Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Zbigniew Wyżewski | -- | 1638 | 2022-07-17 21:44:54 | | | |
| 2 | Catherine Yang | Meta information modification | 1638 | 2022-07-18 03:34:42 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Wyżewski, Z.; Mielcarska, M.B.; Gregorczyk-Zboroch, K.P.; Myszka, A. EBV-Associated Diseases. Encyclopedia. Available online: https://encyclopedia.pub/entry/25211 (accessed on 25 July 2026).
Wyżewski Z, Mielcarska MB, Gregorczyk-Zboroch KP, Myszka A. EBV-Associated Diseases. Encyclopedia. Available at: https://encyclopedia.pub/entry/25211. Accessed July 25, 2026.
Wyżewski, Zbigniew, Matylda Barbara Mielcarska, Karolina Paulina Gregorczyk-Zboroch, Anna Myszka. "EBV-Associated Diseases" Encyclopedia, https://encyclopedia.pub/entry/25211 (accessed July 25, 2026).
Wyżewski, Z., Mielcarska, M.B., Gregorczyk-Zboroch, K.P., & Myszka, A. (2022, July 17). EBV-Associated Diseases. In Encyclopedia. https://encyclopedia.pub/entry/25211
Wyżewski, Zbigniew, et al. "EBV-Associated Diseases." Encyclopedia. Web. 17 July, 2022.
Copy Citation
Epstein-Barr virus (EBV), the representative of the Herpesviridae family, is a pathogen extensively distributed in the human population. One of its most characteristic features is the capability to establish latent infection in the host. The infected cells serve as a sanctuary for the dormant virus, and therefore their desensitization to apoptotic stimuli is part of the viral strategy for long-term survival.
Epstein-Barr virus
apoptosis
1. Introduction
Apoptosis, a type I cell death, plays an essential role in the development of the organism and is necessary for its proper function. The aforementioned process normally happens during organ morphogenesis as a mechanism responsible for the controlled elimination of unwanted cells [1]. Apoptosis may be inscribed in scenario of natural growing and aging, but it also occurs in response to a wide range of incidental factors and conditions, e.g., high temperature [2][3], UV radiation [4][5][6] as well as an exposition to alcohol (ethanol) [7][8] or toxic metal ions [9][10].
Apoptosis is also the biological phenomenon that contributes to defense against intracellular infectious agents, both bacteria and viruses. Programmed cell death frustrates intracellular replication of pathogens, counteracting the development of disease. Therefore, bacteria and viruses have evolved molecular mechanisms to inhibit apoptosis and maintain the environment of their effective multiplication [11][12][13][14][15][16][17]. Research revealed anti-apoptotic activity of intracellular bacteria, Chlamydia trachomatis, [11] as well as of a wide range of viruses, such as vaccinia virus (VACV) [16][17], hepatitis B virus (HBV) [15][18], hepatitis C virus (HCV) [12][14], Kaposi’s sarcoma-associated herpesvirus (KSHV) [19] and Epstein–Barr virus (EBV) [13]. The last two pathogens, γ-herpesviruses, are causative agents of lytic and persistent infections that may lead to the development of several diseases including malignancies [19][20].
2. EBV and Its Pathogenicity
Epstein–Barr virus (EBV), a member of the Herpesviridae family and γ-Herpesvirinae subfamily, is a virus widely distributed in the human population. A total of 95% of people worldwide are estimated to carry EBV. The virion contains a 170–175 kb linear dsDNA genome [21] enclosed in an icosahedral capsid of 100–120 nm in diameter. The nucleocapsid is surrounded by a tegument and encircled by a lipid envelope decorated with glycoproteins. Viral DNA, comprising approximately 90 reading frames [22], is flanked by terminal direct repeats (TRs) and divided into short and long unique sequence domains by internal repeat sequences (IRs) [23]. EBV replicates in B, T, and natural killer (NK) lymphocytes as well as in epithelial cells [21][24]. The virus is characterized by the ability to establish persistent infection. The life cycle of EBV includes primary infection followed by latency and lytic replication [21][25]. The adaptations of EBV to establish latent infection are associated with the oncogenic properties of this pathogen [26].
EBV may be carried by healthy individuals without any symptoms of its presence. However, immunosuppression accompanying acquired immune deficiency syndrome (AIDS) or transplantation treatment favors EBV-driven fatal lymphoproliferative disease [21][25]. Moreover, EBV may contribute to the development of several malignancies, such as B cell lymphomas (i.e., Burkitt’s lymphoma (BL) [27][28][29][30][31], Hodgkin’s lymphoma (HL) [32][33], and diffuse large B-cell lymphoma (DLBCL) [34][35][36]), lung carcinoma (LC) [37][38][39], gastric cancer (GC) [40][41][42], nasopharyngeal carcinoma (NPC) [43][44][45], T cell lymphoma [46], T/NK cell lymphoma (NKTL) [47][48], and AIDS-related primary central nervous system lymphoma (AR-PCNSL) [49]. EBV has also been detected in the saliva of patients with systemic lupus erythematosus (SLE), and found to be associated with one of its symptoms (i.e., oral lesions) [50]. In BL development, the viral products complement with a cellular protein named c-myelocytomatosis oncogene product (c-Myc). The disease results from the deregulation of c-Myc expression following the translocation of its gene into immunoglobulin (Ig) loci. EBV synergizes with c-Myc in pathogenic activity by preventing apoptosis of the mutant cell [51].
