondary metabolite in plants. The structure of anthocyanin is mainly composed of C6-C3-C6 as the basic C skeleton. The differences between anthocyanin molecules are mainly due to the number of hydroxyl groups, the type and bonding position of sugars, and the type and bonding position of acyl groups of modified sugar molecules. There are six kinds of anthocyanins in plants. When positions 3, 5, and 7 of a and C rings are Oh, anthocyanins are aglycones, mainly including delphinidin (12%), cyanidin (50%), pelargonidin (12%), petunidin (7%), malvidin (7%), and peonidin (12%) (Figure 1 and Table 1). Delphinidin and its derivatives, petunidin and malvidin, are the sources of blue and purple, while cyanidin and pelargonidin are the main pigments of bright red fruits. Under natural conditions, the free anthocyanin is unstable, so it is rare that anthocyanin mainly exists in the form of glycoside. The hydroxyl at positions 3, 5, and 7 of anthocyanin can form anthocyanin through glycosidic bond with one or more monosaccharides (glucose, galactose, etc.), disaccharides (rutinose, etc.), or trisaccharides. Due to the different types, positions, and quantities of sugars that are glycosides of anthocyanin, the types of anthocyanin formed are also different. At present, there are more than 250 known natural anthocyanins.
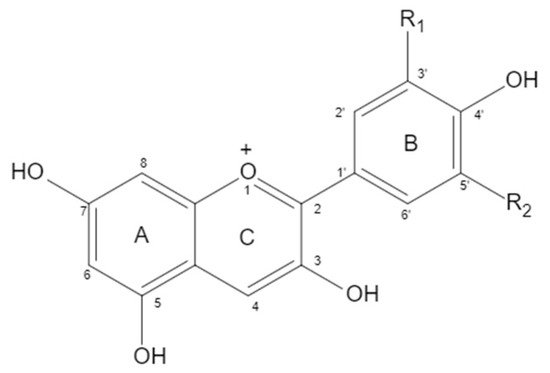
Figure 1. Chemical structure of anthocyanins.
Table 1. Chemical structure of anthocyanins.
| Anthocyanin | Proportion | Substituents | |
|---|---|---|---|
| R1 | R2 | ||
| Cyanidin | 50% | OH | H |
| Delphindin | 12% | OH | OH |
| Pelargonidin | 12% | H | H |
| Peonidin | 12% | OCH3 | H |
| Petunidin | 7% | OCH3 | OH |
| Malvidin | 7% | OCH3 | OCH3 |
2. Physiological Activities of Anthocyanins
2.1. Anti-Cancer
Cancer is a disease caused by uncontrolled growth and progressive development of abnormal cells, killing millions of people every year. By 2030, there will be more than 20 million new cancer cases. Anthocyanins have been shown to have the ability to inhibit the initiation, promotion, and progression of various cancers, such as colon cancer [1], liver and bladder cancer [2], breast cancer [3], brain cancer [4], kidney cancer and skin cancer [5], gastric cancer [6], and thyroid cancer [7]. The ability of anthocyanins to inhibit tumorigenesis and development is closely related to their ability to enhance antioxidant defense; exert anti-inflammatory effects; and interfere with ERK, JNK, PI3K/Akt, MAPK, and NF-κB signaling pathways. Yun et al. reported that purple grape anthocyanins prevented tumor-necrosis-factor-α-induced NF-κB activation by inhibiting IκBα phosphorylation and resisted the invasion of human colon cancer cells in a dose-dependent manner [8]. Fragoso et al. proved through experiments that cyanidin-3-O-rutinoside at 25 µmol/L can effectively reduce the motility of human colon adenocarcinoma cells, reduce the metastasis of cancer cells, and play an anticancer effect [9]. Mazewski et al. reported that anthocyanins extracted from purple and red maize enhanced the expression of apoptotic factors BAX, Bcl-2, cytochrome C, and TRAILR2/D5 and inhibited vascular endothelial growth factor in HCT-116 and HT-29 human colorectal cancer cell (Tie-2, ANGPT2, and PLG) expression to achieve anti-cancer efficacy [10]. The results of Lage et al. showed that black sweet cherry anthocyanins can inhibit the growth of breast cancer cells and have no toxicity to normal MCF-10A breast cells. Anthocyanins work against cancer by reducing oxidative stress, regulation of Akt/mTOR, p38, and survivin, preventing cancer cell proliferation and promoting apoptosis. Anthocyanins can significantly downregulate the mRNA expression of invasive/metastatic biomarkers (Sp1, Sp4, VCAM-1), and anthocyanins from black sweet cherry can effectively prevent and treat cancer [3]. Su et al. reported that hibiscus calyx anthocyanin could inhibit the growth, metastasis, and angiogenesis of B16-F1 cells by triggering PI3K/Akt and Ras/MAPK signaling pathways and downregulating the expression of VEGF and MMP-2/-9, which could effectively prevent and treat melanoma cancer [11]. Sugata et al. found that purple sweet potato anthocyanin blocked all stages of cell cycle by acting on cell cycle regulators (such as p53, p21, p27, Cyclin D1, and Cyclin A), thereby inhibiting the proliferation of breast cancer, colon cancer, and gastric cancer cells in a concentration- and time-dependent manner [12].
