 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Asdren Zajmi | -- | 3080 | 2022-07-13 09:04:03 | | | |
| 2 | Peter Tang | Meta information modification | 3080 | 2022-07-22 03:03:44 | | |
Video Upload Options
Elizabethkingia spp. is a ubiquitous pathogenic bacterium that has been identified as the causal agent for a variety of conditions such as meningitis, pneumonia, necrotizing fasciitis, endophthalmitis, and sepsis and is emerging as a global threat including in Southeast Asia. Elizabethkingia infections tend to be associated with high mortality rates (18.2–41%) and are mostly observed in neonates and immunocompromised patients. Difficulties in precisely identifying Elizabethkingia at the species level by traditional methods have hampered the understanding of this genus in human infections. In Southeast Asian countries, hospital outbreaks have usually been ascribed to E. meningoseptica, whereas in Singapore, E. anophelis was reported as the main Elizabethkingia spp. associated with hospital settings. Misidentification of Elizabethkingia spp. could, however, underestimate the number of cases attributed to the bacterium, as precise identification requires tools such as MALDI-TOF MS, and particularly whole-genome sequencing, which are not available in most hospital laboratories. Elizabethkingia spp. has an unusual antibiotic resistance pattern for a Gram-negative bacterium with a limited number of horizontal gene transfers, which suggests an intrinsic origin for its multidrug resistance.
1. Introduction
2. Identification
3. Antibiotic Resistance
4. Virulence Factors
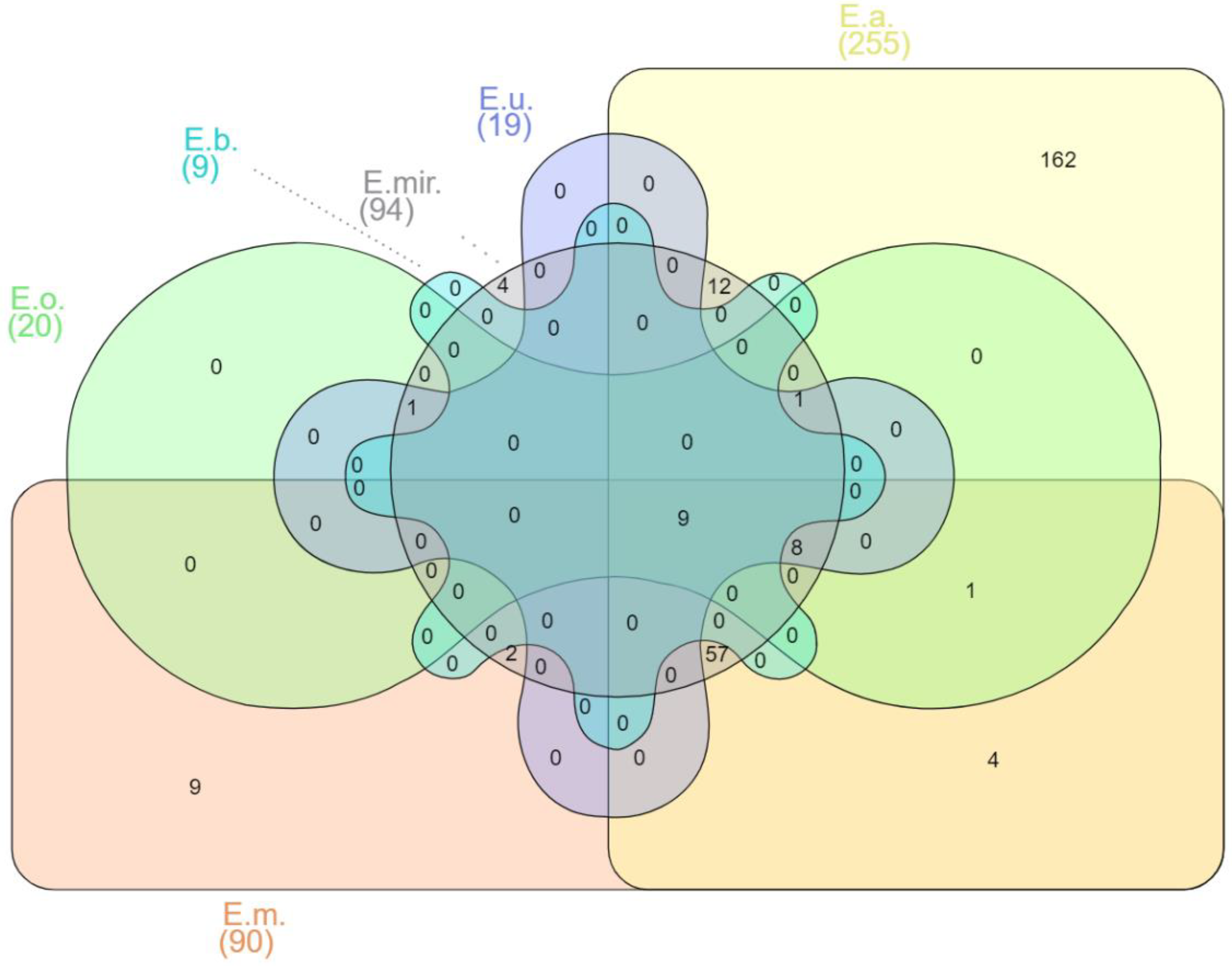
5. Sources of Isolation and Transmission
|
Source of Isolation |
Country of Origin |
Citation |
|---|---|---|
|
Blood |
Malaysia, Singapore, Thailand, Indonesia, Cambodia |
|
|
Peritoneal fluid |
Malaysia |
[77] |
|
Cerebrospinal fluid (CSF) |
Malaysia, Singapore |
|
|
Contact lens |
Malaysia |
[87] |
|
Hospital environment (aerators, sink drains and traps at ICUs, pediatric wards, surgical wards, orthopedic wards) |
Singapore |
|
|
Catheter tips |
Singapore |
[8] |
|
Respiratory specimens |
Singapore Malaysia |
|
|
Rectal swabs |
Singapore |
[91] |
|
Urine |
Malaysia |
[64] |
|
Wound swabs |
Malaysia |
[64] |
|
Nasal swabs |
Malaysia |
[64] |
|
Vitreous culture |
Singapore |
[8] |
|
Frogs (Rana catesbeiana (American bullfrogs) and Theloderma bicolor Chapa bug-eyed frogs, Warty toads (Bombina microdeladigitora), and Northern leopard frogs (Lithobates pipiens) |
Malaysia Vietnam |
|
|
