Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Alanah Sheridan | -- | 4397 | 2022-06-07 14:10:04 | | | |
| 2 | Vivi Li | + 2005 word(s) | 6402 | 2022-06-08 03:46:01 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Sheridan, A.; Gage, M.; Wheeler-Jones, C. The Immunomodulatory Effects of Statins on Macrophages. Encyclopedia. Available online: https://encyclopedia.pub/entry/23787 (accessed on 13 June 2026).
Sheridan A, Gage M, Wheeler-Jones C. The Immunomodulatory Effects of Statins on Macrophages. Encyclopedia. Available at: https://encyclopedia.pub/entry/23787. Accessed June 13, 2026.
Sheridan, Alanah, Matthew Gage, Caroline Wheeler-Jones. "The Immunomodulatory Effects of Statins on Macrophages" Encyclopedia, https://encyclopedia.pub/entry/23787 (accessed June 13, 2026).
Sheridan, A., Gage, M., & Wheeler-Jones, C. (2022, June 07). The Immunomodulatory Effects of Statins on Macrophages. In Encyclopedia. https://encyclopedia.pub/entry/23787
Sheridan, Alanah, et al. "The Immunomodulatory Effects of Statins on Macrophages." Encyclopedia. Web. 07 June, 2022.
Copy Citation
Statins are 3-hydroxy-3-methylglutaryl coenzyme A (HMG-CoA) reductase inhibitors used worldwide to manage dyslipidaemia and thus limit the development of atherosclerotic disease and its complications. These atheroprotective drugs are now known to exert pleiotropic actions outside of their cholesterol-lowering activity, including altering immune cell function. Macrophages are phagocytic leukocytes that play critical functional roles in the pathogenesis of atherosclerosis and are directly targeted by statins. Early studies documented the anti-inflammatory effects of statins on macrophages, but emerging evidence suggests that these drugs can also enhance pro-inflammatory macrophage responses, creating an unresolved paradox.
statins
macrophages
atherosclerosis
inflammation
cholesterol
atorvastatin
simvastatin
rosuvastatin
fluvastatin
lovastatin
pitavastatin
cerivastatin
metavastatin
pravastatin
1. Introduction
1.1. Statins Are the Most Widely Prescribed Medications for the Prevention of Cardiovascular Disease
Cardiovascular disease (CVD) is the leading cause of mortality worldwide, accounting for an estimated 17.9 million deaths in 2019 [1], which equates to 32% of all global deaths. Atherosclerosis, a chronic inflammatory disease characterised by a narrowing of the arteries, is the main underlying cause of CVD [2] and is driven by an imbalance in lipid metabolism and a maladaptive immune response [3]. Despite its causal role in deaths globally, CVD-related mortality in the UK and other industrialised countries has declined over the last 40 years [4][5], and statins, which have revolutionized the prevention of atherosclerotic CVD, have significantly contributed to this change [6]. The efficacy of statins in the preventative treatment of CVD has led to them becoming one of the most prescribed medications worldwide, with over 200 million people taking them [7].
The clinical benefit of statins in CVD prevention is thought to be primarily driven by their lipid-lowering effects [8][9], as epidemiological studies have revealed high plasma levels of low-density lipoprotein cholesterol (LDL-C) to be a significant risk factor for atherosclerosis [10]. Mechanistically, statins inhibit cellular cholesterol biosynthesis through the inhibition of the mevalonate pathway via the rate-limiting enzyme 3-hydroxy-3-methylglutaryl coenzyme-A (HMG-CoA) reductase. In addition, statins upregulate hepatic low-density lipoprotein receptor transcription, increasing blood LDL-C removal. Together, these factors result in a 20–60% reduction in circulating LDL-C depending on the particular statin type and dose administered (Table 1).