2.2. Anti-Inflammatory
Inflammation is usually regulated by the body to secrete inflammatory cytokines and mediators. Therefore, it is generally believed that the downregulation of factor secretion may contribute to the treatment of diseases such as inflammation [13]. Epidemiology and research have shown that anthocyanin has an anti-inflammatory effect and can improve a variety of inflammation-related diseases, such as colitis [14], periodontitis, pharyngitis, and postprandial inflammatory response. Anthocyanin can change the redox state of cells and affect redox-sensitive inflammatory mediators through Nrf2-ARE signal modulation [15]. Hou et al. showed that anthocyanin inhibited COX-2 by inhibiting C/EBP, AP-1, and NF-κB, thereby reducing the production of pro-inflammatory cytokines IL-1β, IL-6, IL-8, and TNF-α [16]. Min et al. found that cyanidin-3-glucoside showed an effective anti-inflammatory effect by regulating NF-κB and MAPK activity [17]. Studies have shown that fenugreein can inhibit HcPT degradation, p65 nuclear translocation, and JNK phosphorylation, showing an indigenous anti-inflammatory activity [18]. In general, B-ring o-dihydroxyphenyl anthocyanin, such as fayashinin and cyanidin, has strong anti-inflammatory activity, while geranium pigment, peony pigment, and kumquat pigment do not show the above activity without o-dihydroxy structure [19]. Aboonabi et al. showed that 320 mg anthocyanidin daily intake in people with metabolic syndrome can significantly inhibit the expression of NF-κB-pathway-related proinflammatory factor genes and enhance the expression of PPAR-γ gene to reduce the risk of inflammation [20]. Duarte et al. showed that geranium pigment-3-O-glucoside in strawberry could inhibit the activation of IkB-α and reduce the phosphorylation of JNK-MAPK, leading to the decrease in NF-κB and AP-1 activation factors in the inflammatory pathway stimulated by TLR4, indicating that geranium pigment-3-O-glucoside had an anti-inflammatory effect [21]. The study of Karnarathne and other studies have shown that anthocyanin from Hibiscus can inhibit the secretion of nitric oxide and prostaglandin E2 in LPS-induced endotoxic shock zebrafish, while down-regulating the expression of inducible nitric oxide synthase and cyclooxygenase 2. Furthermore, LPS inhibited the production of pro-inflammatory cytokines such as TNF-α, IL-6 and IL-12 in RAW 264.7 macrophages. Anthocyanin also inhibits LPS-induced TLR4 dimerization or cell surface formation, thereby reducing MyD88 growth and IRAK4 phosphorylation, thereby inhibiting NF-κB activity [22].