Mosquitoes (Anopheles minimus, Anopheles dirus, Anopheles maculatus, Anopheles sawadwongporni, and Anopheles dravidicus) |
Thailand |
|
|
Fish (Clarias gariepinus (African sharptooth catfish) and Pangasius hypophthalmus (Tra catfish) |
Malaysia, Vietnam |
|
|
Retail sausages |
Malaysia |
[74] |
|
Gnetum gnemon (Tree) |
Malaysia |
References
- Lau, S.K.; Chow, W.-N.; Foo, C.-H.; Curreem, S.O.; Lo, G.C.-S.; Teng, J.L.; Chen, J.H.; Ng, R.H.; Wu, A.K.; Cheung, I.Y.; et al. Elizabethkingia anophelis bacteremia is associated with clinically significant infections and high mortality. Sci. Rep. 2016, 6, 26045.
- Dziuban, E.J.; Franks, J.L.; So, M.; Peacock, G.; Blaney, D.D. Elizabethkingia in children: A comprehensive review of symptomatic cases reported from 1944 to 2017. Clin. Infect. Dis. 2018, 67, 144–149.
- Opota, O.; Diene, S.M.; Bertelli, C.; Prod’hom, G.; Eckert, P.; Greub, G. Genome of the carbapenemase-producing clinical isolate Elizabethkingia miricola EM_CHUV and comparative genomics with Elizabethkingia meningoseptica and Elizabethkingia anophelis: Evidence for intrinsic multidrug resistance trait of emerging pathogens. Int. J. Antimicrob. Agents 2017, 49, 93–97.
- Swain, B.; Rout, S.; Otta, S.; Rakshit, A. Elizabethkingia meningoseptica: An unusual cause for septicaemia. JMM Case Rep. 2015, 2, e000005.
- Lee, C.-H.; Lin, W.-C.; Chia, J.-H.; Su, L.-H.; Chien, C.-C.; Mao, A.-H.; Liu, J.-W. Community-acquired osteomyelitis caused by Chryseobacterium meningosepticum: Case report and literature review. Diagn. Microbiol. Infect. Dis. 2008, 60, 89–93.
- Gupta, P.; Zaman, K.; Mohan, B.; Taneja, N. Elizabethkingia miricola: A rare non-fermenter causing urinary tract infection. World J. Clin. Cases 2017, 5, 187.
- Raghavan, S.; Thomas, B.; Shastry, B. Elizabethkingia meningoseptica: Emerging multidrug resistance in a nosocomial pathogen. Case Rep. 2017, 2017, bcr-2017-221076.
- Young, S.M.; Lingam, G.; Tambyah, P.A. Elizabethkingia meningoseptica Engodenous Endophthalmitis—A Case Report. Antimicrob. Resist. Infect. Control 2014, 3, 35.
- Yang, J.; Xue, W.; Yu, X. Elizabethkingia meningosepticum endocarditis: A rare case and special therapy. Anatol. J. Cardiol. 2015, 15, 427.
- Chi, S.; Fekete, T. Epididymo-orchitis. In Clinical Infectious Disease, 2nd ed.; Schlossberg, D., Ed.; Cambridge University Press: Cambridge, UK, 2015; pp. 401–405.
- Gonzalez, C.; Coolen-Allou, N.; Allyn, J.; Esteve, J.; Belmonte, O.; Allou, N. Severe sepsis and pulmonary abscess with bacteremia due to Elizabethkingia miricola. Med. Mal. Infect. 2015, 46, 49–51.
