1. Biochemical Aspects of Cannabinoids
Substances from the class of cannabinoids are found in high concentrations in the plant of
Cannabis sativa [1]. Out of the many structural forms of cannabinoids, the most widely studied are delta-9-tetrahydrocannabinol (Δ9-THC) and cannabidiol (CBD)
[2]. Nevertheless, there are over 100 molecular structures
[3] existing as types of C-terpenopenols, which, based on their atomic dispositions and their macrostructures, have been classified as delta-8-tetrahydrocannabinol (Δ8-THC), Δ9-THC, CBD, and cannabicyclol. Moreover, based on their origin, they can be classified as phytocannabinoids, endocannabinoids, and synthetic cannabinoids
[4]. The endocannabinoid system is involved in various processes, such as lipolysis, energy balance, metabolism, and cognition and behavior. The most important endocannabinoid system are the anandamide (AEA) and 2-arachidonoylglycerol (2-AG), which also have high concentrations in the brain
[5][6][7].
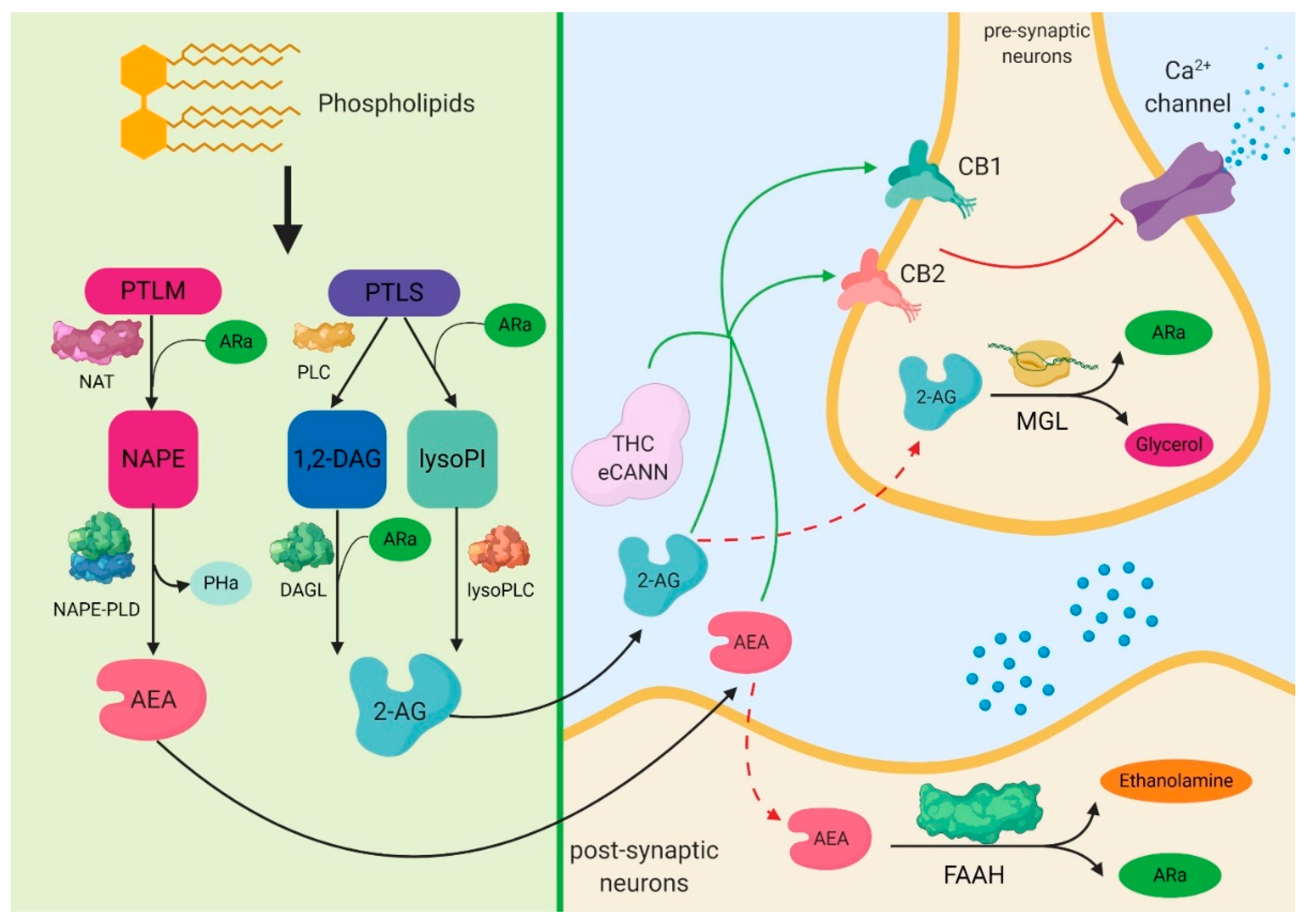
From biochemical point of view the endogenous cannabinoids are obtained through the action of
N-acyl-transferases on the endogenous phospholipids. The lipid precursor phosphatidylethanolamine (PTLM), through the action of
N-acyltransferase (NAT) and of arachidonic acid (ARa), leads to the formation of
N-arachidonoyl-phosphatidylethonolamine (NAPE), which, through the action of NAPE-phospholipase D (NAPE-PLD), forms