2.3. Anti-Oxidation
Humans produce free radicals during metabolism. Excessive free radicals can lead to lipid, protein, DNA, RNA, and sugar oxidation, which is closely related to cancer, Alzheimer′s disease, Parkinson′s disease, autoimmune deficiency, diabetes, obesity, and other diseases. As a natural plant pigment, anthocyanin not only can be used as a colorant, but also has prominent antioxidant activity. Anthocyanins can scavenge reactive oxygen species (ROS) and reactive nitrogen (RNS), such as superoxide anion (O2−), singlet oxygen (1O2), peroxide free radical (RCOO·), hydrogen peroxide, hydroxyl free radical (OH·), and peroxynitrite anion (ONOO−) [23]. The phenolic ring, hydroxyl side chain, and double bond in the glycosylation reaction of anthocyanin are helpful to scavenge free radicals. Compared with cyanidins and philoxerin, anthocyanin lacking O-phenyl structure in the B ring (sunflower pigment, geranium pigment, petunia pigment, and peony pigment) had low DPPH radical scavenging efficiency. Peonidin has methyl at 3′ position and OH at 4′ position, which is more active than pelargonidin. As reported by Fukumoto and Mazza, the hydroxyl at the third position of the B ring enhances the activity. Similarly, delphinidin with hydroxyl at 3′, 4′, and 5′ is more effective than cyanidin with hydroxyl at only 3′ and 4′ [24]. Harakotr et al. reported that the anthocyanin extract of purple corn had strong DPPH radical scavenging activity, and the anthocyanin content in the extract was positively correlated with antioxidant capacity [25]. Matera et al. reported that cyanidins in radish buds could significantly inhibit the automatic oxidation of linoleic acid and scavenge hydrogen-peroxide-free radicals [26]. Coklar et al. reported that anthocyanin extracts from Mahonia aquifolium (cyanidins, delphinidin, malvidin, peonidin, pelargonidin) had strong DPPH and ABTS radical scavenging ability and FRAP reduction ability [27]. Lu et al. fed D-galactose-induced aging mice black rice anthocyanin extract (cyanidin-3-O-glucoside). The activities of superoxide dismutase and catalase in mice were significantly improved, and the content of malondialdehyde and the activity of monoamine oxidase were reduced. Black rice anthocyanin extract showed a strong anti-aging effect in mice [28]. Huang et al. studied the antioxidant effect of main anthocyanins in blueberry on endothelial cells. The results showed that brocade pigment and its two glycosides decreased the levels of reactive oxygen species (ROS) and xanthine oxidase-1 (XO-1), but increased superoxide dismutase (SOD) and heme oxygenase-1 (HO-1). Moreover, the presence of glycoside greatly improved the antioxidant capacity of malvidin [29].
2.4. Protective Effect on the Liver
Daveri et al. fed high-fat diet mice with 40 mg anthocyanin/kg BW (cyanidins and delphinidins). The changes of chemokine MCP-1, cytokine TNF-α, macrophage marker F4/80, and enzyme NOS2 were measured. The results showed that anthocyanin played a role in preventing liver injury [30]. Jiang et al. showed that when carbon tetrachloride-induced liver injury mice were fed with cyanidin-3-O-glucoside 800 mg/kg BW, cyanidin-3-O-glucoside could significantly alleviate liver injury and prevent fibrosis in mice. Cyanidin-3-O-glucose can protect the liver by reducing liver oxidative stress, reducing liver cell apoptosis, inhibiting liver inflammatory response, and ultimately inhibiting the activation of liver star [31]. Arjinajarn et al. showed that the anthocyanin extract of riceberry bran could prevent gentamicin-induced liver injury in rats by inhibiting intracellular oxidative stress and the activation of NF-κB factor, reducing liver cell inflammation and apoptosis [32]. Zhang et al. found that purple sweet potato anthocyanin could effectively inhibit the production of reactive oxygen species in mice and inhibit the accumulation of liver fat induced by high-fat diet by activating adenosine-monophosphate-activated protein kinase (AMPK) signaling pathway [33]. Cai et al. studied the effects of different doses of purple sweet potato anthocyanin on the main liver function indexes, liver histological changes, and oxidation state of mice with alcoholic fatty liver, finding that medium dose of purple sweet potato anthocyanin had an obvious protective effect on the release of alanine aminotransferase (ALT) in the mice with liver injury [34].