- Lee, C.-C.; Chen, P.-L.; Wang, L.-R.; Lee, H.-C.; Chang, C.-M.; Lee, N.-Y.; Wu, C.-J.; Shih, H.-I.; Ko, W.-C. Fatal case of community-acquired bacteremia and necrotizing fasciitis caused by Chryseobacterium meningosepticum: Case report and review of the literature. J. Clin. Microbiol. 2006, 44, 1181–1183.
- Taufiq Kadafi, K.; Yuliarto, S.; Aji Cahyono, H.; Ratridewi, I.; Khalasha, T. Cerebral Salt Wasting Due to Bacteremia Caused by Elizabethkingia meningoseptica: A Case Report. Arch. Pediatr. Infect. Dis. 2020, 8, e44832.
- Kenna, D.T.; Fuller, A.; Martin, K.; Perry, C.; Pike, R.; Burns, P.J.; Narayan, O.; Wilkinson, S.; Hill, R.; Woodford, N.; et al. rpoB gene sequencing highlights the prevalence of an E. miricola cluster over other Elizabethkingia species among UK cystic fibrosis patients. Diagn. Microbiol. Infect. Dis. 2018, 90, 109–114.
- Amir, A.; IC Sam, J.; Nawi, S. Elizabethkingia meningoseptica neonatal meningitis in a premature infant. Asian J. Med. Biomed. 2018, 2 (Suppl. 1), 22.
- Seong, H.; Kim, J.H.; Kim, J.H.; Lee, W.J.; Ahn, J.Y.; Ku, N.S.; Choi, J.Y.; Yeom, J.S.; Song, Y.G.; Jeong, S.J. Risk factors for mortality in patients with elizabethkingia infection and the clinical impact of the antimicrobial susceptibility patterns of elizabethkingia species. J. Clin. Med. 2020, 9, 1431.
- Nori, P.; Cowman, K.; Chen, V.; Bartash, R.; Szymczak, W.; Madaline, T.; Katiyar, C.P.; Jain, R.; Aldrich, M.; Weston, G.; et al. Bacterial and fungal coinfections in COVID-19 patients hospitalized during the New York City pandemic surge. Infect. Control Hosp. Epidemiol. 2021, 42, 84–88.
- Hayek, S.S.; Abd, T.T.; Cribbs, S.K.; Anderson, A.M.; Melendez, A.; Kobayashi, M.; Polito, C.; Wang, Y.F.W. Rare Elizabethkingia meningosepticum meningitis case in an immunocompetent adult. Emerg. Microbes Infect. 2013, 2, 1–4.
- Sebastiampillai, B.S.; Luke, N.V.; Silva, S.; De Silva, S.T.; Premaratna, R. Septicaemia caused by Elizabethkingia-sp in a ‘healthy’Sri Lankan man. Trop. Dr. 2018, 48, 62–63.
- Yang, C.; Liu, Z.; Yu, S.; Ye, K.; Li, X.; Shen, D. Comparison of three species of Elizabethkingia genus by whole-genome sequence analysis. FEMS Microbiol. Lett. 2021, 368, fnab018.
- King, E.O. Studies on a group of previously unclassified bacteria associated with meningitis in infants. Am. J. Clin. Pathol. 1959, 31, 241–247.
- Buttiaux, R.; Vandepitte, J. Flavobacterium in Epidemic Meningitis of New-Born Infants. Ann. Inst. Pasteur 1960, 98, 398–404.
- Chan, J.; Chong, C.; Thoon, K.; Tee, N.; Maiwald, M.; Lam, J.; Bhattacharya, R.; Chandran, S.; Yung, C.; Tan, N. Invasive paediatric Elizabethkingia meningoseptica infections are best treated with a combination of piperacillin/tazobactam and trimethoprim/sulfamethoxazole or fluoroquinolone. J. Med. Microbiol. 2019, 68, 1167–1172.
- Saetiew, N.; Nilkate, S.; Suankratay, C. Elizabethkingia meningoseptica Infection: The First Case Series in Thailand. Presented at the 26th European Congress of Clinical Microbiology and Infectious Diseases, Bangkok, Thailand, 9–12 April 2016.
- Agustini, N.M.A.; Wati, D.K.; Suparyatha, I.; Hartawan, I.N.B.; Utama, I.M.G.D.L.; Budayanti, N.N.S.; Tunas, I.K. The relationship between bacterial types and antibiotic resistance with the clinical outcomes of sepsis patients in Pediatric Intensive Care Unit at Sanglah Hospital Denpasar, Bali-Indonesia. Indones. J. Biomed. Sci. 2018, 12, 13–18.
- Reed, T.A.; Watson, G.; Kheng, C.; Tan, P.; Roberts, T.; Ling, C.L.; Miliya, T.; Turner, P. Elizabethkingia anophelis Infection in Infants, Cambodia, 2012–2018. Emerg. Infect. Dis. 2020, 26, 320.
- Frederiksen, W.; Ursing, J. Proposed new bacterial taxa and proposed changes of bacterial names published during 1994 and considered to be of interest to medical or veterinary bacteriology. APMIS 1995, 103, 651–654.