N-Arachidonoylethanolamine (anandamide—AEA) and phosphatidic acid (PHa). There are two pathways regarding the mechanisms that lead to the formation of 2-arachidonoylglycerol (2-AG). One of them is represented by the phospholipase C (PLC), which acts on the lipid precursor phosphatidylinositol (PTLS) to form 1,2-diacylglycerol (1,2-DAG). Through the action of diacylglycerol lipase (DAGL) and arachidonic acid (ARa) on 1,2-DAG, 2-AG is formed. Another pathway is represented by the action of ARa and phospholipase A1 on phosphatidylinositol, which leads to the formation of 2-arachidonoyl-lysophospholipid (lysoPI). This further leads to the formation of 2-AG through the action of lysophosphilipase C (Lyso-PLC). The result of this activation of cannabinoid receptors (CB) by different molecules, including endogenous and exogenous cannabinoids (eCANN) such as tetrahydrocannabinol (THC) or cannabidiol (CBD), will act on the calcium channels in order to decrease and inhibit, at a pre-synaptic level, the influx of calcium, therefore inhibiting the release of glutamate or gamma aminobutyric acid (GABA). The inactivation of endocannabinoids takes place by their transport inside the cell and by hydrolysis. Two different enzymes are involved in this reaction, monoacylglycerol lipase (MGL) for 2-AG, and fatty-acid amide hydrolase (FAAH) for AEA. These have been targets in recent research on the treatment of pain, with pharmacologic inhibitors being studied for both the deactivation of the endocannabinoid action pathway and for their transport
[7][8][9][10][11][12][13][14][15][16][17][18][19][20][21][22][23][24][25][26][27][28][29][30][31][32][33] (
Figure 1).