2.5. Lowering Blood Glucose
Diabetes is a non-infectious endocrine metabolic disease that can lead to serious complications of various organs, and the number of patients with diabetes is increasing. Maintaining normal blood glucose level is a necessary condition for maintaining body function. In the human body, glucose homeostasis is controlled by various organs, including the pancreas, liver, and other tissues, as well as complex networks of hormones and neuropeptides. The pancreas plays a key role in glucose homeostasis by secreting hypoglycemic hormone insulin [35]. Purple corn anthocyanins have significant effects on β-cell function and insulin secretion, which can protect pancreatic β cells from high-glucose-induced oxidative stress and improve insulin secretion ability of β cells [36]. The liver is the main part of human body and plays a fundamental role in glycogen storage, plasma protein synthesis, and detoxification [37]. Studies have shown that anthocyanin-rich mulberry extract inhibits gluconeogenesis and stimulates glycogen synthesis by increasing AMPK phosphorylation in the liver [38].
2.6. Anti-Aging
Oxidative stress is one of the main inducing factors of aging, and excessive expression of inflammatory factors, DNA damage, and a series of inflammatory reactions activated by NLPR3 and NF-κB can also promote the aging of the body [39]. Many studies have shown that anthocyanin has an anti-aging effect. Jin et al. fed aged mice with anthocyanin from purple sweet potato and found that compared with the control group, anthocyanin from purple sweet potato could significantly reduce the serum MDA level and improve the activities of SOD and GSH-PX, and low-dose anthocyanin could achieve the same effect as the equivalent amount of vitamin, indicating that anthocyanin from purple sweet potato could play a role in delaying aging by improving antioxidant activity [40]. Wang et al. showed that Cy-3-glu and Pg-3-glu treatments could significantly inhibit the galactosidase in the aging process of human retinal pigment epithelium (RPE) cells induced by visible light irradiation and play a protective role in anti-aging [41]. Gao et al. found that Ribes meyeri anthocyanins can promote the proliferation of neural stem cells, improve cell senescence phenotype, reduce ROS and senescence-associated P16Ink4a gene expression levels, increase DNA synthesis, and prolong telomeres [42]. Wei et al. showed that anthocyanin could maintain the stability of the redox system in plasma and liver structure, as well as reduce the levels of inflammatory factors such as IL-1, IL-6, and TNF-α in the liver. At the same time, the decrease in the expression levels of sensors (ATM and ATR), media (H2AX and γ-H2AX), and effectors (Chk1, Chk2, p53 and p-p53) in the DNA damage signaling pathway indicate that anthocyanin can slow down aging by inhibiting DNA damage [43].
2.7. Other Effects
Qin et al. used purple sweet potato anthocyanin (PSPC 500 mg/kg/day) to orally take high-fat model mice. The results showed that PSPC corrected the abnormal metabolic indexes induced by HFD, including improving obesity, reducing fasting blood glucose concentration, and improving glucose tolerance [44]. Lee et al. found the effect of black soybean anthocyanin on obesity. The results showed that TC/HDLc/LDLc/HDLc of obese patients taking black soybean anthocyanin were significantly decreased [45]. Farrell et al. established a mouse model of hyperlipidemia and high-density lipoprotein dysfunction to explore and determine that an anthocyanin-rich blackcurrant extract (BEE) (13% anthocyanin) can prevent inflammation-related HDL functional damage and apolipoprotein E atherosclerosis. The results showed that the total cholesterol content in the aorta of mice was significantly decreased, and the aspartate aminotransferase (AST) and fasting blood glucose were decreased, indicating that blackberry may affect chronic inflammation-related HDL dysfunction by affecting liver gene expression [46]. In addition, studies have shown that purple sweet potato anthocyanin has a protective effect on the kidneys. Qun and other studies have found that purple sweet potato anthocyanin can significantly improve kidney injury in mice fed with high fat diet by reducing the production of AGEs and ROS and improving insulin sensitivity. Its protective effect is played by inhibiting the expression of TXNIP and RAGE and further inhibiting the activation of NLRP3 inflammasome and IKKb/NFκB pathway [39].
The physiological activities of anthocyanins are summarized in Table 2.