- Kim, K.; Kim, M.; Lim, J.; Park, H.; Lee, S. Transfer of Chryseobacterium meningosepticum and Chryseobacterium miricola to Elizabethkingia gen. nov. as Elizabethkingia meningoseptica comb. nov. and Elizabethkingia miricola comb. nov. Int. J. Syst. Evol. Microbiol. 2005, 55, 1287–1293.
- Nicholson, A.C.; Gulvik, C.A.; Whitney, A.M.; Humrighouse, B.W.; Graziano, J.; Emery, B.; Bell, M.; Loparev, V.; Juieng, P.; Gartin, J.; et al. Revisiting the taxonomy of the genus Elizabethkingia using whole-genome sequencing, optical mapping, and MALDI-TOF, along with proposal of three novel Elizabethkingia species: Elizabethkingia bruuniana sp. nov., Elizabethkingia ursingii sp. nov., and Elizabethkingia occulta sp. nov. Antonie Van Leeuwenhoek 2018, 111, 55–72.
- Hwang, J.-H.; Kim, J.; Kim, J.-H.; Mo, S. Elizabethkingia argenteiflava sp. nov., isolated from the pod of soybean, Glycine max. Int. J. Syst. Evol. Microbiol. 2021, 71, 004767.
- Hem, S.; Jarocki, V.M.; Baker, D.J.; Charles, I.G.; Drigo, B.; Aucote, S.; Donner, E.; Burnard, D.; Bauer, M.J.; Harris, P.N.; et al. Genomic analysis of Elizabethkingia species from aquatic environments: Evidence for potential clinical transmission. Curr. Res. Microb. Sci. 2022, 3, 100083.
- Kukutla, P.; Lindberg, B.G.; Pei, D.; Rayl, M.; Yu, W.; Steritz, M.; Faye, I.; Xu, J. Insights from the genome annotation of Elizabethkingia anophelis from the malaria vector Anopheles gambiae. PLoS ONE 2014, 9, e97715.
- Lin, J.-N.; Lai, C.-H.; Yang, C.-H.; Huang, Y.-H.; Lin, H.-F.; Lin, H.-H. Comparison of four automated microbiology systems with 16S rRNA gene sequencing for identification of Chryseobacterium and Elizabethkingia species. Sci. Rep. 2017, 7, 13824.
- Ekcharoenkul, K.; Ngamskulrungroj, P.; Joyjamras, K.; Leelaporn, A.; Harun, A.; Kiratisin, P. Identification of Uncommon Pathogenic Bacteria by MALDI-TOF Mass Spectrometry Using a Custom Library of Siriraj Hospital. Siriraj Med. J. 2018, 70, 127–130.
- Han, M.-S.; Kim, H.; Lee, Y.; Kim, M.; Ku, N.S.; Choi, J.Y.; Yong, D.; Jeong, S.H.; Lee, K.; Chong, Y. Relative prevalence and antimicrobial susceptibility of clinical isolates of Elizabethkingia species based on 16S rRNA gene sequencing. J. Clin. Microbiol. 2017, 55, 274–280.
- Burnard, D.; Gore, L.; Henderson, A.; Ranasinghe, A.; Bergh, H.; Cottrell, K.; Sarovich, D.S.; Price, E.P.; Paterson, D.L.; Harris, P.N. Comparative Genomics and Antimicrobial Resistance Profiling of Elizabethkingia Isolates Reveal Nosocomial Transmission and In Vitro Susceptibility to Fluoroquinolones, Tetracyclines, and Trimethoprim-Sulfamethoxazole. J. Clin. Microbiol. 2020, 58, e00730-20.
- González, L.J.; Vila, A.J. Carbapenem resistance in Elizabethkingia meningoseptica is mediated by metallo-β-lactamase BlaB. Antimicrob. Agents Chemother. 2012, 56, 1686–1692.
- Teo, J.; Tan, S.Y.-Y.; Liu, Y.; Tay, M.; Ding, Y.; Li, Y.; Kjelleberg, S.; Givskov, M.; Lin, R.T.P.; Yang, L. Comparative Genomic Analysis of Malaria Mosquito Vector-Associated Novel Pathogen Elizabethkingia anophelis. Genome Biol. Evol. 2014, 6, 1158–1165.
- Breurec, S.; Criscuolo, A.; Diancourt, L.; Rendueles, O.; Vandenbogaert, M.; Passet, V.; Caro, V.; Rocha, E.P.; Touchon, M.; Brisse, S. Genomic epidemiology and global diversity of the emerging bacterial pathogen Elizabethkingia anophelis. Sci. Rep. 2016, 6, 30379.
- Hu, R.; Zhang, Q.; Gu, Z. Molecular diversity of chromosomal metallo-β-lactamase genes in Elizabethkingia genus. Int. J. Antimicrob. Agents 2020, 56, 105978.
- Hu, R.; Zhang, Q.; Gu, Z. Whole-genome analysis of the potentially zoonotic Elizabethkingia miricola FL160902 with two new chromosomal MBL gene variants. J. Antimicrob. Chemother. 2020, 75, 526–530.
