 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Junfa Li | -- | 2954 | 2022-07-01 08:50:34 | | | |
| 2 | Sirius Huang | Meta information modification | 2954 | 2022-07-01 11:27:32 | | |
Video Upload Options
Synapses serve as the interface for the transmission of information between neurons in the central nervous system. The structural and functional characteristics of synapses are highly dynamic, exhibiting extensive plasticity that is shaped by neural activity and regulated primarily by trans-synaptic cell-adhesion molecules (CAMs). Prototypical trans-synaptic CAMs, such as neurexins (Nrxs) and neuroligins (Nlgs), directly regulate the assembly of presynaptic and postsynaptic molecules, including synaptic vesicles, active zone proteins, and receptors. Therefore, the trans-synaptic adhesion mechanisms mediated by Nrx–Nlg interaction can contribute to a range of synaptopathies in the context of pathological pain and other neurological disorders.
1. The Neurexin–Neuroligin Mediated Trans-Synaptic Modulation
1.1. The Dynamic Synaptic Regulation by Neurexins
1.2. The Dynamic Synaptic Regulation by Neuroligins
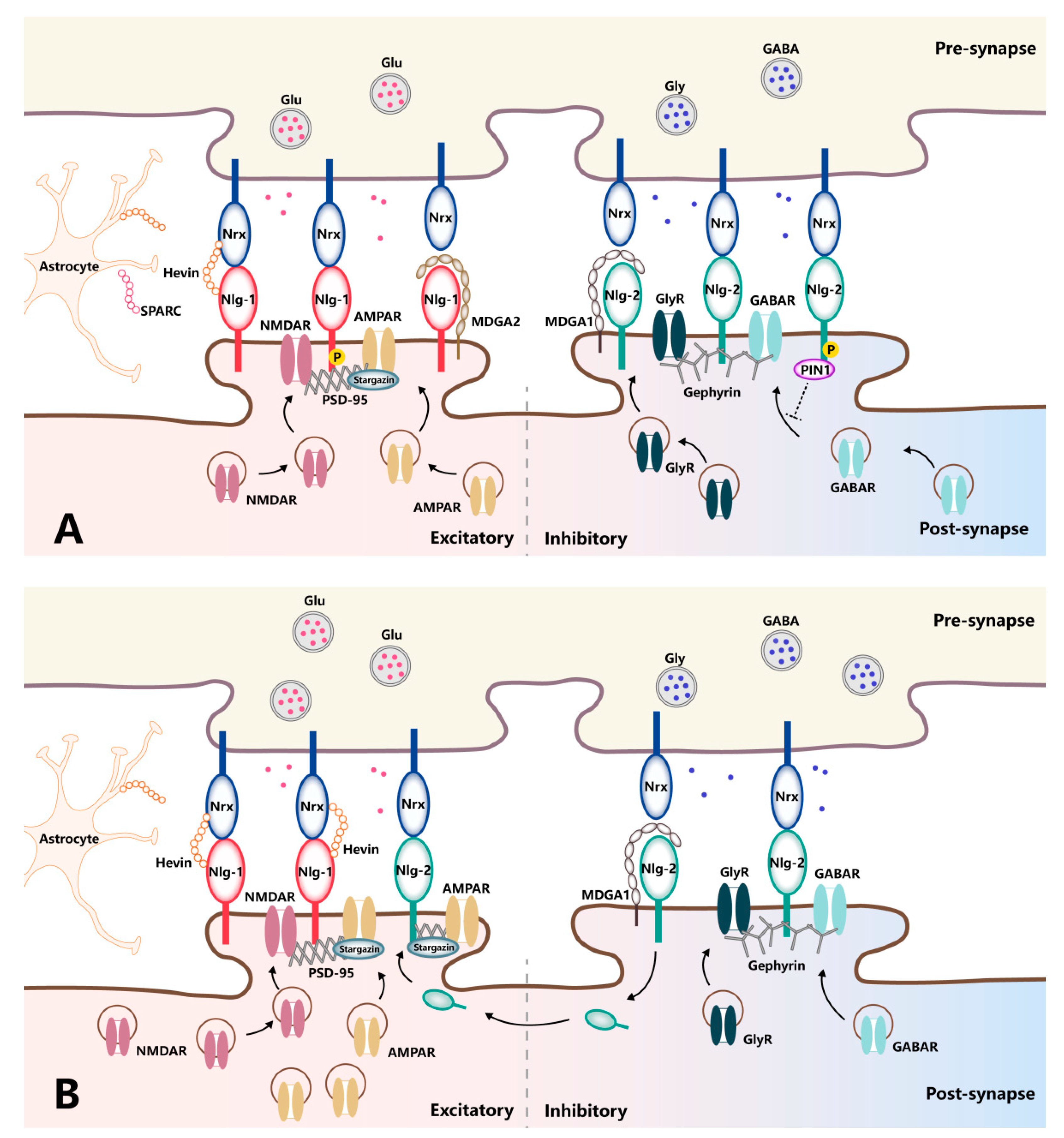
2. The Regulators of Neurexin–Neuroligin Interaction
2.1. Hevin
2.2. SPARC
2.3. MDGA
References
- Lin, T.B.; Lai, C.Y.; Hsieh, M.C.; Jiang, J.L.; Cheng, J.K.; Chau, Y.P.; Ruan, T.; Chen, G.D.; Peng, H.Y. Neuropathic allodynia involves spinal neurexin-1β-dependent neuroligin-1/postsynaptic density-95/NR2B cascade in rats. Anesthesiology 2015, 123, 909–926.