Table 1. Pharmacokinetic properties of different statins.
| Statin Name | Brand Name | Daily Dose (mg) | Effect on LDL Cholesterol (% Decrease) | Lipophilicity | Marketed Drug Form | Half-Life (h) | Primary Metabolizing Enzyme(s) |
|---|---|---|---|---|---|---|---|
| Atorvastatin | Lipitor | 10–80 [11] | 37–55 [12][13] | Lipophilic [14] | Acid [14] | 14 [11][14] | CYP3A4 [14] |
| Cerivastatin a | Baycol | 0.02–0.8 [15] | 12–42 [16] | Lipophilic [14] | Acid [14] | 2–4 [15] | CYP3A4, 2C8 [14][15] |
| Fluvastatin | Lescol | 20–80 [17] | 21–33 [12][13] | Lipophilic [14] | Acid [14] | 3 [17] | CYP2C9 [14][17] |
| Lovastatin | Mevacor | 10–80 [18] | 21–45 [12] | Lipophilic [14] | Lactone [14] | 3 [14] | CYP3A4 [14][18] |
| Metavastatin b | Lipophilic [19] | ||||||
| Pitavastatin | Livalo | 1–4 [20] | 33–44 [21] | Lipophilic [14] | Acid [14] | 12 [20] | CYP2C8, 2C9 [14][20] |
| Pravastatin | Pravachol | 10–80 [22] | 20–33 [12] | Hydrophilic [14] | Acid [14] | 1.8 [14][22] | Non-CYP [14] |
| Rosuvastatin | Crestor | 5–40 [23] | 38–53 [12][13] | Hydrophilic [14] | Acid [14] | 19 [14][23] | CYP2C9 [23] |
| Simvastatin | Zocor | 5–80 [24] | 23–42 [12][13] | Lipophilic [14] | Lactone [14] | 2 [14] | CYP3A4 [14][24] |
There are several statins clinically available, with atorvastatin, simvastatin, and rosuvastatin being the most popular and most widely prescribed [27][28]. The different statins vary in their lipophilicity, metabolism, elimination half-lives, and potency and evidence suggests that these distinct characteristics may lead to differential effects on their efficacy (Table 1). For example, studies have suggested that the variability in different statins’ solubility affects their ability to enter cells, with lipophilic statins being found to passively diffuse into numerous cell types, whilst hydrophilic statins are hypothesized to be more liver-selective due to their dependence on membrane transporters [29][30]. These different properties have been suggested to potentially result in varying distributions of the drugs in different tissues, thereby resulting in differential effects on the mevalonate pathway [14].
1.2. The Central Role of Macrophages in Inflammation and CVD
Atherosclerosis is recognised as a chronic inflammatory disease characterised by a lipid imbalance and maladaptive inflammation exacerbated by the accumulation of inflammatory cells in the arterial wall. Cholesterol-laden macrophages (known as foam cells) are protagonists in the development and progression of atherosclerosis, making up the main immune cellular constituents of atherosclerotic lesions [31][32]. Foam cells contribute to the maintenance of the local endothelial inflammatory response by secreting proinflammatory cytokines and chemokines, as well as producing reactive oxygen and nitrogen species. Macrophages also engage in crosstalk with vascular smooth muscle cells, amplifying the inflammatory cycle by producing additional proinflammatory signals, promoting the growth of lipid-rich lesions [33]. Over time, these lesions can undergo further remodelling and form a fibrous cap, a layer of connective tissue that shields the lesion from the lumen (together with the lesion, this is known as an atherosclerotic plaque) [34]. Plaques can become unstable and rupture unexpectedly, exposing the lipid core to the blood and triggering thrombosis, which can result in partial or complete vessel occlusion and culminate in myocardial infarction, stroke, and other ischemic events.
Macrophages are tissue-resident leukocytes present in virtually all tissues of the body and have diverse roles, acting as both pro and anti-inflammatory mediators and being associated with the resolution of infections, tissue development, homeostasis, repair, and remodelling [35]. Macrophages display remarkable plasticity, which is shaped by their specific microenvironment [36]. Following their differentiation from monocytes, macrophages are often classified into one of two distinct functional polarization states (based on surface expression markers), M1, classically activated, or M2, alternatively activated [37]. These states represent the two extremes of a spectrum of macrophage phenotypes, describing a pro-inflammatory and anti-inflammatory/pro-resolving phenotype, respectively. Additionally, M0 is used to denote resting/non-activated cells.