Figure 1. Molecular signaling mediated by the endocannabinoid system in the central nervous system. The most important endocannabinoid systems are the anandamide and the 2-arachidonoylglycerol, which also have high concentrations in the brain. Anandamide can be obtained through a series of biological mechanisms that involve the restructuring of phospholipids by the N-acyl-transferases. 2-arachidonoylglycerol results from the action of phospholipase C-beta enzymes or after the dephosphorylation of lysophosphatidic acid. Following these reactions, both anandamide and 2-arachidonoylglycerol activate the G-protein coupled receptors, transient receptor potential vanilloid type 1, and peroxisome proliferator-activated nuclear receptors. Finally, cyclooxygenase-2 can act on these two endocannabinoids through redox mechanisms, leading to the production of prostamides and prostaglandin-glycerol esters. PTLM: lipid precursor phosphatidylethanolamine; NAT: N-acyltransferase; ARa: arachidonic acid; NAPE: N-arachidonoyl-phosphatidylethonolamine; NAPE-PLD: NAPE-phospholipase D; PHa: phosphatidic acid; AEA: anandamide; PTLS: lipid precursor phosphatidylinositol; PLC: phospholipase C; 1,2-DAG: 1,2-diacylglycerol; lysoPI: 2-arachidonoyl-lysophospholipid; DAGL: diacylglycerol lipase; lysoPLC: lysophosphilipase C; 2-AG: 2-arachidonoylglycerol; CB1: cannabinoid receptor 1; CB2: cannabinoid receptor 2; THC: tetrahydrocannabinol; eCANN: endogenous and exogenous cannabinoids; MGL: monoacylglycerol lipase; FAAH: fatty-acid amide hydrolase.
The activity of cannabinoids depends solely on their interaction with certain cannabinoid receptors (CB), part of the G-protein coupled receptor family. CB1, CB2, and CB3 have been identified. From a biological point of view, CB1 can be found in the central nervous system
[13][14][15]. Their share is higher in certain regions, such as the amygdala, cortex, hypothalamus, hippocampus, and the cerebellum. CB2 can be mainly found in the immune system, with higher concentrations in lymphocytes, macrophages, and inside the spleen. CB3 has been identified inside the endothelial cell and in the spleen
[16]. Gαi is one of the most well understood biochemical pathways for the activation of CB1 and CB2 receptors, which also includes adenyl cyclase (Ac), mitogen activated protein kinase (MAPK), and cyclicadenosine monophosphate (cAMP)
[8][17][18].
2. The Expression of Cannabinoid Signaling System in Sepsis