- Graf, E.R.; Zhang, X.; Jin, S.X.; Linhoff, M.W.; Craig, A.M. Neurexins induce differentiation of GABA and glutamate postsynaptic specializations via neuroligins. Cell 2004, 119, 1013–1026.
- Südhof, T.C. Towards an Understanding of Synapse Formation. Neuron 2018, 100, 276–293.
- Luo, F.; Sclip, A.; Merrill, S.; Südhof, T.C. Neurexins regulate presynaptic GABAB-receptors at central synapses. Nat. Commun. 2021, 12, 2380.
- Roberts, B.M.; Lopes, E.F.; Cragg, S.J. Axonal modulation of striatal dopamine release by local γ-aminobutyric acid (GABA) signalling. Cells 2021, 10, 709.
- Luo, F.; Sclip, A.; Jiang, M.; Südhof, T.C. Neurexins cluster Ca2+ channels within the presynaptic active zone. EMBO J. 2020, 39, e103208.
- Sánchez-Hidalgo, A.C.; Arias-Aragón, F.; Romero-Barragán, M.T.; Martín-Cuevas, C.; Delgado-García, J.M.; Martinez-Mir, A.; Scholl, F.G. Selective expression of the neurexin substrate for presenilin in the adult forebrain causes deficits in associative memory and presynaptic plasticity. Exp. Neurol. 2022, 347, 113896.
- Anderson, G.R.; Aoto, J.; Tabuchi, K.; Földy, C.; Covy, J.; Yee, A.X.; Wu, D.; Lee, S.J.; Chen, L.; Malenka, R.C.; et al. β-neurexins control neural circuits by regulating synaptic endocannabinoid signaling. Cell 2015, 162, 593–606.
- Aoto, J.; Martinelli, D.C.; Malenka, R.C.; Tabuchi, K.; Südhof, T.C. Presynaptic neurexin-3 alternative splicing trans-synaptically controls postsynaptic AMPA receptor trafficking. Cell 2013, 154, 75–88.
- Dai, J.; Aoto, J.; Südhof, T.C. Alternative splicing of presynaptic neurexins differentially controls postsynaptic NMDA and AMPA receptor responses. Neuron 2019, 102, 993–1008.e5.
- Miyazaki, T.; Morimoto-Tomita, M.; Berthoux, C.; Konno, K.; Noam, Y.; Yamasaki, T.; Verhage, M.; Castillo, P.E.; Watanabe, M.; Tomita, S. Excitatory and inhibitory receptors utilize distinct post- and trans-synaptic mechanisms in vivo. Elife 2021, 10, e59613.
- Boxer, E.E.; Seng, C.; Lukacsovich, D.; Kim, J.; Schwartz, S.; Kennedy, M.J.; Földy, C.; Aoto, J. Neurexin-3 defines synapse- and sex-dependent diversity of GABAergic inhibition in ventral subiculum. Cell Rep. 2021, 37, 110098.
- Sato, Y.; Iijima, Y.; Darwish, M.; Sato, T.; Iijima, T. Distinct expression of SLM2 underlies splicing-dependent trans-synaptic signaling of neurexin across GABAergic neuron subtypes. Neurochem. Res. 2021, in press.