M1-like activated macrophages are induced by microbial products, such as lipopolysaccharides (LPS) and toll-like receptor (TLR) ligands, or by cytokines secreted from other immune cells, such as interferon (IFN)-gamma (IFN-γ) [38] (Figure 1). These inflammatory signals trigger both transmembrane receptors (e.g., TLRs and IFN-γ receptor (IFN-γR)) and cytoplasmic receptors (e.g., nucleotide-binding and oligomerization domain (NOD)-like receptors (NLRs)). Traditionally, M1-like macrophages are functionally associated with pathogen clearance and antigen presentation to T cells to initiate the adaptive immune response, which they achieve by secreting high levels of pro-inflammatory cytokines, such as tumour necrosis factor-alpha (TNFα), interleukin (IL) 1β (IL-1β), IL-6, and IL-12, and by expressing activation markers including cluster of differentiation (CD)80, CD86, class II transactivator (CIITA), and major histocompatibility complex class II receptor (MHC-II). Pro-inflammatory macrophages also express high levels of inducible nitric oxide synthase, which enables the synthesis of nitric oxide (NO) that can, in turn, form reactive oxygen species (ROS) with microbicidal properties. The expression of these inflammatory mediators is predominantly controlled by the activation and nuclear translocation of transcription factors in response to initial receptor recognition of inflammatory stimuli. NF-κB (nuclear factor kappa-light-chain enhancer of B-cell) [39], together with STAT1 (Signal transducer and activator of transcription) [38], STAT3 [40], IRF (IFN-γ regulatory factor) [41], and AP-1 (activator protein 1) [42] are all associated with the polarization of macrophages to an M1-like phenotype.
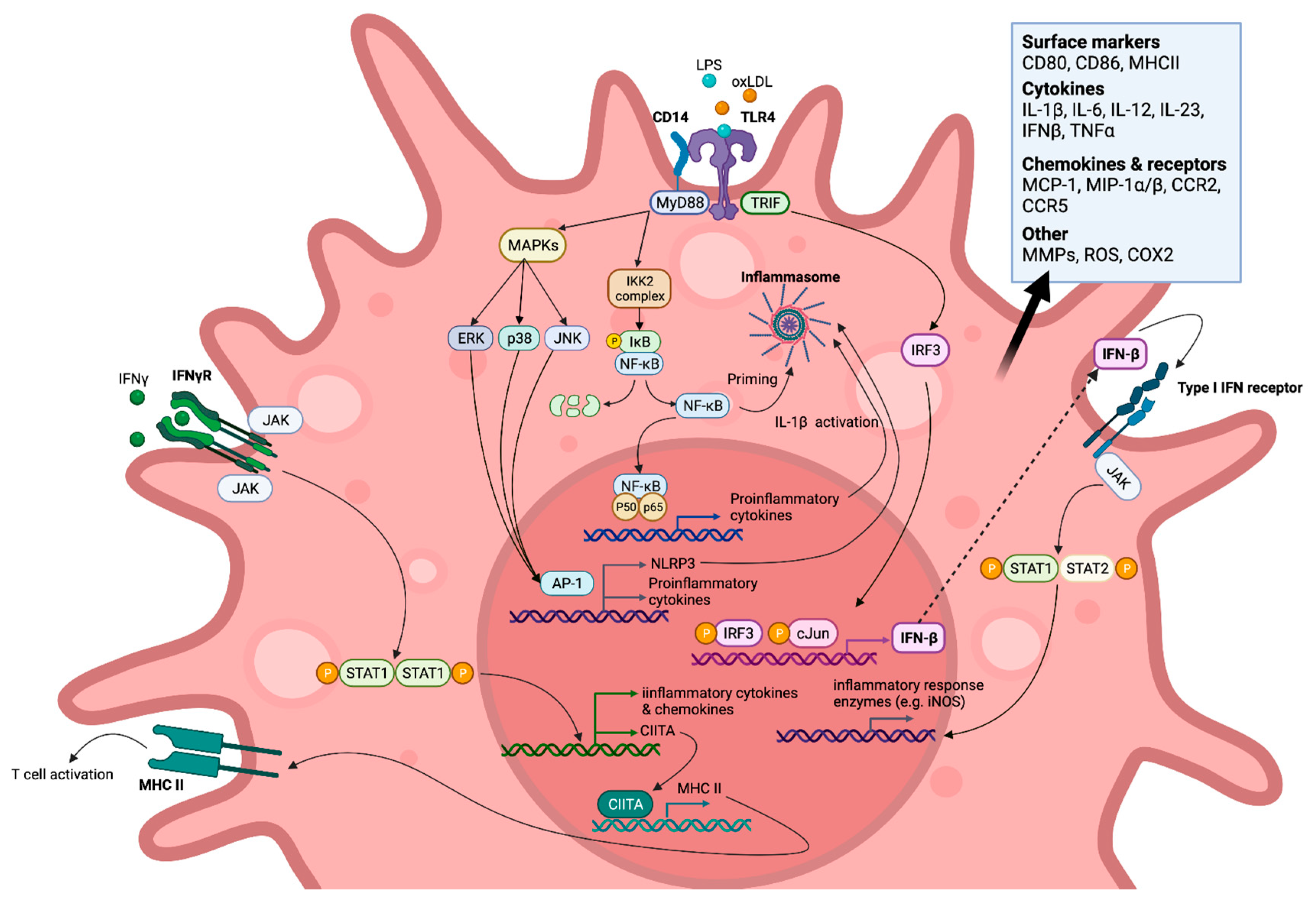
Figure 1. M1-like polarised macrophage signalling pathways (simplified) induced by toll-like receptor (TLR) and IFN-γ receptor (IFN-γR) endogenous and exogenous agonists. Created with BioRender.com, accessed on 7 March 2022.