The virus encodes proteins that promote the progression of EBV-associated diseases. Some of them, BamH1 fragment H rightward facing (BHRF1), BamH1-A reading frame-1 (BARF1), latent membrane proteins (LMP)-1 and -2A, EBV nuclear antigen (EBNA)-1, -2, -leader protein (LP), -3A and -3C, inhibits apoptosis of infected cells. Research has also revealed the anti-apoptotic role of EBV-encoded small RNAs (EBERs) and microRNAs (miRNAs) of EBV (Figure 1). Therefore, treatment strategies discussed in medical literature include therapies aimed at inducing apoptosis in infected cells [52][53][54].
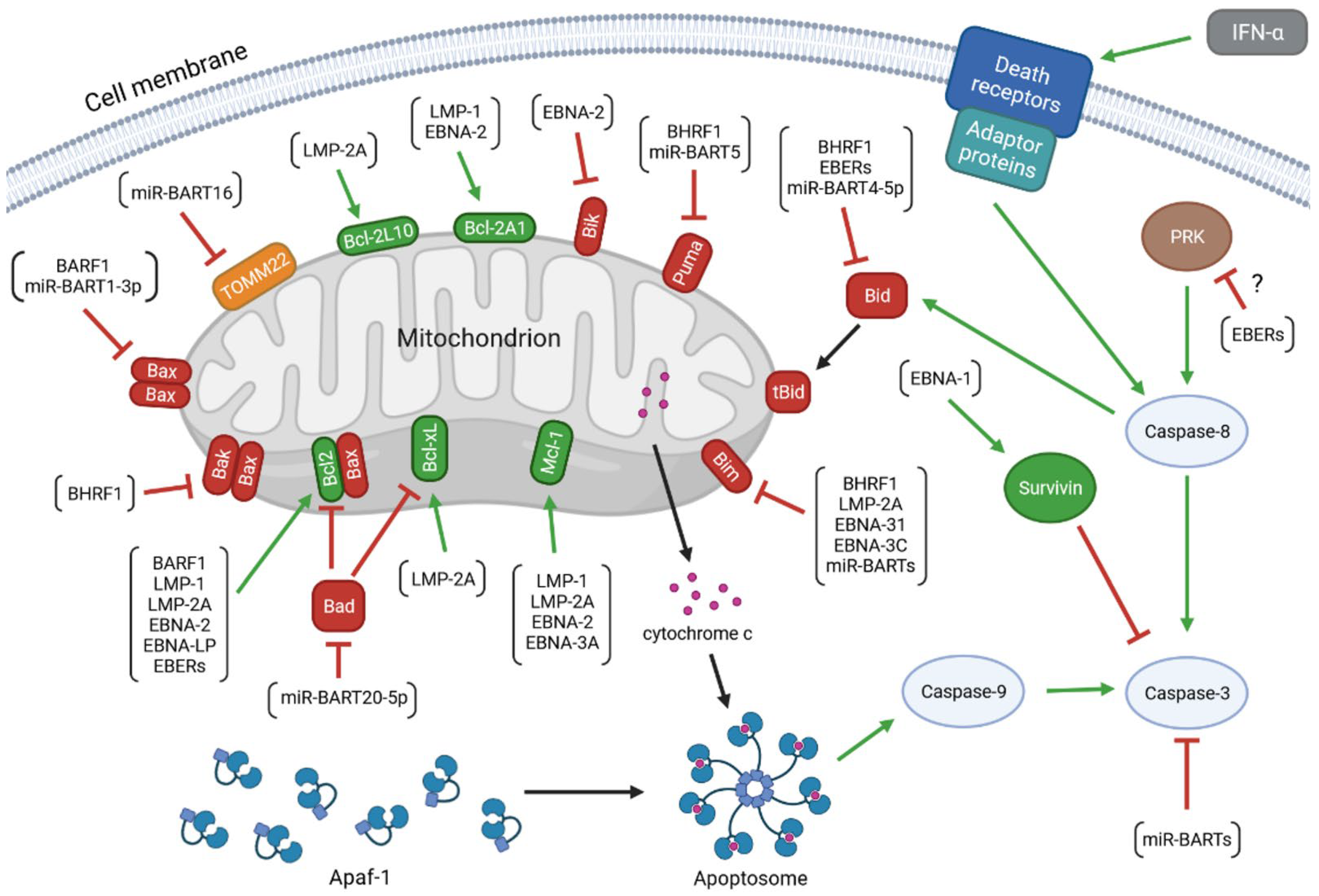
Figure 1. The anti-apoptotic activity of the EBV-encoded products. The colors of arrows represent the impact of the upstream molecules on the expression and/or activity of the target proteins (green: up-regulation; red: down-regulation). The molecular network determining the viability of the infected cell includes: (1) viral proteins: BamH1 fragment H rightward facing (BHRF1), BamH1-A reading frame-1 (BARF1), latent membrane proteins (LMP)-1 and -2A, EBV nuclear antigen (EBNA)-1, -2, -leader protein (LP), -3A and -3C; (2) viral non-coding transcripts: EBV-encoded small RNAs (EBERs), BamHI A rightward transcripts (miR-BARTs), and miR-BHRF1s; (3) anti-apoptotic members of the B-cell lymphoma 2 (Bcl-2) protein family: Bcl-2, B-cell lymphoma-extra large protein (Bcl-xL), myeloid leukemia cell differentiation protein 1 (Mcl-1), Bcl-2-like protein 10 (Bcl-2L10), and Bcl-2-related protein A1 (Bcl-2A1); (4) pro-apoptotic members of the Bcl-2 family: Bcl-2-associated X protein (Bax), Bcl-2 homologous antagonist/killer (Bak), Bcl-2-associated agonist of cell death (Bad), Bcl-2-like protein 11 (Bim), BH3-interacting domain death agonist (Bid), p53 up-regulated modulator of apoptosis (Puma), and Bcl-2 interacting killer (Bik); (5) other cellular molecules: mitochondrial import receptor subunit TOM22 homolog (TOMM22), apoptotic protease activating factor 1 (Apaf-1), cytochrome c, caspases(-8, -9, and -3), survivin, interferon (IFN)-α, death receptors, adaptor proteins, and dsRNA-dependent protein kinase (PKR).