- Trotter, J.H.; Hao, J.; Maxeiner, S.; Tsetsenis, T.; Liu, Z.; Zhuang, X.; Südhof, T.C. Synaptic neurexin-1 assembles into dynamically regulated active zone nanoclusters. J. Cell Biol. 2019, 218, 2677–2698.
- Klatt, O.; Repetto, D.; Brockhaus, J.; Reissner, C.; El Khallouqi, A.; Rohlmann, A.; Heine, M.; Missler, M. Endogenous β-neurexins on axons and within synapses show regulated dynamic behavior. Cell Rep. 2021, 35, 109266.
- Saura, C.A.; Servián-Morilla, E.; Scholl, F.G. Presenilin/γ-secretase regulates neurexin processing at synapses. PLoS ONE 2011, 6, e19430.
- Zhang, P.; Lu, H.; Peixoto, R.T.; Pines, M.K.; Ge, Y.; Oku, S.; Siddiqui, T.J.; Xie, Y.; Wu, W.; Archer-Hartmann, S.; et al. Heparan sulfate organizes neuronal synapses through neurexin partnerships. Cell 2018, 174, 1450–1464.e23.
- Hu, X.; Luo, J.H.; Xu, J. The interplay between synaptic activity and neuroligin function in the CNS. Biomed. Res. Int. 2015, 2015, 498957.
- Varoqueaux, F.; Aramuni, G.; Rawson, R.L.; Mohrmann, R.; Missler, M.; Gottmann, K.; Zhang, W.; Südhof, T.C.; Brose, N. Neuroligins determine synapse maturation and function. Neuron 2006, 51, 741–754.
- Taylor, S.C.; Ferri, S.L.; Grewal, M.; Smernoff, Z.; Bucan, M.; Weiner, J.A.; Abel, T.; Brodkin, E.S. The role of synaptic cell adhesion molecules and associated scaffolding proteins in social affiliative behaviors. Biol. Psychiatry 2020, 88, 442–451.
- Bemben, M.A.; Shipman, S.L.; Nicoll, R.A.; Roche, K.W. The cellular and molecular landscape of neuroligins. Trends Neurosci. 2015, 38, 496–505.
- Prange, O.; Wong, T.P.; Gerrow, K.; Wang, Y.T.; El-Husseini, A. A balance between excitatory and inhibitory synapses is controlled by PSD-95 and neuroligin. Proc. Natl. Acad. Sci. USA 2004, 101, 13915–13920.
- Ali, H.; Marth, L.; Krueger-Burg, D. Neuroligin-2 as a central organizer of inhibitory synapses in health and disease. Sci. Signal. 2020, 13, eabd8379.
- Chubykin, A.A.; Atasoy, D.; Etherton, M.R.; Brose, N.; Kavalali, E.T.; Gibson, J.R.; Südhof, T.C. Activity-dependent validation of excitatory versus inhibitory synapses by neuroligin-1 versus neuroligin-2. Neuron 2007, 54, 919–931.
- Chubykin, A.A.; Atasoy, D.; Etherton, M.R.; Brose, N.; Kavalali, E.T.; Gibson, J.R.; Südhof, T.C. Neuroligin-1 controls synaptic abundance of NMDA-type glutamate receptors through extracellular coupling. Proc. Natl. Acad. Sci. USA 2013, 110, 725–730.
- Jiang, M.; Polepalli, J.; Chen, L.Y.; Zhang, B.; Südhof, T.C.; Malenka, R.C. Conditional ablation of neuroligin-1 in CA1 pyramidal neurons blocks LTP by a cell-autonomous NMDA receptor-independent mechanism. Mol. Psychiatry 2017, 22, 375–383.
- Wu, X.; Morishita, W.K.; Riley, A.M.; Hale, W.D.; Südhof, T.C.; Malenka, R.C. Neuroligin-1 signaling controls LTP and NMDA receptors by distinct molecular pathways. Neuron 2019, 102, 621–635.e3.
- Hines, R.M.; Wu, L.; Hines, D.J.; Steenland, H.; Mansour, S.; Dahlhaus, R.; Singaraja, R.R.; Cao, X.; Sammler, E.; Hormuzdi, S.G.; et al. Synaptic imbalance, stereotypies, and impaired social interactions in mice with altered neuroligin 2 expression. J. Neurosci. 2008, 28, 6055–6067.