Table 2. Physiological activities of anthocyanins.
| Effects | Source | Mechanisms | Ref. | |
|---|---|---|---|---|
| Anti-cancer | colon cancer | Purple grape anthocyanins |
|
[8] |
| colon cancer | Cyanidin-3-O-rutinoside |
|
[9] | |
| colon cancer | Purple and red maize anthocyanins |
|
[10] | |
| breast cancer | Black sweet cherry anthocyanins |
|
[3] | |
| melanoma cancer | Hibiscus calyx anthocyanin |
|
[11] | |
| Purple sweet potato anthocyanin |
|
[12] | ||
| Anti-inflammatory |
|
[15] | ||
|
[16] [19] |
|||
| Cyanidin-3-glucoside |
|
[17] | ||
| Geranium pigment-3-O-glucoside in strawberry |
|
[21] | ||
| Hibiscus anthocyanin |
|
[22] | ||
| Anti-oxidation | Purple corn anthocyanin |
|
[25] | |
| Cyanidins in radish buds |
|
[26] | ||
| Mahonia aquifolium anthocyanin |
|
[27] | ||
| Black rice anthocyanin extract (cyanidin-3-O-glucoside) |
|
[28] | ||
| Blueberry anthocyanins |
|
[29] | ||
| Protective effect on liver | Cyanidin-3-O-glucoside |
|
[31] | |
| Riceberry bran anthocyanin |
|
[32] | ||
| Purple sweet potato anthocyanin |
|
[33] | ||
| Purple sweet potato anthocyanin |
|
[34] | ||
| Lowering blood glucose | Purple corn anthocyanins |
|
[47] | |
| Mulberry anthocyanin |
|
[36] | ||
| Anti-aging | Purple sweet potato anthocyanin |
|
[40] | |
| Cy-3-glu Pg-3-glu |
|
[41] | ||
| Ribes meyeri anthocyanins |
|
[42] | ||
|
[43] | |||
References
- Goncalves, A.C.; Rodrigues, M.; Santos, A.O.; Alves, G.; Silva, L.R. Antioxidant Status, Antidiabetic Properties and Effects on Caco-2 Cells of Colored and Non-Colored Enriched Extracts of Sweet Cherry Fruits. Nutrients 2018, 10, 1688.
- Masci, A.; Coccia, A.; Lendaro, E.; Mosca, L.; Paolicelli, P.; Cesa, S. Evaluation of different extraction methods from pomegranate whole fruit or peels and the antioxidant and antiproliferative activity of the polyphenolic fraction. Food Chem. 2016, 202, 59–69.
- Lage, N.N.; Layosa, M.A.A.; Arbizu, S.; Chew, B.P.; Pedrosa, M.L.; Mertens-Talcott, S.; Talcott, S.; Noratto, G.D. Dark sweet cherry (Prunus avium) phenolics enriched in anthocyanins exhibit enhanced activity against the most aggressive breast cancer subtypes without toxicity to normal breast cells. J. Funct. Foods 2020, 64, 103710.
- Hogan, S.; Chung, H.; Zhang, L.; Li, J.; Lee, Y.; Dai, Y.; Zhou, K. Antiproliferative and antioxidant properties of anthocyanin-rich extract from açai. Food Chem. 2010, 118, 208–214.
- Vilkickyte, G.; Raudone, L.; Petrikaite, V. Phenolic Fractions from Vaccinium vitis-idaea L. and Their Antioxidant and Anticancer Activities Assessment. Antioxidants 2020, 9, 1261.
- Bowen-Forbes, C.S.; Zhang, Y.; Nair, M.G. Anthocyanin content, antioxidant, anti-inflammatory and anticancer properties of blackberry and raspberry fruits. J. Food Compos. Anal. 2010, 23, 554–560.
- Long, H.L.; Zhang, F.F.; Wang, H.L.; Yang, W.S.; Hou, H.T.; Yu, J.K.; Liu, B. Mulberry anthocyanins improves thyroid cancer progression mainly by inducing apoptosis and autophagy cell death. Kaohsiung J. Med. Sci. 2018, 34, 255–262.
- Yun, J.W.; Lee, W.S.; Kim, M.J.; Lu, J.N.; Kang, M.H.; Kim, H.G.; Kim, D.C.; Choi, E.J.; Choi, J.Y.; Kim, H.G.; et al. Characterization of a profile of the anthocyanins isolated from Vitis coignetiae Pulliat and their anti-invasive activity on HT-29 human colon cancer cells. Food Chem. Toxicol. 2010, 48, 903–909.