EBV infection may lead to the host cell transformation, a process associated with changes in the levels of several viral proteins: BHRF1, EBNA-1, -2, -Leader Protein (LP), -3A, -3B, 3C and LMP-1, -2A, -2B. The EBV products include also non-coding RNAs: EBERs and microRNAs (miRNAs) including BamHI A rightward transcripts (miR-BARTs) and miR-BHRF1 molecules [13]. There are a few infection patterns with different viral gene expression profiles (Figure 2). One of them, the Latency I pattern, is restricted to the expression of EBNA-1, EBERs, and miR-BARTs. The small set of EBV products implicates limited anti-apoptotic effect of EBV infection. Latency I program occurs in BL tumor cells. Another expression pattern, Latency II, is characterized by the expression of three LMPs (LMP-1, -2A, and -2B) and the products present in Latency I. The program of Latency II is realized in HL, NPC, and NKTL. The exclusive expression of BARF1 has also been detected in the NPC cells displaying this program. Next, a latent viral promoter Wp-restricted latency includes synthesis of BHRF1, EBNA-1, -LP, -3A, -3B, -3C, EBERs, and miR-BARTs. This pattern is characteristic of some BL tumor cells. Another program, Latency III, is distinguished by the expression of the full range of proteins and non-coding RNAs (BHRF1, EBNA-1, -2, -LP, -3A, -3B, 3C, LMP-1, -2A, -2B, EBERs, miR-BARTs, and miR-BHRF1s), and observed in vitro in resting B-lymphocytes converting into lymphoblastoid cell lines (LCLs). This pattern occurs also in DLBCL [13][42][55][56][57][58][59][60].
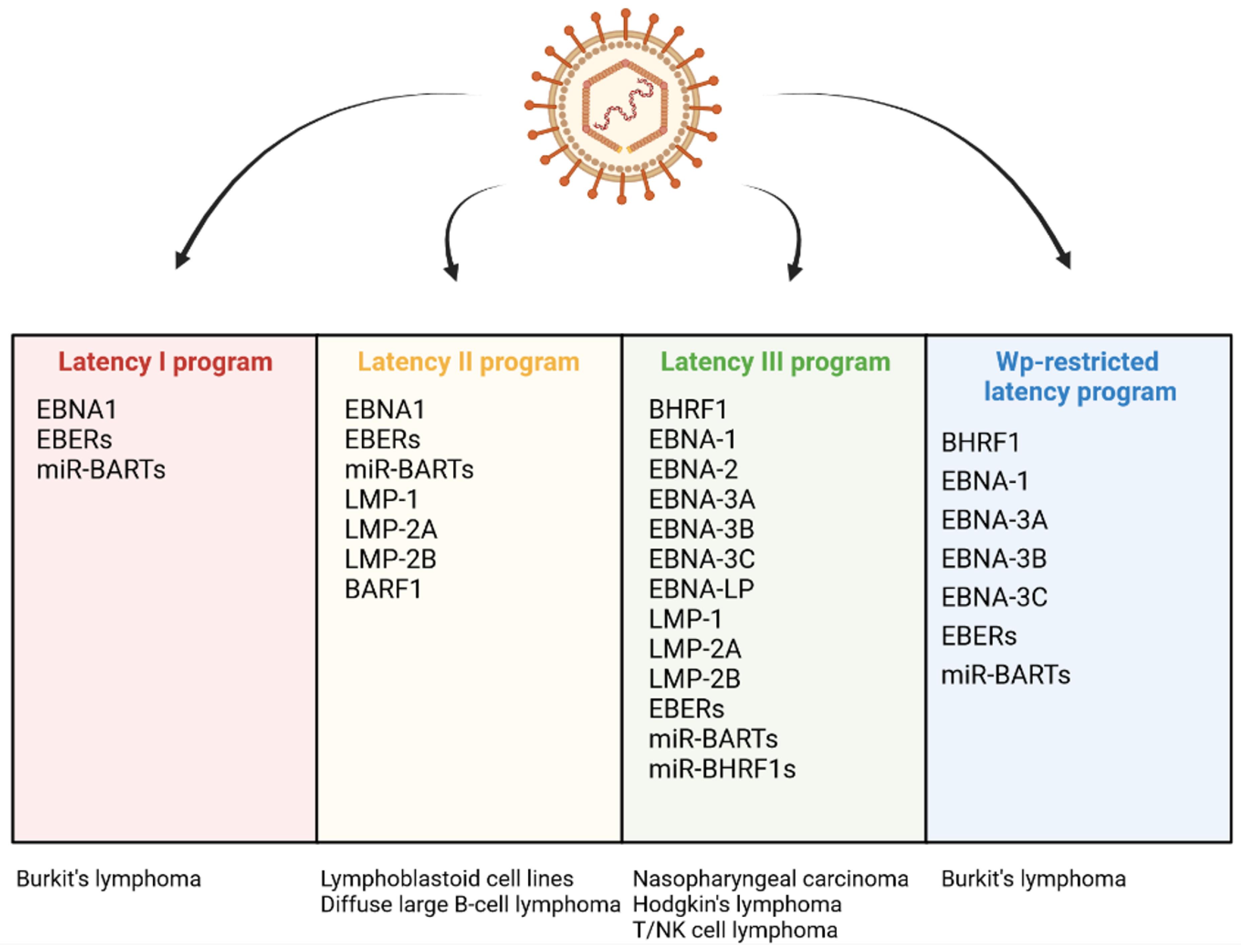
Figure 2. Differential expression programs in distinct types of EBV latency and various EBV-associated diseases.
In some cases, the establishment of latency may be followed by a switch to a productive cycle aimed at the formation and release of progeny virions [61][62]. The lytic cycle consists of three stages: the immediate-early (IE) phase, the early one, and the late one. In the first phase, the IE proteins are involved in the transcription of the viral genetic material. Next, the products of the early genes act to mediate DNA replication. In the third stage of the lytic cycle, the late proteins are engaged in the formation of progeny virions. The complete productive cycle ends in the infected cell lysis [63]. The switch between the latent period and the lytic cycle depends on the expression of the genes encoding two IE products, BamHI Z EBV replication activator (ZEBRA/BZLF1) and BRLF1 [62][64]. These genes are inactive during the latent infection, whereas upon the lytic phase, they undergo activation. ZEBRA and BRLF1 play the role of transcription factors responsible for the induction of the expression of the early genes encoding a set of lytic proteins, including BHRF1, BARF1, DNA primase (BSLF1), mRNA export factor ICP27 homolog (BMLF1/BSLF2), major DNA-binding proteins (BALF)2 and 5, primase-associated factors (BBLF)2/3 and 4, DNA polymerase processivity factor (BMRF1), thymidine kinase (TK), uracil-DNA glycosylase (BKRF3) and tegument protein (BKRF4) [62][65][66][67][68]. The replication of viral DNA, occurring in the early phase of the lytic cycle, is followed by the production of the late proteins (i.e., structural molecules such as major tegument protein (BNRF1), viral capsid antigen (VCA), major capsid protein (MCP), envelope glycoprotein gp350 (BLLF1), and envelope glycoprotein B (gB)). The progeny virions are assembled from the late viral products. Interestingly, the viral particles leave the nucleus and the cell, acquiring and losing sequentially the envelopes derived from the inner nuclear membrane and Golgi apparatus, respectively. The release of the virions to the extracellular environment occurs via the fusion of the organelle-derived envelope with the plasma membrane [62][66].
The lytic cycle leads to the multiplication of the virus and its effective spreading between the host cells. As a way of the carcinogenic pathogen propagation, it may favor the EBV-associated oncogenesis, contributing to the progression of both lymphoid and epithelial malignancies. Moreover, the individual viral lytic proteins may perform various oncogenic activities. They have been found to induce the genomic aberration, promote the expression and secretion of pro-inflammatory cytokines, counteract NK cell and CD8+ T cell response, increase angiogenesis and tumor invasiveness, as well as down-regulate major histocompatibility complexes (MHCs) and suppress MHC class II-restricted antigen presentation [63].