- Troyano-Rodriguez, E.; Wirsig-Wiechmann, C.R.; Ahmad, M. Neuroligin-2 determines inhibitory synaptic transmission in the lateral septum to optimize stress-induced neuronal activation and avoidance behavior. Biol. Psychiatry 2019, 85, 1046–1055.
- Uchigashima, M.; Konno, K.; Demchak, E.; Cheung, A.; Watanabe, T.; Keener, D.G.; Abe, M.; Le, T.; Sakimura, K.; Sasaoka, T.; et al. Specific Neuroligin3-αNeurexin1 signaling regulates GABAergic synaptic function in mouse hippocampus. Elife 2020, 9, e59545.
- Uchigashima, M.; Leung, M.; Watanabe, T.; Cheung, A.; Le, T.; Pallat, S.; Dinis, A.L.M.; Watanabe, M.; Kawasawa, Y.I.; Futai, K. Neuroligin3 splice isoforms shape inhibitory synaptic function in the mouse hippocampus. J. Biol. Chem. 2020, 295, 8589–8595.
- Bemben, M.A.; Shipman, S.L.; Hirai, T.; Herring, B.E.; Li, Y.; Badger, J.D.; Nicoll, R.A.; Diamond, J.S.; Roche, K.W. CaMKII phosphorylation of neuroligin-1 regulates excitatory synapses. Nat. Neurosci. 2014, 7, 56–64.
- Jeong, J.; Pandey, S.; Li, Y.; Badger, J.D.; Lu, W.; Roche, K.W. PSD-95 binding dynamically regulates NLGN1 trafficking and function. Proc. Natl. Acad. Sci. USA 2019, 116, 12035–12044.
- Letellier, M.; Szíber, Z.; Chamma, I.; Saphy, C.; Papasideri, I.; Tessier, B.; Sainlos, M.; Czöndör, K.; Thoumine, O. A unique intracellular tyrosine in neuroligin-1 regulates AMPA receptor recruitment during synapse differentiation and potentiation. Nat. Commun. 2018, 9, 3979.
- Antonelli, R.; Pizzarelli, R.; Pedroni, A.; Fritschy, J.M.; Del-Sal, G.; Cherubini, E.; Zacchi, P. Pin1-dependent signalling negatively affects GABAergic transmission by modulating neuroligin2/gephyrin interaction. Nat. Commun. 2014, 5, 5066.
- Letellier, M.; Lagardère, M.; Tessier, B.; Janovjak, H.; Thoumine, O. Optogenetic control of excitatory post-synaptic differentiation through neuroligin-1 tyrosine phosphorylation. Elife 2020, 9, e52027.
- Suzuki, K.; Hayashi, Y.; Nakahara, S.; Kumazaki, H.; Prox, J.; Horiuchi, K.; Zeng, M.; Tanimura, S.; Nishiyama, Y.; Osawa, S.; et al. Activity-dependent proteolytic cleavage of neuroligin-1. Neuron 2012, 76, 410–422.
- Peixoto, R.T.; Kunz, P.A.; Kwon, H.; Mabb, A.M.; Sabatini, B.L.; Philpot, B.D.; Ehlers, M.D. Transsynaptic signaling by activity-dependent cleavage of neuroligin-1. Neuron 2012, 76, 396–409.
- Gjørlund, M.D.; Carlsen, E.M.M.; Kønig, A.B.; Dmytrieva, O.; Petersen, A.V.; Jacobsen, J.; Berezin, V.; Perrier, J.F.; Owczarek, S. Soluble ectodomain of neuroligin 1 decreases synaptic activity by activating metabotropic glutamate receptor 2. Front. Mol. Neurosci. 2017, 10, 116.
- Bemben, M.A.; Nguyen, T.A.; Li, Y.; Wang, T.; Nicoll, R.A.; Roche, K.W. Isoform-specific cleavage of neuroligin-3 reduces synapse strength. Mol. Psychiatry 2019, 24, 145–160.
- Murphy-Ullrich, J.E.; Sage, E.H. Revisiting the matricellular concept. Matrix Biol. 2014, 37, 1–14.
- Fan, S.; Gangwar, S.P.; Machius, M.; Rudenko, G. Interplay between hevin, SPARC, and MDGAs: Modulators of neurexin-neuroligin transsynaptic bridges. Structure 2021, 29, 664–678.e6.