- Fragoso, M.F.; Romualdo, G.R.; Vanderveer, L.A.; Franco-Barraza, J.; Cukierma, E.; Clapper, M.L.; Carvalho, R.F.; Barbisa, L.F. Lyophilized acai pulp (Euterpe oleracea Mart) attenuates colitis-associated colon carcinogenesis while its main anthocyanin has the potential to affect the motility of colon cancer cells. Food Chem. Toxicol. 2018, 121, 237–245.
- Mazewski, C.; Liang, K.; de Mejia, E.G. Inhibitory potential of anthocyanin-rich purple and red corn extracts on human colorectal cancer cell proliferation in vitro. J. Funct. Foods 2017, 34, 254–265.
- Su, C.-C.; Wang, C.-J.; Huang, K.-H.; Lee, Y.-J.; Chan, W.-M.; Chang, Y.-C. Anthocyanins from Hibiscus sabdariffa calyx attenuate in vitro and in vivo melanoma cancer metastasis. J. Funct. Foods 2018, 48, 614–631.
- Lim, S.; Xu, J.; Kim, J.; Chen, T.Y.; Su, X.; Standard, J.; Carey, E.; Griffin, J.; Herndon, B.; Katz, B.; et al. Role of anthocyanin-enriched purple-fleshed sweet potato p40 in colorectal cancer prevention. Mol. Nutr. Food Res. 2013, 57, 1908–1917.
- Teng, H.; Fang, T.; Lin, Q.; Song, H.; Liu, B.; Chen, L. Red raspberry and its anthocyanins: Bioactivity beyond antioxidant capacity. Trends Food Sci. Technol. 2017, 66, 153–165.
- Akiyama, S.; Nesumi, A.; Maeda-Yamamoto, M.; Uehara, M.; Murakami, A. Effects of anthocyanin-rich tea “Sunrouge” on dextran sodium sulfate-induced colitis in mice. Biofactors 2012, 38, 226–233.
- Folmer, F.; Basavaraju, U.; Jaspars, M.; Hold, G.; El-Omar, E.; Dicato, M.; Diederich, M. Anticancer effects of bioactive berry compounds. Phytochem. Rev. 2013, 13, 295–322.
- Hou, D.X.; Kai, K.; Li, J.J.; Lin, S.; Terahara, N.; Wakamatsu, M.; Fujii, M.; Young, M.R.; Colburn, N. Anthocyanidins inhibit activator protein 1 activity and cell transformation: Structure-activity relationship and molecular mechanisms. Carcinogenesis 2004, 25, 29–36.
- Min, S.W.; Ryu, S.N.; Kim, D.H. Anti-inflammatory effects of black rice, cyanidin-3-O-beta-D-glycoside, and its metabolites, cyanidin and protocatechuic acid. Int. Immunopharmacol. 2010, 10, 959–966.
- Hou, D.X.; Yanagita, T.; Uto, T.; Masuzaki, S.; Fujii, M. Anthocyanidins inhibit cyclooxygenase-2 expression in LPS-evoked macrophages: Structure-activity relationship and molecular mechanisms involved. Biochem. Pharmacol. 2005, 70, 417–425.
- Chen, L.; Teng, H.; Fang, T.; Xiao, J. Agrimonolide from Agrimonia pilosa suppresses inflammatory responses through down-regulation of COX-2/iNOS and inactivation of NF-kappaB in lipopolysaccharide-stimulated macrophages. Phytomedicine 2016, 23, 846–855.
- Aboonabi, A.; Aboonabi, A. Anthocyanins reduce inflammation and improve glucose and lipid metabolism associated with inhibiting nuclear factor-kappaB activation and increasing PPAR-gamma gene expression in metabolic syndrome subjects. Free Radic. Biol. Med. 2020, 150, 30–39.
- Duarte, L.J.; Chaves, V.C.; Nascimento, M.; Calvete, E.; Li, M.; Ciraolo, E.; Ghigo, A.; Hirsch, E.; Simoes, C.M.O.; Reginatto, F.H.; et al. Molecular mechanism of action of Pelargonidin-3-O-glucoside, the main anthocyanin responsible for the anti-inflammatory effect of strawberry fruits. Food Chem. 2018, 247, 56–65.