- Perrin, A.; Larsonneur, E.; Nicholson, A.C.; Edwards, D.J.; Gundlach, K.M.; Whitney, A.M.; Gulvik, C.A.; Bell, M.E.; Rendueles, O.; Cury, J.; et al. Evolutionary dynamics and genomic features of the Elizabethkingia anophelis 2015 to 2016 Wisconsin outbreak strain. Nat. Commun. 2017, 8, 15483.
- Lin, J.-N.; Lai, C.-H.; Yang, C.-H.; Huang, Y.-H. Elizabethkingia infections in humans: From genomics to clinics. Microorganisms 2019, 7, 295.
- Liang, C.-Y.; Yang, C.-H.; Lai, C.-H.; Huang, Y.-H.; Lin, J.-N. Comparative Genomics of 86 Whole-Genome Sequences in the Six Species of the Elizabethkingia Genus Reveals Intraspecific and Interspecific Divergence. Sci. Rep. 2019, 9, 19167.
- Bellais, S.; Poirel, L.; Naas, T.; Girlich, D.; Nordmann, P. Genetic-Biochemical Analysis and Distribution of the Ambler Class A β-Lactamase CME-2, Responsible for Extended-Spectrum Cephalosporin Resistance in Chryseobacterium (Flavobacterium) meningosepticum. Antimicrob. Agents Chemother. 2000, 44, 1–9.
- Chang, J.; Hsueh, P.; Wu, J.; Ho, S.; Hsieh, W.; Luh, K. Antimicrobial susceptibility of flavobacteria as determined by agar dilution and disk diffusion methods. Antimicrob. Agents Chemother. 1997, 41, 1301–1306.
- Moulin, V.; Freney, J.; Hansen, W.; Philippon, A. Comportement phénotypique des Flavobacterium vis-à-vis de 39 antibiotiques. Méd. Mal. Infect. 1992, 22, 902–907.
- Kwambana-Adams, B.; Laxton, C.; Foster-Nyarko, E.; Weinstock, G.; Antonio, M. Isolation of Methicillin-resistant Staphylococcus aureus and Multidrug-resistant Elizabethkingia meningoseptica from Neonates within Minutes of Birth. Pediatric Infect. Dis. J. 2017, 36, 123–124.
- Huang, Y.; Huang, Y.; Lin, Y.; Wang, F.; Chan, Y.; Yang, T. Risk factors and outcome of levofloxacin-resistant Elizabethkingia meningoseptica bacteraemia in adult patients in Taiwan. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 1–8.
- Jean, S.-S.; Hsieh, T.-C.; Ning, Y.-Z.; Hsueh, P.-R. Role of vancomycin in the treatment of bacteraemia and meningitis caused by Elizabethkingia meningoseptica. Int. J. Antimicrob. Agents 2017, 50, 507–511.
- Lee, E.; Robinson, M.; Thong, M.; Puthucheary, S.; Ong, T.; Ng, K. Intraventricular chemotherapy in neonatal meningitis. J. Pediatr. 1977, 91, 991–995.
- Lim, V.; Halijah, M. A comparative study of the in-vitro activity of cefepime and other cephalosporins. Malays. J. Pathol. 1993, 15, 65–68.
- Chang, T.-Y.; Chen, H.-Y.; Chou, Y.-C.; Cheng, Y.-H.; Sun, J.-R. In vitro activities of imipenem, vancomycin, and rifampicin against clinical Elizabethkingia species producing BlaB and GOB metallo-beta-lactamases. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 2045–2052.
- Santona, A.; Paglietti, B.; Al-Qahtani, A.A.; Bohol, M.F.F.; Senok, A.; Deligios, M.; Rubino, S.; Al-Ahdal, M.N. Novel type of VanB2 teicoplanin-resistant hospital-associated Enterococcus faecium. Int. J. Antimicrob. Agents 2014, 44, 156–159.
- Sader, H.S.; Jones, R.N.; Pfaller, M.A. Relapse of catheter-related Flavobacterium meningosepticum bacteremia demonstrated by DNA macrorestriction analysis. Clin. Infect. Dis. 1995, 21, 997–1000.
- Ozkalay, N.; Anil, M.; Agus, N.; Helvaci, M.; Sirti, S. Community-acquired meningitis and sepsis caused by Chryseobacterium meningosepticum in a patient diagnosed with thalassemia major. J. Clin. Microbiol. 2006, 44, 3037–3039.
- Chen, S.; Soehnlen, M.; Blom, J.; Terrapon, N.; Henrissat, B.; Walker, E.D. Comparative genomic analyses reveal diverse virulence factors and antimicrobial resistance mechanisms in clinical Elizabethkingia meningoseptica strains. PLoS ONE 2019, 14, e0222648.
- Lin, J.-N.; Lai, C.-H.; Yang, C.-H.; Huang, Y.-H.; Lin, H.-H. Genomic features, phylogenetic relationships, and comparative genomics of Elizabethkingia anophelis strain EM361-97 isolated in Taiwan. Sci. Rep. 2017, 7, 14317.
- Heberle, H.; Meirelles, G.V.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinform. 2015, 16, 169.