BHRF1 and BARF1 are present in the infected cells in some cases of latency, but also during the lytic cycle. The anti-apoptotic activity of these proteins increases the viability of the cell and delays its death in order to preserve the environment of replication and enable the completion of the progeny virions formation [62][69]. BHRF1 and BARF1 are synthesized in the early phase of the lytic cycle. BHRF1 gene is also expressed during the transient pre-latent abortive lytic cycle, a phase that precedes the latent period of infection [62][70].
References
- Singh, R.; Letai, A.; Sarosiek, K. Regulation of apoptosis in health and disease: The balancing act of BCL-2 family proteins. Nat. Rev. Mol. Cell Biol. 2019, 20, 175.
- Ibtisham, F.; Zhao, Y.; Nawab, A.; Liguang, H.; Wu, J.; Xiao, M.; Zhao, Z.; An, L. The Effect of High Temperature on Viability, Proliferation, Apoptosis and Anti-oxidant Status of Chicken Embryonic Fibroblast Cells. Braz. J. Poult. Sci. 2018, 20, 463–470.
- Kim, M.; Kim, G.; Kim, D.; Yoo, J.; Kim, D.-K.; Kim, H. Numerical Study on Effective Conditions for the Induction of Apoptotic Temperatures for Various Tumor Aspect Ratios Using a Single Continuous-Wave Laser in Photothermal Therapy Using Gold Nanorods. Cancers 2019, 11, 764.
- Stanojevic-Pirkovic, M.; Andelkovic, M.; Lukovic, J.; Zelen, I.; Milosavljevic, Z.; Mitrovic, M.; Jurisic, V.; Nikolic, I. UV irradiation induces apoptosis in the human endometrial stromal cell line (ThESC). J. BUON 2020, 25, 1541–1546.
- Sample, A.; He, Y.Y. Autophagy in UV Damage Response. Photochem. Photobiol. 2017, 93, 943–955.
- Ritchhart, C.; Joy, A. Reversal of drug-induced gingival overgrowth by UV-mediated apoptosis of gingival fibroblasts—An in vitro study. Ann. Anat. 2018, 217, 7–11.
- Lim, J.R.; Lee, H.J.; Jung, Y.H.; Kim, J.S.; Chae, C.W.; Kim, S.Y.; Han, H.J. Ethanol-activated CaMKII signaling induces neuronal apoptosis through Drp1-mediated excessive mitochondrial fission and JNK1-dependent NLRP3 inflammasome activation. Cell Commun. Signal. 2020, 18, 123.
- Li, C.; Li, J.; Xu, G.; Sun, H. Influence of Chronic Ethanol Consumption on Apoptosis and Autophagy Following Transient Focal Cerebral Ischemia in Male Mice. Sci. Rep. 2020, 10, 6164.
- Bauer, C.; Stotter, C.; Jeyakumar, V.; Niculescu-Morzsa, E.; Simlinger, B.; Rodríguez Ripoll, M.; Klestil, T.; Franek, F.; Nehrer, S. Concentration-Dependent Effects of Cobalt and Chromium Ions on Osteoarthritic Chondrocytes. Cartilage 2019, 13, 908S–919S.
- Genchi, G.; Sinicropi, M.S.; Lauria, G.; Carocci, A.; Catalano, A. The Effects of Cadmium Toxicity. Int. J. Environ. Res. Public Health 2020, 17, 3782.
- Brokatzky, D.; Kretz, O.; Häcker, G. Apoptosis Functions in Defense against Infection of Mammalian Cells with Environmental Chlamydiae. Infect. Immun. 2020, 88, e00851-19.
- Carloni, G.; Fioretti, D.; Rinaldi, M.; Ponzetto, A. Heterogeneity and coexistence of oncogenic mechanisms involved in HCV-associated B-cell lymphomas. Crit. Rev. Oncol. Hematol. 2019, 138, 156–171.
- Fitzsimmons, L.; Cartlidge, R.; Chang, C.; Sejic, N.; Galbraith, L.C.A.; Suraweera, C.D.; Croom-Carter, D.; Dewson, G.; Tierney, R.J.; Bell, A.I.; et al. EBV BCL-2 homologue BHRF1 drives chemoresistance and lymphomagenesis by inhibiting multiple cellular pro-apoptotic proteins. Cell Death Differ. 2020, 27, 1554–1568.
- Jahan, S.; Khaliq, S.; Siddiqi, M.H.; Ijaz, B.; Ahmad, W.; Ashfaq, U.A.; Hassan, S. Anti-apoptotic effect of HCV core gene of genotype 3a in Huh-7 cell line. Virol. J. 2011, 8, 1–7.
- Hösel, M.; Quasdorff, M.; Ringelhan, M.; Kashkar, H.; Debey-Pascher, S.; Sprinzl, M.F.; Bockmann, J.-H.; Arzberger, S.; Webb, D.; von Olshausen, G.; et al. Hepatitis B Virus Activates Signal Transducer and Activator of Transcription 3 Supporting Hepatocyte Survival and Virus Replication. Cell. Mol. Gastroenterol. Hepatol. 2017, 4, 339.