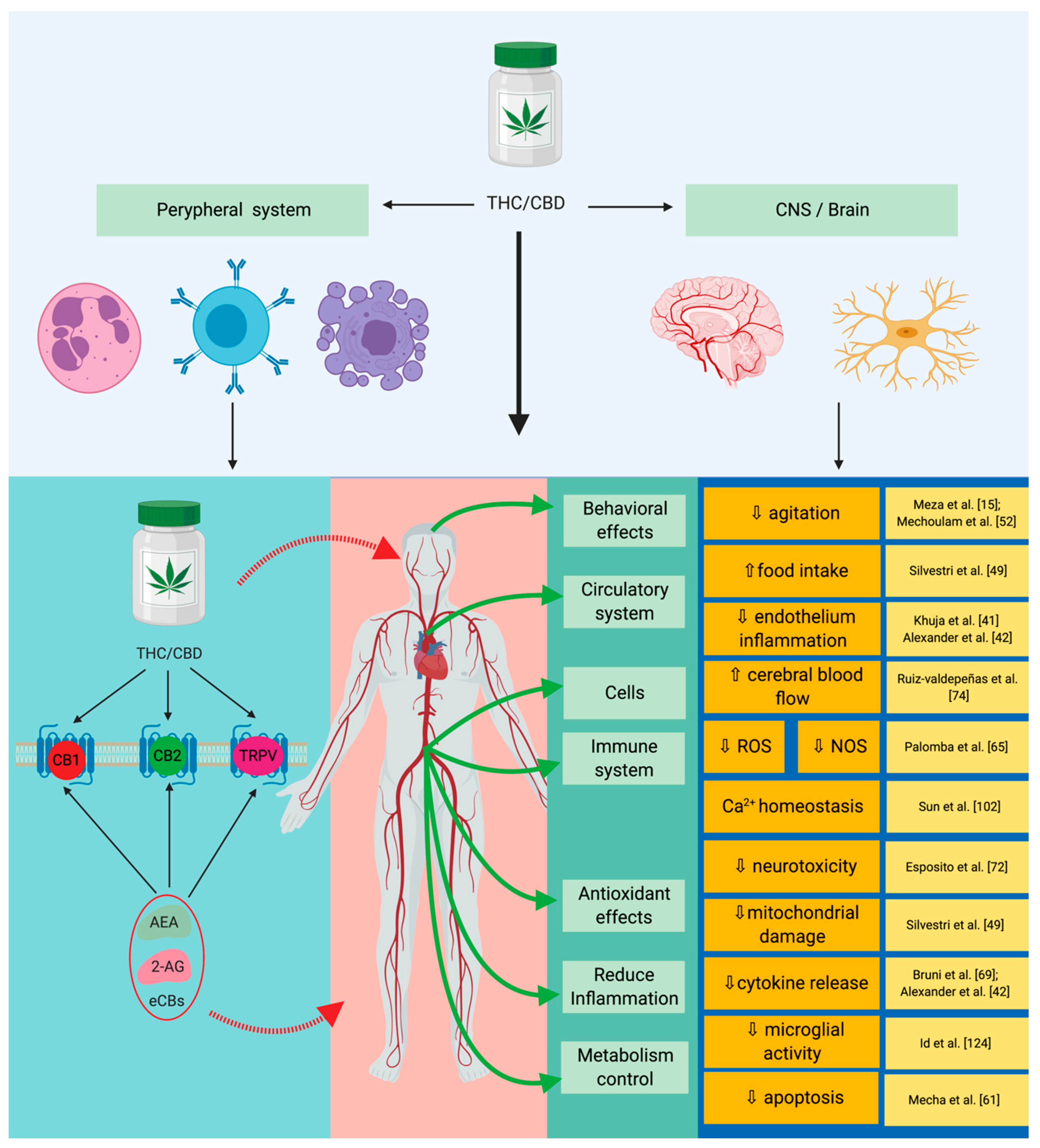
Recent studies have shown a series of anti-inflammatory, antioxidant, and antiapoptotic properties of cannabinoids in sepsis (
Figure 2). In practice, the link between the cannabinoid system and sepsis can be described in a simplistic manner, through its effects on inflammation, on the immune system, and on the redox activity (
Table 1)
[34].
Figure 2. Implications of the endocannabinoid system in sepsis. By acting on specific receptors CBD/THC lead to the suppression of cytokine production, as well as to a reduction in redox mechanisms. Through their synergistic action these will lead to the reduction of the systemic inflammatory response and to an immune modulation with neuronal protection. THC: tetrahydrocannabinol; CBD: cannabidiol; CB1: cannabinoid receptor 1; CB2: cannabinoid receptor 2; TRPV: transient receptor potential vanilloid type 1; AEA: anandamide; 2-AG: 2-arachidonoylglycerol; eCBs: endogenous cannabinoids.
Table 1. Endocannabinoid signaling system as a potential therapeutic target intervention.
A very important, but also specific, aspect for the septic patient is malnutrition. It can be induced either by the specific hypermetabolism, or by the under-dosed nutrition of patients in the ICU. Moreover, in the same context, a phenomenon that impacts the quality of life is represented by the recovery period and by the reintegration of the former patient in the society
[19][20][21][22].
Recent studies have concluded that endocannabinoids are characterized by a very fast response time to metabolic changes throughout the body. The activation of the CB1 receptor leads to the release of certain orexigenic peptides via neurons in the hypothalamus. What this means is that the stimulation of the CB1 receptor by Δ9-THC leads to an increased appetite for foods. These mechanisms are regulated by feeding-regulation hormones, such as orexigenic leptin, orexigenic ghrelin, and glucocorticoids. Another very important aspect regarding the impact of CB1 and CB2 receptors on the metabolism is given by the modulating properties of the CB1 on lypogenesis and on the control of the glucose levels. Some studies have shown that through the interaction of CB1 and 2-AG receptors, as well as with AES, one can increase the glucose metabolism, activate mitochondrial biosynthesis, and reduce adiponectin production
[16][23][24].