- Li, Y.; Liu, Y.; Chew, S.C.; Tay, M.; Salido, M.M.S.; Teo, J.; Lauro, F.M.; Givskov, M.; Yang, L. Complete genome sequence and transcriptomic analysis of the novel pathogen Elizabethkingia anophelis in response to oxidative stress. Genome Biol. Evol. 2015, 7, 1676–1685.
- Lau, S.K.; Wu, A.K.; Teng, J.L.; Tse, H.; Curreem, S.O.; Tsui, S.K.; Huang, Y.; Chen, J.H.; Lee, R.A.; Yuen, K.-Y.; et al. Evidence for Elizabethkingia anophelis transmission from mother to infant, Hong Kong. Emerg. Infect. Dis. 2015, 21, 232.
- Jacobs, A.; Chenia, H.Y. Biofilm formation and adherence characteristics of an Elizabethkingia meningoseptica isolate from Oreochromis mossambicus. Ann. Clin. Microbiol. Antimicrob. 2011, 10, 16.
- Alav, I.; Sutton, J.M.; Rahman, K.M. Role of bacterial efflux pumps in biofilm formation. J. Antimicrob. Chemother. 2018, 73, 2003–2020.
- Puah, S.M.; Fong, S.P.; Kee, B.P.; Puthucheary, S.; Chua, K.H. Molecular identification and biofilm-forming ability of Elizabethkingia species. Microb. Pathog. 2022, 162, 105345.
- Dunn, M.; Ramirez-Trujillo, J.; Hernández-Lucas, I. Major roles of isocitrate lyase and malate synthase in bacterial and fungal pathogenesis. Microbiology 2009, 155, 3166–3175.
- McKinney, J.D.; Zu Bentrup, K.H.; Muñoz-Elías, E.J.; Miczak, A.; Chen, B.; Chan, W.-T.; Swenson, D.; Sacchettini, J.C.; Jacobs, W.R.; Russell, D.G. Persistence of Mycobacterium tuberculosis in macrophages and mice requires the glyoxylate shunt enzyme isocitrate lyase. Nature 2000, 406, 735–738.
- Roberts, M.F.; Khan, H.M.; Goldstein, R.; Reuter, N.; Gershenson, A. Search and subvert: Minimalist bacterial phosphatidylinositol-specific phospholipase C enzymes. Chem. Rev. 2018, 118, 8435–8473.
- Monturiol-Gross, L.; Villalta-Romero, F.; Flores-Díaz, M.; Alape-Girón, A. Bacterial phospholipases C with dual activity: Phosphatidylcholinesterase and sphingomyelinase. FEBS Open Bio 2021, 11, 3262–3275.
- Vicente, C.S.; Nascimento, F.X.; Ikuyo, Y.; Cock, P.J.; Mota, M.; Hasegawa, K. The genome and genetics of a high oxidative stress tolerant Serratia sp. LCN16 isolated from the plant parasitic nematode Bursaphelenchus xylophilus. BMC Genom. 2016, 17, 301.
- Hassett, D.J.; Alsabbagh, E.; Parvatiyar, K.; Howell, M.L.; Wilmott, R.W.; Ochsner, U.A. A protease-resistant catalase, KatA, released upon cell lysis during stationary phase is essential for aerobic survival of a Pseudomonas aeruginosa oxyR mutant at low cell densities. J. Bacteriol. 2000, 182, 4557–4563.
- Wang, M.; Gao, H.; Lin, N.; Zhang, Y.; Huang, N.; Walker, E.D.; Ming, D.; Chen, S.; Hu, S. The antibiotic resistance and pathogenicity of a multidrug-resistant Elizabethkingia anophelis isolate. Microbiologyopen 2019, 8, e804.
- Yang, C.; Liu, Z.; Yu, S.; Ye, K.; Li, X.; Shen, D. Comparison of Whole-Genome Sequences for Three Species of the Elizabethkingia Genus. 2020. Available online: https://www.researchsquare.com/article/rs-61004/v1 (accessed on 21 April 2022).
- Laith, A.; Mazlan, A.; Ambak, M.; Jabar, A.; Najiah, M. Isolation and Identification of Elizabethkingia meningoseptica from Diseased African Catfish Clarias gariepinus. J. Microbiol. Biotechnol. Food Sci. 2017, 6, 1070–1076.
- Tew, L.-S.; She, L.-Y.; Chew, C.-H. Isolation, Antimicrobial Susceptibility Profile and Detection of Sul1, blaTEM, and blaSHV in Amoxicillin-Clavulanate-Resistant Bacteria Isolated From Retail Sausages in Kampar, Malaysia. Jundishapur J. Microbiol. 2016, 9, e37897.
- Chen, C.; Chen, Y.; Wang, F. Risk factors of healthcare-associated Elizabethkingia meningoseptica infections in Taiwan medical center. Int. J. Antimicrob. Agents 2017, 50, S141.
- Khan, I.; Lall, M.; Sen, S.; Ninawe, S.; Chandola, P. Multiresistant Elizabethkingia meningoseptica infections in tertiary care. Med. J. Armed Forces India 2015, 71, 282.
- Thong, M.; Puthucheary, S.; Lee, E. Flavobacterium meningosepticum infection: An epidemiological study in a newborn nursery. J. Clin. Pathol. 1981, 34, 429–433.