- Ryerson, M.R.; Richards, M.M.; Kvansakul, M.; Hawkins, C.J.; Shisler, J.L. Vaccinia Virus Encodes a Novel Inhibitor of Apoptosis That Associates with the Apoptosome. J. Virol. 2017, 91, e01385-17.
- Veyer, D.L.; Carrara, G.; Maluquer de Motes, C.; Smith, G.L. Vaccinia virus evasion of regulated cell death. Immunol. Lett. 2017, 186, 68–80.
- Lin, S.; Zhang, Y.J. Interference of apoptosis by hepatitis B virus. Viruses 2017, 9, 230.
- Manners, O.; Murphy, J.C.; Coleman, A.; Hughes, D.J.; Whitehouse, A. Contribution of the KSHV and EBV lytic cycles to tumourigenesis. Curr. Opin. Virol. 2018, 32, 60–70.
- Hawkins, J.B.; Delgado-Eckert, E.; Thorley-Lawson, D.A.; Shapiro, M. The Cycle of EBV Infection Explains Persistence, the Sizes of the Infected Cell Populations and Which Come under CTL Regulation. PLoS Pathog. 2013, 9, e1003685.
- He, H.-P.; Luo, M.; Cao, Y.-L.; Lin, Y.-X.; Zhang, H.; Zhang, X.; Ou, J.-Y.; Yu, B.; Chen, X.; Xu, M.; et al. Structure of Epstein-Barr virus tegument protein complex BBRF2-BSRF1 reveals its potential role in viral envelopment. Nat. Commun. 2020, 11, 5405.
- Lee, C.P.; Chen, M.R. Conquering the nuclear envelope barriers by ebv lytic replication. Viruses 2021, 13, 702.
- Zanella, L.; Riquelme, I.; Buchegger, K.; Abanto, M.; Ili, C.; Brebi, P. A reliable Epstein-Barr Virus classification based on phylogenomic and population analyses. Sci. Rep. 2019, 9, 9829.
- Cai, Q.; Chen, K.; Young, K.H. Epstein–Barr virus-positive T/NK-cell lymphoproliferative disorders. Exp. Mol. Med. 2015, 47, e133.
- Dugan, J.P.; Coleman, C.B.; Haverkos, B. Opportunities to Target the Life Cycle of Epstein-Barr Virus (EBV) in EBV-Associated Lymphoproliferative Disorders. Front. Oncol. 2019, 9, 127.
- Saha, A.; Robertson, E.S. Impact of EBV essential nuclear protein EBNA-3C on B-cell proliferation and apoptosis. Future Microbiol. 2013, 8, 323.
- Caetano, B.F.R.; Jorge, B.A.S.; Müller-Coan, B.G.; Elgui de Oliveira, D. Epstein-Barr virus microRNAs in the pathogenesis of human cancers. Cancer Lett. 2021, 499, 14–23.
- Graham, B.S.; Lynch, D.T. Cancer, Burkitt Lymphoma; StatPearls Publishing: Treasure Island, FL, USA, 2019.
- Hutcheson, R.L.; Chakravorty, A.; Sugden, B. Burkitt Lymphomas Evolve to Escape Dependencies on Epstein-Barr Virus. Front. Cell. Infect. Microbiol. 2021, 10, 606412.
- Sinclair, A.J.; Moalwi, M.H.; Amoaten, T. Is ebv associated with breast cancer in specific geographic locations? Cancers 2021, 13, 819.
- Stanland, L.J.; Luftig, M.A. The role of ebv-induced hypermethylation in gastric cancer tumorigenesis. Viruses 2020, 12, 1222.
- Grewal, R.; Irimie, A.; Naidoo, N.; Mohamed, N.; Petrushev, B.; Chetty, M.; Tomuleasa, C.; Abayomi, E.A. Hodgkin’s lymphoma and its association with EBV and HIV infection. Crit. Rev. Clin. Lab. Sci. 2018, 55, 102–114.
- Knecht, H.; Mai, S. LMP1 and dynamic progressive telomere dysfunction: A major culprit in EBV-associated Hodgkin’s lymphoma. Viruses 2017, 9, 164.
- Crombie, J.L.; LaCasce, A.S. Epstein Barr virus associated B-cell lymphomas and iatrogenic lymphoproliferative disorders. Front. Oncol. 2019, 9, 109.
- Murthy, S.L.; Hitchcock, M.A.; Endicott-Yazdani, T.R.; Watson, J.T.; Krause, J.R. Epstein-Barr virus–positive diffuse large B-cell lymphoma. Proc. (Bayl. Univ. Med. Cent.) 2017, 30, 443.
- Shannon-Lowe, C.; Rickinson, A.B.; Bell, A.I. Epstein-barr virus-associated lymphomas. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160271.
- Becnel, D.; Abdelghani, R.; Nanbo, A.; Avilala, J.; Kahn, J.; Li, L.; Lin, Z. Pathogenic role of epstein–barr virus in lung cancers. Viruses 2021, 13, 877.