3. The Cannabinoid Signaling System and Inflammation-Linked with Sepsis
Cassol-Ju et al. carried out a study regarding the effects of cannabinoids on the pro-oxidative profile in the brain and other organs, and on cognitive function in lab animals with induced sepsis. Following their study, they reported a significant decrease in the prooxidative and proinflammatory profiles. Moreover, they observed a decrease in the expression of thiobarbituric acid reactive species (TBARs) and of protein carbonyl in subjects that benefitted from CBD
[39]. Furthermore, Cassol-Jr et al. also reported a decrease in mortality in subjects treated with CBD
[39]. Another interesting study was carried out by Prester et al., regarding the proinflammatory changes induces by cancer therapies with Irinotecan, and the effects Δ9-THC has in this field. One of the already known effects of Irinotecan therapies is the increase of aspartate aminotransferase (AST). Following this study, they observed a significant decrease in the AST levels in the group that received double therapy with irinotecan and Δ9-THC
[30]. Acute lung injury (ALI) and acute respiratory distress syndrome (ARDS) are two other pathologies frequently associated with sepsis, and yielding high mortality rates. From a morphopathological point of view, ALI and ARDS present similarities, such as the hyperinflammation of the lung tissues. From a clinical viewpoint, these contexts are associated with severe hypoxemia, leading to the impossibility of assuring effective ventilatory support. Regarding the mechanisms behind them, the causes can be divided into direct and indirect, such as lung injury, inhalation injury, pulmonary contusions, burns affecting the airway, disseminated or localized infections, and cellular lesions induced by massive blood transfusions. In the case of sepsis, the inflammatory profile is augmented by the recognition of certain bacterial antigens on the macrophage receptors, such as Toll-like receptors 2 and 4 (TLR2 and TLR4). This affects the integrity of the endothelial tissue, with the alveoli being exposed to a series of molecular and cellular attacks by fluids and macromolecules. Vuolo et al., in another study, focused on the evaluation of the anti-inflammatory activity of cannabinoids in asthma, have reported a decrease of IL-4, IL-5, IL-13, IL-6, and TNF-α. Following this study, the group highlighted the potential of cannabinoids to modulate the anti-inflammatory response in the context of asthma
[40]. Chiurchiu et al., in a similar study, proved a decrease in the IL-6, IL-12, and IFN-α levels in dendritic cells
[7]. In regard with the antimicrobial activity of
Cannabis sativa extracts, studies have reported antimicrobial characteristics targeting gram-positive (
Staphylococcus aureus,
Micrococcus flavus, and
Baccilus subtilus) and Gram-negative bacteria (
Proteus vulgaris and
Pseudomonas savastanoe), as well as an antifungal activity (
Aspergilus niger)
[1][41][42][43][44]. Appendino et al. also proved an increased antimicrobial activity for Δ9-THC and CBD on
Staphylococcus aureus [45]. A similar study was carried out by Bass et al., who reported a significant decrease in neurological damage inflicted by infections with
Streptococcus pneumoniae following long-term CBD administration
[46]. Finally, Chakraborty et al. reported the beneficial antimicrobial effect of
Cannabis sativa on methicillin-resistant
Staphylococcus aureus [47]. A high percentage of critically ill patients with sepsis present a complete loss of appetite, as well as a reduction of metabolic activity. Recent studies have shown that by stimulating the CB1 receptor and the central modulation mechanisms, one can obtain a significant increase in voluntary food intake. Bellocchio et al. have shown, in an experimental study, that the administration of titrated doses of CB1 receptor agonists significantly increases appetite. On the other hand, the same group has also proven that by increasing the CB1 receptor agonist doses, there is a sudden decline in feeding in lab animals
[48]. Sardinha et al. carried out a study on the CB2 receptor modulation under conditions of lipopolysachharide (LPS)-induced sepsis. Following this study, they observed a decrease in both the leukocyte expression and endothelial interactions. For this study, the group used different cannabinoids, reporting a decrease in the number of adherent leukocytes induced by the HU-308-CB2 receptor agonist
[49].