- Chew, K.L.; Cheng, B.; Lin, R.T.; Teo, J.W. Elizabethkingia anophelis is the dominant Elizabethkingia species found in blood cultures in Singapore. J. Clin. Microbiol. 2018, 56, e01445-17.
- Yung, C.-F.; Maiwald, M.; Loo, L.H.; Soong, H.Y.; Tan, C.B.; Lim, P.K.; Li, L.; Tan, N.W.; Chong, C.-Y.; Tee, N.; et al. Elizabethkingia anophelis and association with tap water and handwashing, Singapore. Emerg. Infect. Dis. 2018, 24, 1730.
- Loo, L.W.; Liew, Y.X.; Choong, H.L.L.; Tan, A.L.; Chlebicki, P. Microbiology and audit of vascular access-associated bloodstream infections in multi-ethnic Asian hemodialysis patients in a tertiary hospital. Infect. Dis. 2015, 47, 225–230.
- Sooklin, L.; Anand, A.J.; Rajadurai, V.S.; Chandran, S. Management of large congenital chylous ascites in a preterm infant: Fetal and neonatal interventions. BMJ Case Rep. CP 2020, 13, e235849.
- Liestiadi, D.E.F.; Azlin, E.; Nafianti, S. A hematologic scoring system and C-reactive protein compared to blood cultures for diagnosing bacterial neonatal sepsis. Paediatr. Indones 2017, 57, 71.
- Lee, E.; Robinson, M.; Thong, M.; Puthucheary, S. Rifamycin in Neonatal Flavobacteria meningitis. Arch. Dis. Child. 1976, 51, 209–213.
- Raimondi, A.; Moosdeen, F.; Williams, J. Antibiotic resistance pattern of Flavobacterium meningosepticum. Eur. J. Clin. Microbiol. Infect. Dis. 1986, 5, 461–463.
- Zakaria, Z.; Idris, B. Intraoperative Cerebrospinal Fluid Sample from First Ventriculoperitoneal Shunt Operation: Is it Indicated? Malays. J. Med. Sci. MJMS 2013, 20, 102.
- Wan Hassan, W.M.N.; Paramasivam, R.P.; Kandasamy, R.; Hassan, M.H.; Zaini, R.H.M. An uncommon Elizabethkingia meningoseptica septicemia in hemorrhagic stroke with septic shock patient during prolonged neuro-intensive care management. Anaesth. Pain Intensive Care 2017, 21, 268–271.
- Ali, N.A.M.; Reddy, S.C. Bilateral simultaneous infectious keratitis secondary to contact lens wear: An unusual case report with rare organisms. Eye Contact Lens 2007, 33, 338–340.
- Phoon, H.Y.; Hussin, H.; Hussain, B.M.; Lim, S.Y.; Woon, J.J.; Er, Y.X.; Thong, K.L. Distribution, genetic diversity and antibiotic resistance of clinically important bacteria from the environment of a tertiary hospital. J. Glob. Antimicrob. Resist. 2018, 14, 132–140.
- Haller, L.; Chen, H.; Ng, C.; Le, T.H.; Koh, T.H.; Barkham, T.; Sobsey, M.; Gin, K.Y.-H. Occurrence and characteristics of extended-spectrum β-lactamase-and carbapenemase-producing bacteria from hospital effluents in Singapore. Sci. Total Environ. 2018, 615, 1119–1125.
- Balm, M.; Salmon, S.; Jureen, R.; Teo, C.; Mahdi, R.; Seetoh, T.; Teo, J.; Lin, R.; Fisher, D. Bad design, bad practices, bad bugs: Frustrations in controlling an outbreak of Elizabethkingia meningoseptica in intensive care units. J. Hosp. Infect. 2013, 85, 134–140.
- Venkatachalam, I.; Teo, J.; Balm, M.N.; Fisher, D.A.; Jureen, R.; Lin, R.T. Klebsiella pneumoniae carbapenemase-producing enterobacteria in hospital, Singapore. Emerg. Infect. Dis. 2012, 18, 1381.
- Trimpert, J.; Eichhorn, I.; Vladimirova, D.; Haake, A.; Schink, A.K.; Klopfleisch, R.; Lübke-Becker, A. Elizabethkingia miricola infection in multiple anuran species. Transbound. Emerg. Dis. 2020, 68, 931–940.
- Zainuri, N.; Ransangan, J.; Lal, T.; Jintoni, B.; Chung, V. Identification of Elizabethkingia meningoseptica from American bullfrog (Rana catesbeiana) farmed in Sabah, Malaysia using PCR method and future management of outbreak. Malays. J. Microbiol. 2013, 9, 13–23.
- Tee, L.; Najiah, M. Antibiogram and heavy metal tolerance of bullfrog bacteria in Malaysia. Open Vet. J. 2011, 1, 39–45.
- Tainchum, K.; Dupont, C.; Chareonviriyaphap, T.; Jumas-Bilak, E.; Bangs, M.J.; Manguin, S. Bacterial microbiome in wild-caught Anopheles mosquitoes in western Thailand. Front. Microbiol. 2020, 11, 965.