- Hong, S.; Liu, D.; Luo, S.; Fang, W.; Zhan, J.; Fu, S.; Zhang, Y.; Wu, X.; Zhou, H.; Chen, X.; et al. The genomic landscape of Epstein-Barr virus-associated pulmonary lymphoepithelioma-like carcinoma. Nat. Commun. 2019, 10, 3108.
- Kheir, F.; Zhao, M.; Strong, M.J.; Yu, Y.; Nanbo, A.; Flemington, E.K.; Morris, G.F.; Reiss, K.; Li, L.; Lin, Z. Detection of Epstein-Barr Virus Infection in Non-Small Cell Lung Cancer. Cancers 2019, 11, 759.
- Sun, K.; Jia, K.; Lv, H.; Wang, S.Q.; Wu, Y.; Lei, H.; Chen, X. EBV-Positive Gastric Cancer: Current Knowledge and Future Perspectives. Front. Oncol. 2020, 10, 2096.
- Tavakoli, A.; Monavari, S.H.; Solaymani Mohammadi, F.; Kiani, S.J.; Armat, S.; Farahmand, M. Association between Epstein-Barr virus infection and gastric cancer: A systematic review and meta-analysis. BMC Cancer 2020, 20, 493.
- Yang, J.; Liu, Z.; Zeng, B.; Hu, G.; Gan, R. Epstein–Barr virus-associated gastric cancer: A distinct subtype. Cancer Lett. 2020, 495, 191–199.
- Fan, C.; Tang, Y.; Wang, J.; Guo, C.; Wang, Y.; Xiang, B.; Zhou, M.; Li, X.; Wu, X.; Li, Y.; et al. The emerging role of Epstein-Barr virus encoded microRNAs in nasopharyngeal carcinoma. J. Cancer 2018, 9, 2852.
- Hau, P.M.; Lung, H.L.; Wu, M.; Tsang, C.M.; Wong, K.L.; Mak, N.K.; Lo, K.W. Targeting Epstein-Barr Virus in Nasopharyngeal Carcinoma. Front. Oncol. 2020, 10, 600.
- Tsao, S.W.; Tsang, C.M.; Lo, K.W. Epstein–Barr virus infection and nasopharyngeal carcinoma. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160270.
- Chen, Z.; Gao, L.; Wang, M.; Tang, Y.; Zhao, S.; Liu, W. Epstein-Barr virus positive peripheral T cell lymphoma with novel variants in STAT5B of a pediatric patient: A case report. BMC Cancer 2018, 18, 373.
- Allen, P.B.; Lechowicz, M.J. Management of NK/T-Cell Lymphoma, Nasal Type. J. Oncol. Pract. 2019, 15, 513–520.
- Tse, E.; Kwong, Y.L. The diagnosis and management of NK/T-cell lymphomas. J. Hematol. Oncol. 2017, 10, 85.
- Camilleri-Broët, S.; Davi, F.; Feuillard, J.; Bourgeois, C.; Seilhean, D.; Hauw, J.J.; Raphaël, M. High Expression of Latent Membrane Protein 1 of Epstein-Barr Virus and BCL-2 Oncoprotein in Acquired Immunodeficiency Syndrome-Related Primary Brain Lymphomas. Blood 1995, 86, 432–435.
- Buonavoglia, A.; Leone, P.; Prete, M.; Solimando, A.G.; Guastadisegno, C.; Lanave, G.; Camero, M.; Martella, V.; Muzio, L.L.; Racanelli, V. Epstein–Barr Virus in Salivary Samples from Systemic Lupus Erythematosus Patients with Oral Lesions. J. Clin. Med. 2021, 10, 4995.
- Janz, S. Myc translocations in B cell and plasma cell neoplasms. DNA Repair 2006, 5, 1213–1224.
- Hui, K.F.; Yeung, P.L.; Tam, K.P.; Chiang, A.K.S. Counteracting survival functions of EBNA3C in Epstein-Barr virus (EBV)-driven lymphoproliferative diseases by combination of SAHA and bortezomib. Oncotarget 2018, 9, 25101–25114.
- Sun, X.; Barlow, E.A.; Ma, S.; Hagemeier, S.R.; Duellman, S.J.; Burgess, R.R.; Tellam, J.; Khanna, R.; Kenney, S.C. Hsp90 inhibitors block outgrowth of EBV-infected malignant cells in vitro and in vivo through an EBNA1-dependent mechanism. Proc. Natl. Acad. Sci. USA 2010, 107, 3146–3151.
- Ma, J.; Nie, K.; Redmond, D.; Liu, Y.; Elemento, O.; Knowles, D.M.; Tam, W. EBV-miR-BHRF1-2 targets PRDM1/Blimp1: Potential role in EBV lymphomagenesis. Leukemia 2016, 30, 594–604.
- Fitzsimmons, L.; Boyce, A.J.; Wei, W.; Chang, C.; Croom-Carter, D.; Tierney, R.J.; Herold, M.J.; Bell, A.I.; Strasser, A.; Kelly, G.L.; et al. Coordinated repression of BIM and PUMA by Epstein-Barr virus latent genes maintains the survival of Burkitt lymphoma cells. Cell Death Differ. 2018, 25, 241–254.