Another complex study regarding the modulation of the inflammatory response induced by CB2 receptor activation was carried out by Wang et al. on skin wound healing
[50]. For CB2 activation, they used extremely specific agonists and antagonists for cannabinoid receptors, such as GP1a [1-(2,4-dichlorophenyl)-6-methyl-
N-piperidin-1-yl-4
H-indeno-[1,2-c]pyrazole-3-carboxamide], and AM-630 [6-iodo-2-methyl-1-(2-morpholin-4-ylethyl)indol-3-yl-(4-methoxyphenyl)methanone]. Following the activation of the CB2 receptor, they observed a decrease in the expressions of IL-6, IL-1β, TNF-α, vascular endothelial growth factor (VEGF)-A, stromal cell-derived factor (SDF)-1, and monocyte chemotactic protein (MCP)-1
[50]. Chiurchiu et al., in a study on the AEA endocannabinoid effects on primary human T-lymphocytes, showed a strong immunomodulatory effect on the cellular function. Furthermore, they highlighted a reduction in cell proliferation induced by AEA in this particular context. Last but not least, an important decrease in AEA levels has also been proven in this context, as well as a decrease in IFN-γ and TNF-α from CD8
+ T cells
[5]. Eisenstein et al. also reported a reduction in the primary and secondary antibody production in splenocytes, as well as a decrease for the Th2 activity in human T cells
[51]. Regarding the specific inflammation characteristics for lung injuries, Ribeiro et al., observed a reduction in IL-6, MCP-1, TNF-α, and MIP-2 expressions following the administration of a single dose of CBD (20 mg/kg)
[52].
4. Cannabinoid Signaling System and Redox Activity-Linked with Sepsis
One of the most aggressive molecule structures responsible for the augmentation of inflammation is represented by atoms with a deficit of electronic stability that are called free radicals (FRs). FRs are produced under normal metabolic conditions, with their activation being normally inhibited by a series of endogenous and exogenous antioxidant substrates. In hypermetabolic states like inflammation and sepsis, the oxidant/antioxidant balance is significantly disrupted, with an exaggerated increase in FR expressions. This major imbalance that becomes systemic in a very short period of time is called oxidative stress (OS), and is involved in the majority of biochemical pathways
[53][54][55][56][57][58].
In recent last years, academia has brought great focus on the redox expression in critically ill patients, such as an FR attack, and modified a series of biological systems, such as the mitochondrial DNA, the structure of lipids, the structure and activity of proteins, and the cytoplasmic systems. If we look at the overall context of sepsis, macrophages and neutrophils are the main substrates responsible for increased FR, reactive oxygen species (ROS), and reactive nitrogen species (RNS), affecting important processes such as cellular metabolism, angiogenesis, and the mitochondrial respiratory chain
[59]. Under prooxidative and proinflammatory conditions specific for critically ill patients with sepsis, a disrupted mitochondrial activity leads to major energy deficits, therefore increasing the production of new inflammatory mediators, and in the end leading to clinical complications associated with sepsis
[54][60][61]. This fact becomes extremely important as the mitochondrion represents the “power engine” of the cell, directly responsible for energy production under the form of adenosine triphosphate (ATP). The mitochondrial genome will be affected during the redox stress specific for sepsis, and during increasing the inflammatory response. Structurally, the mitochondrial DNA (mtDNA) contains approximately 16 kilobases, and is responsible for coding certain specific and essential processes involved in the metabolic functions of the cell. Among these is the modulation of 13 genes from the respiratory chain, I, III, IV, and V, as well as 22 genes responsible for the expression of the mitochondrial tRNA
[62][63][64]. mtDNA is extremely vulnerable to redox attack because of the limited DNA repair mechanisms, as well as the lack of adequate histone coating. In this critical situation, the mitochondrial activity is substantially affected by the alteration of the molecular mechanisms, and of the respiratory chain and by the imbalances in the energy pathways
[65]. The specific reactions for redox imbalances are represented especially by the coupling of the superoxide anion with nitrogen monoxide, leading to the production of a series of extremely reactive molecular species, such as peroxynitrite. Other substrates with a strong oxidative character are represented by the hypochlorous acid and hypobromous acid resulting from the oxidant reactions of the hydrogen peroxidase. The attack of these free radicals on the mitochondrial structures first impacts on the respiratory chain and on the electron transport chain, leading to an accelerated increase in membrane permeability through the reduction of its electric potential. As a consequence, numerous fragments resulting from the mtDNA destruction will be released, which will further interact with the immune system. This cascade of molecular interactions will lead to the activation of macrophages, neutrophils, and TLR-cells, with a further augmentation of the inflammatory status and acceleration of the cellular redox stress
[58][59][60][61][63][64][65].