- Singh, S.K.; Stogsdill, J.A.; Pulimood, N.S.; Dingsdale, H.; Kim, Y.H.; Pilaz, L.J.; Kim, I.H.; Manhaes, A.C.; Rodrigues, W.S., Jr.; Pamukcu, A.; et al. Astrocytes assemble thalamocortical synapses by bridging NRX1α and NL1 via hevin. Cell 2016, 164, 183–196.
- Kim, J.H.; Jung, H.G.; Kim, A.; Shim, H.S.; Hyeon, S.J.; Lee, Y.S.; Han, J.; Jung, J.H.; Lee, J.; Ryu, H.; et al. Hevin-calcyon interaction promotes synaptic reorganization after brain injury. Cell Death Differ. 2021, 28, 2571–2588.
- Lively, S.; Brown, I.R. The extracellular matrix protein SC1/hevin localizes to excitatory synapses following status epilepticus in the rat lithium-pilocarpine seizure model. J. Neurosci. Res. 2008, 86, 2895–2905.
- Vincent, A.J.; Lau, P.W.; Roskams, A.J. SPARC is expressed by macroglia and microglia in the developing and mature nervous system. Dev. Dyn. 2008, 237, 1449–1462.
- Kucukdereli, H.; Allen, N.J.; Lee, A.T.; Feng, A.; Ozlu, M.I.; Conatser, L.M.; Chakraborty, C.; Workman, G.; Weaver, M.; Sage, E.H.; et al. Control of excitatory CNS synaptogenesis by astrocyte-secreted proteins Hevin and SPARC. Proc. Natl. Acad. Sci. USA 2011, 108, E440–E449.
- Jones, E.V.; Bernardinelli, Y.; Tse, Y.C.; Chierzi, S.; Wong, T.P.; Murai, K.K. Astrocytes control glutamate receptor levels at developing synapses through SPARC-beta-integrin interactions. J. Neurosci. 2011, 31, 4154–4165.
- Jones, E.V.; Bernardinelli, Y.; Zarruk, J.G.; Chierzi, S.; Murai, K.K. SPARC and GluA1-containing AMPA receptors promote neuronal health following CNS injury. Front. Cell Neurosci. 2018, 12, 22.
- Litwack, E.D.; Babey, R.; Buser, R.; Gesemann, M.; O’Leary, D.D. Identification and characterization of two novel brain-derived immunoglobulin superfamily members with a unique structural organization. Mol. Cell. Neurosci. 2004, 25, 263–274.
- Connor, S.A.; Elegheert, J.; Xie, Y.; Craig, A.M. Pumping the brakes: Suppression of synapse development by MDGA-neuroligin interactions. Curr. Opin. Neurobiol. 2019, 57, 71–80.
- Kim, J.A.; Kim, D.; Won, S.Y.; Han, K.A.; Park, D.; Cho, E.; Yun, N.; An, H.J.; Um, J.W.; Kim, E.; et al. Structural insights into modulation of neurexin-neuroligin trans-synaptic adhesion by MDGA1/neuroligin-2 complex. Neuron 2017, 94, 1121–1131.e6.
- Elegheert, J.; Cvetkovska, V.; Clayton, A.J.; Heroven, C.; Vennekens, K.M.; Smukowski, S.N.; Regan, M.C.; Jia, W.; Smith, A.C.; Furukawa, H.; et al. Structural mechanism for modulation of synaptic neuroligin-neurexin signaling by MDGA proteins. Neuron 2021, 109, 189–190.
- Lee, K.; Kim, Y.; Lee, S.J.; Qiang, Y.; Lee, D.; Lee, H.W.; Kim, H.; Je, H.S.; Südhof, T.C.; Ko, J. MDGAs interact selectively with neuroligin-2 but not other neuroligins to regulate inhibitory synapse development. Proc. Natl. Acad. Sci. USA 2013, 110, 336–341.
- Connor, S.A.; Ammendrup-Johnsen, I.; Chan, A.W.; Kishimoto, Y.; Murayama, C.; Kurihara, N.; Tada, A.; Ge, Y.; Lu, H.; Yan, R.; et al. Altered cortical dynamics and cognitive function upon haploinsufficiency of the autism-linked excitatory synaptic suppressor MDGA2. Neuron 2016, 91, 1052–1068.