- Wang, M.; Gu, B.; Chen, X.; Wang, Y.; Li, P.; Wang, K. The Function and Therapeutic Potential of Epstein-Barr Virus-Encoded MicroRNAs in Cancer. Mol. Ther.-Nucleic Acids 2019, 17, 657–668.
- Poling, B.C.; Price, A.M.; Luftig, M.A.; Cullen, B.R. The Epstein-Barr Virus miR-BHRF1 microRNAs Regulate Viral Gene Expression in cis. Virology 2017, 512, 113.
- De Re, V.; Caggiari, L.; De Zorzi, M.; Fanotto, V.; Miolo, G.; Puglisi, F.; Cannizzaro, R.; Canzonieri, V.; Steffan, A.; Farruggia, P.; et al. Epstein-Barr virus BART microRNAs in EBV- associated Hodgkin lymphoma and gastric cancer. Infect. Agents Cancer 2020, 15, 42.
- Torres, K.; Landeros, N.; Wichmann, I.A.; Polakovicova, I.; Aguayo, F.; Corvalan, A.H. EBV miR-BARTs and human lncRNAs: Shifting the balance in competing endogenous RNA networks in EBV-associated gastric cancer. Biochim. Biophys. Acta-Mol. Basis Dis. 2021, 1867, 166049.
- Lo, A.K.F.; Kwok, W.L.; Sai, W.T.; Hing, L.W.; Hui, J.W.Y.; Ka, F.T.; Hayward, S.D.; Yiu, L.C.; Yu, L.L.; Takada, K.; et al. Epstein-Barr Virus Infection Alters Cellular Signal Cascades in Human Nasopharyngeal Epithelial Cells. Neoplasia 2006, 8, 173.
- Mckenzie, J.; El-Guindy, A. Epstein-Barr Virus Lytic Cycle Reactivation. Curr. Top. Microbiol. Immunol. 2015, 391, 237–261.
- Murata, T.; Sugimoto, A.; Inagaki, T.; Yanagi, Y.; Watanabe, T.; Sato, Y.; Kimura, H. Molecular Basis of Epstein–Barr Virus Latency Establishment and Lytic Reactivation. Viruses 2021, 13, 2344.
- Rosemarie, Q.; Sugden, B. Epstein-Barr Virus: How Its Lytic Phase Contributes to Oncogenesis. Microorganisms 2020, 8, 1824.
- Song, H.; Lim, Y.; Im, H.; Bae, J.M.; Kang, G.H.; Ahn, J.; Baek, D.; Kim, T.Y.; Yoon, S.S.; Koh, Y. Interpretation of EBV infection in pan-cancer genome considering viral life cycle: LiEB (Life cycle of Epstein-Barr virus). Sci. Rep. 2019, 9, 3465.
- Hoebe, E.; Wille, C.; Hagemeier, S.; Kenney, S.; Greijer, A.; Middeldorp, J. Epstein–Barr Virus Gene BARF1 Expression is Regulated by the Epithelial Differentiation Factor ΔNp63α in Undifferentiated Nasopharyngeal Carcinoma. Cancers 2018, 10, 76.
- Novalic, Z.; van Rossen, T.M. Agents and Approaches for Lytic Induction Therapy of Epstein-Barr Virus Associated Malignancies. Med. Chem. 2016, 6, 449–466.
- Chen, L.W.; Hung, C.H.; Wang, S.S.; Yen, J.B.; Liu, A.C.; Hung, Y.H.; Chang, P.J. Expression and regulation of the BKRF2, BKRF3 and BKRF4 genes of Epstein-Barr virus. Virus Res. 2018, 256, 76–89.
- Oudejans, J.J.; Van Den Brule, A.J.C.; Jiwa, N.M.; De Bruin, P.C.; Ossenkoppele, G.J.; Van Der Valk, P.; Walboomers, J.M.M.; Meijer, C.J.L.M. BHRF1, the Epstein-Barr Virus (EBV) Homologue of the BCL-2 Proto-oncogene, Is Transcribed in EBV-Associated B-Cell Lymphomas and in Reactive Lymphocytes. Blood 1995, 86, 1893–1902.
- Lo, A.K.F.; Dawson, C.W.; Lung, H.L.; Wong, K.L.; Young, L.S. The Therapeutic Potential of Targeting BARF1 in EBV-Associated Malignancies. Cancers 2020, 12, 1940.
- Germini, D.; Sall, F.B.; Shmakova, A.; Wiels, J.; Dokudovskaya, S.; Drouet, E.; Vassetzky, Y. Oncogenic Properties of the EBV ZEBRA Protein. Cancers 2020, 12, 1479.
More
Information
Subjects:
Biochemistry & Molecular Biology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.5K
Revisions:
2 times
(View History)
Update Date:
18 Jul 2022
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No