- Karunarathne, W.; Lee, K.T.; Choi, Y.H.; Jin, C.Y.; Kim, G.Y. Anthocyanins isolated from Hibiscus syriacus L. attenuate lipopolysaccharide-induced inflammation and endotoxic shock by inhibiting the TLR4/MD2-mediated NF-kappaB signaling pathway. Phytomedicine 2020, 76, 153237.
- Bueno, J.M.; Sáez-Plaza, P.; Ramos-Escudero, F.; Jiménez, A.M.; Fett, R.; Asuero, A.G. Analysis and Antioxidant Capacity of Anthocyanin Pigments. Part II: Chemical Structure, Color, and Intake of Anthocyanins. Crit. Rev. Anal. Chem. 2012, 42, 126–151.
- Fukumoto, L.R.; Mazza, G. Assessing antioxidant and prooxidant activities of phenolic compounds. J. Agric. Food Chem. 2000, 48, 3597–3604.
- Harakotr, B.; Suriharn, B.; Tangwongchai, R.; Scott, M.P.; Lertrat, K. Anthocyanins and antioxidant activity in coloured waxy corn at different maturation stages. J. Funct. Foods 2014, 9, 109–118.
- Matera, R.; Gabbanini, S.; Berretti, S.; Amorati, R.; De Nicola, G.R.; Iori, R.; Valgimigli, L. Acylated anthocyanins from sprouts of Raphanus sativus cv. Sango: Isolation, structure elucidation and antioxidant activity. Food Chem. 2015, 166, 397–406.
- Coklar, H.; Akbulut, M. Anthocyanins and phenolic compounds of Mahonia aquifolium berries and their contributions to antioxidant activity. J. Funct. Foods 2017, 35, 166–174.
- Lu, X.; Zhou, Y.; Wu, T.; Hao, L. Ameliorative effect of black rice anthocyanin on senescent mice induced by D-galactose. Food Funct. 2014, 5, 2892–2897.
- Huang, W.; Zhu, Y.; Li, C.; Sui, Z.; Min, W. Effect of Blueberry Anthocyanins Malvidin and Glycosides on the Antioxidant Properties in Endothelial Cells. Oxid. Med. Cell. Longev. 2016, 2016, 1591803.
- Daveri, E.; Cremonini, E.; Mastaloudis, A.; Hester, S.N.; Wood, S.M.; Waterhouse, A.L.; Anderson, M.; Fraga, C.G.; Oteiza, P.I. Cyanidin and delphinidin modulate inflammation and altered redox signaling improving insulin resistance in high fat-fed mice. Redox Biol. 2018, 18, 16–24.
- Jiang, X.; Guo, H.; Shen, T.; Tang, X.; Yang, Y.; Ling, W. Cyanidin-3-O-beta-glucoside Purified from Black Rice Protects Mice against Hepatic Fibrosis Induced by Carbon Tetrachloride via Inhibiting Hepatic Stellate Cell Activation. J. Agric. Food Chem. 2015, 63, 6221–6230.
- Arjinajarn, P.; Chueakula, N.; Pongchaidecha, A.; Jaikumkao, K.; Chatsudthipong, V.; Mahatheeranont, S.; Norkaew, O.; Chattipakorn, N.; Lungkaphin, A. Anthocyanin-rich Riceberry bran extract attenuates gentamicin-induced hepatotoxicity by reducing oxidative stress, inflammation and apoptosis in rats. Biomed. Pharmacother. 2017, 92, 412–420.
- Zhang, Z.F.; Lu, J.; Zheng, Y.L.; Wu, D.M.; Hu, B.; Shan, Q.; Cheng, W.; Li, M.Q.; Sun, Y.Y. Purple sweet potato color attenuates hepatic insulin resistance via blocking oxidative stress and endoplasmic reticulum stress in high-fat-diet-treated mice. J. Nutr. Biochem. 2013, 24, 1008–1018.