Sun et al. investigated the neuroprotective effects of cannabinoids in the context of oxygen–glucose-deprivation/reperfusion. Following their study, they observed that cannabinoids have direct implications on minimizing OS and on reestablishing mitochondrial and glucose metabolism through the pentose-phosphate pathways
[66]. Another effect on the molecular mechanisms of cannabinoids is represented by the reduction of the NO synthase protein (iNOS) activity, and by the inhibition of the TNF-α activity. Furthermore, they reported their involvement in the excessive transmembrane Ca
2+, as well as in minimizing the enzymatic hydrolysis in the redox context. Burstein et al. reported a decrease in redox activity due to hypoxia and ischemia in newborns
[67]. Carrier et al., in a similar study, showed an increase in the adenosine levels in the brain, leading to reduced inflammation-related neurotoxicity
[68]. Castillo et al., after carrying out a similar study, reported a significant decrease in hypoxia-induced brain damage phenomena. The main segments involved are represented by the reduction in the expression of IL-6, TNF-α, and iNOS
[69]. Moreover, they have shown that the neuroprotective mechanisms are mediated by the CB2 receptors and by adenosine through the A2A receptor
[69]. Even more interesting is the fact that increased levels for CB1 receptors have been reported on the surface of the mitochondrial coating. Hebert-Chatelain et al. showed that CB1 receptors interact with the respiratory chain complex I, which is responsible for the modulation of the redox activity in this segment
[70]. Palomba et al. carried out a study on CB1 receptors and on the redox modulation capacity in the arcuate nucleus neurons, and its interaction with the appetite-regulating hypothalamic neuropeptides. The group observed a decrease in leptin-mediated processes, with a significant reduction in the ROS production in hypothalamic neurons cellular lines. Furthermore, they observed that in these redox processes, blocking mechanisms intervene in the activation of catalase and peroxisome proliferator-activated receptors gamma (PPAR-γ)
[27]. Under the critical conditions expressed in sepsis, due to accentuated inflammation, and because of accelerated redox processes, there is an overexpression of neuronal inflammation, with a negative impact on the molecular and metabolic mechanisms, as well as on the clinical prognosis and post-ICU recovery. An intensely studied mechanism is represented by neuronal Ca
2+ homeostasis, and by the neuronal metabolic activation. In this situation, it was proven that both biological regulation pathways are modulated by the mitochondria, as it plays a crucial role in the neuronal integrity under the conditions of an accelerated Ca
2+ dynamic. Recent studies have highlighted a series of mechanisms responsible for CBD induced Ca
2+ homeostasis. Ryan et al. carried out a complex study regarding CBD action mechanisms in Ca
2+ homeostasis in neuronal preparation. They observed a series of neuroprotective properties of CBD in neuroblastoma cell lines treated with mitochondrial toxins. Moreover, they showed an important decrease in redox activity and an adequate modulation of Ca
2+ homeostasis
[71].
 +1 credit
+1 credit