- Surat, W.; Mhuantong, W.; Sangsrakru, D.; Chareonviriyaphap, T.; Arunyawat, U.; Kubera, A.; Sittivicharpinyo, T.; Siripan, O.; Pootakham, W. Gut Bacterial Diversity in Plasmodium-infected and Plasmodium-uninfected Anopheles minimus. Chiang Mai J. Sci. 2016, 43, 427–440.
- Karl, H.; Lehmann, I.; Rehbein, H.; Schubring, R. Composition and quality attributes of conventionally and organically farmed Pangasius fillets (Pangasius hypophthalmus) on the German market. Int. J. Food Sci. Technol. 2010, 45, 56–66.
- Thi, A.N.T.; Noseda, B.; Samapundo, S.; Nguyen, B.L.; Broekaert, K.; Rasschaert, G.; Heyndrickx, M.; Devlieghere, F. Microbial ecology of Vietnamese Tra fish (Pangasius hypophthalmus) fillets during processing. Int. J. Food Microbiol. 2013, 167, 144–152.
- Kim, M.; Singh, D.; Lai-Hoe, A.; Go, R.; Rahim, R.A.; Ainuddin, A.; Chun, J.; Adams, J.M. Distinctive phyllosphere bacterial communities in tropical trees. Microb. Ecol. 2012, 63, 674–681.
- Oh, Y.M.; Kim, M.; Lee-Cruz, L.; Lai-Hoe, A.; Go, R.; Ainuddin, N.; Rahim, R.A.; Shukor, N.; Adams, J.M. Distinctive bacterial communities in the rhizoplane of four tropical tree species. Microb. Ecol. 2012, 64, 1018–1027.
- Kämpfer, P.; Matthews, H.; Glaeser, S.P.; Martin, K.; Lodders, N.; Faye, I. Elizabethkingia anophelis sp. nov., isolated from the midgut of the mosquito Anopheles gambiae. Int. J. Syst. Evol. Microbiol. 2011, 61, 2670–2675.
- Chen, S.; Johnson, B.K.; Yu, T.; Nelson, B.N.; Walker, E.D. Elizabethkingia anophelis: Physiologic and transcriptomic responses to iron stress. Front. Microbiol. 2020, 11, 804.
- Onyango, M.; Payne, A.; Stout, J.; Dieme, C.; Kuo, L.; Kramer, L.; Ciota, A. Potential for transmission of Elizabethkingia anophelis by Aedes albopictus and the role of microbial interactions in Zika virus competence. bioRxiv 2020, 702464.
- Akhouayri, I.; Habtewold, T.; Christophides, G. Melanotic pathology and vertical transmission of the gut commensal Elizabethkingia meningoseptica in the major malaria vector Anopheles gambiae. PLoS ONE 2013, 8, e77619.
- Rani, A.; Sharma, A.; Rajagopal, R.; Adak, T.; Bhatnagar, R.K. Bacterial diversity analysis of larvae and adult midgut microflora using culture-dependent and culture-independent methods in lab-reared and field-collected Anopheles stephensi-an Asian malarial vector. BMC Microbiol. 2009, 9, 96.
- Ngwa, C.; Glöckner, V.; Abdelmohsen, U.R.; Scheuermayer, M.; Fischer, R.; Hentschel, U.; Pradel, G. 16S rRNA gene-based identification of Elizabethkingia meningoseptica (Flavobacteriales: Flavobacteriaceae) as a dominant midgut bacterium of the Asian malaria vector Anopheles stephensi (Dipteria: Culicidae) with antimicrobial activities. J. Med. Entomol. 2013, 50, 404–414.
- Lindh, J.; Borg-Karlson, A.; Faye, I. Transstadial and horizontal transfer of bacteria within a colony of Anopheles gambiae (Diptera: Culicidae) and oviposition response to bacteria-containing water. Acta Trop. 2008, 107, 242–250.
- Wang, Y.; Gilbreath, T.M., III; Kukutla, P.; Yan, G.; Xu, J. Dynamic gut microbiome across life history of the malaria mosquito Anopheles gambiae in Kenya. PLoS ONE 2011, 6, e24767.
- Boissière, A.; Tchioffo, M.; Bachar, D.; Abate, L.; Marie, A.; Nsango, S.; Shahbazkia, H.; Awono-Ambene, P.; Levashina, E.; Christen, R.; et al. Midgut microbiota of the malaria mosquito vector Anopheles gambiae and interactions with Plasmodium falciparum infection. PLoS Pathog. 2012, 8, e1002742.
- Wang, S.; Ghosh, A.K.; Bongio, N.; Stebbings, K.A.; Lampe, D.J.; Jacobs-Lorena, M. Fighting malaria with engineered symbiotic bacteria from vector mosquitoes. Proc. Natl. Acad. Sci. USA 2012, 109, 12734–12739.
- Dong, Y.; Manfredini, F.; Dimopoulos, G. Implication of the mosquito midgut microbiota in the defense against malaria parasites. PLoS Pathog. 2009, 5, e1000423.