- Cai, Z.; Song, L.; Qian, B.; Xu, W.; Ren, J.; Jing, P.; Oey, I. Understanding the effect of anthocyanins extracted from purple sweet potatoes on alcohol-induced liver injury in mice. Food Chem. 2018, 245, 463–470.
- Gowd, V.; Jia, Z.Q.; Chen, W. Anthocyanins as promising molecules and dietary bioactive components against diabetes—A review of recent advances. Trends Food Sci. Technol. 2017, 68, 1–13.
- Hong, S.H.; Heo, J.I.; Kim, J.H.; Kwon, S.O.; Yeo, K.M.; Bakowska-Barczak, A.M.; Kolodziejczyk, P.; Ryu, O.H.; Choi, M.K.; Kang, Y.H.; et al. Antidiabetic and Beta Cell-Protection Activities of Purple Corn Anthocyanins. Biomol. Ther. 2013, 21, 284–289.
- Sarikaya, I.; Schierz, J.-H.; Sarikaya, A. Liver: Glucose metabolism and 18F-fluorodeoxyglucose PET findings in normal parenchyma and diseases. Am. J. Nucl. Med. Mol. Imaging 2021, 11, 233–249.
- Choi, K.H.; Lee, H.A.; Park, M.H.; Han, J.S. Mulberry (Morus alba L.) Fruit Extract Containing Anthocyanins Improves Glycemic Control and Insulin Sensitivity via Activation of AMP-Activated Protein Kinase in Diabetic C57BL/Ksj-db/db Mice. J. Med. Food 2016, 19, 737–745.
- Huang, Y.; Hu, C.; Ye, H.; Luo, R.; Fu, X.; Li, X.; Huang, J.; Chen, W.; Zheng, Y. Inflamm-Aging: A New Mechanism Affecting Premature Ovarian Insufficiency. J. Immunol. Res. 2019, 2019, 8069898.
- Zhao, J.G.; Yan, Q.Q.; Lu, L.Z.; Zhang, Y.Q. In vivo antioxidant, hypoglycemic, and anti-tumor activities of anthocyanin extracts from purple sweet potato. Nutr. Res. Pract. 2013, 7, 359–365.
- Wang, Y.; Zhang, D.; Liu, Y.; Wang, D.; Liu, J.; Ji, B. The protective effects of berry-derived anthocyanins against visible light-induced damage in human retinal pigment epithelial cells. J. Sci. Food Agric. 2015, 95, 936–944.
- Gao, J.M.; Wu, Y.T.; He, D.J.; Zhu, X.Q.; Li, H.B.; Liu, H.F.; Liu, H.L. Anti-aging effects of Ribes meyeri anthocyanins on neural stem cells and aging mice. Aging 2020, 12, 17738–17753.
- Wei, J.; Zhang, G.; Zhang, X.; Xu, D.; Gao, J.; Fan, J. Anthocyanins Delay Ageing-Related Degenerative Changes in the Liver. Plant Foods Hum. Nutr. 2017, 72, 425–431.
- Qin, S.; Sun, D.; Mu, J.; Ma, D.; Tang, R.; Zheng, Y. Purple sweet potato color improves hippocampal insulin resistance via down-regulating SOCS3 and galectin-3 in high-fat diet mice. Behav. Brain Res. 2019, 359, 370–377.
- Lee, M.; Sorn, S.R.; Park, Y.; Park, H.K. Anthocyanin Rich-Black Soybean Testa Improved Visceral Fat and Plasma Lipid Profiles in Overweight/Obese Korean Adults: A Randomized Controlled Trial. J. Med. Food 2016, 19, 995–1003.
- Farrell, N.; Norris, G.; Lee, S.G.; Chun, O.K.; Blesso, C.N. Anthocyanin-rich black elderberry extract improves markers of HDL function and reduces aortic cholesterol in hyperlipidemic mice. Food Funct. 2015, 6, 1278–1287.
- Roder, P.V.; Wu, B.; Liu, Y.; Han, W. Pancreatic regulation of glucose homeostasis. Exp. Mol. Med. 2016, 48, e219.
More
Information
Subjects:
Food Science & Technology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
10.7K
Revisions:
4 times
(View History)
Update Date:
01 Aug 2022
Table of Contents



Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No