The switch to M2-like, or alternatively activated, macrophages is mediated by factors such as IL-4 and IL-13 released from innate and adaptive immune cells [38]. M2-like macrophages are considered to be anti-inflammatory as they are noted to resolve inflammation and stimulate tissue repair. They exhibit increased expression of pro-inflammatory cytokine decoy and scavenger receptors, such as IL-1R [43], which act as molecular traps, preventing canonical signalling and thereby regulating inflammation. In addition, they secrete high levels of IL-10, transforming growth factor β, and vascular endothelial growth factor, which ameliorate the excessive activity of both innate and adaptive immune cells, stimulate fibroblast and endothelial cell proliferation, and promote blood-vessel development, allowing wound healing [38][44]. M2 polarization is also characterised by the expression of the transcription factors STAT6, SOCS1 (suppressor of cytokine signalling), and PPARγ (peroxisome proliferator-activated receptor gamma), along with the markers CD163 and CD36.
Recent evidence suggests that the M1/M2 classification system greatly oversimplifies macrophage heterogeneity. Instead, research indicates that macrophages exist on an activation spectrum with a wide array of phenotypes between these M1 and M2 extremes, dependent on their exposure to biochemical stimuli. Researchers refer the reader to recent reviews [45][46][47] for detailed discussion. Despite the evolving views of macrophage polarization, to better compare the findings of the literature referenced in this entry, the simplified M1/M2 nomenclature will be used as appropriate.
Atherosclerotic lesions house a heterogeneous population of macrophages, although M1-like cells are the predominant sub-type [48][49]. M1-like macrophages, expressing pro-inflammatory markers, are known to be associated with unstable and rupture-prone areas, whilst M2-like macrophages are found in stable regions [48]. M2-like macrophages have also been implicated in plaque regression in several different models suggesting that this polarization state’s enrichment may aid the resolution of atherosclerosis [50][51][52]. Therefore, therapeutic agents that encourage this switch from an M1 to an M2-like state, suppressing inflammation, could be a promising treatment strategy to reduce cardiovascular events [53]. Macrophages also play a central role in many other disease states and have therefore emerged as important therapeutic targets in several other pathologies, such as the development and progression of cancerous tumours [54], autoimmune disorders [55] and sepsis [56].
2. In Vitro Evidence Demonstrating the Direct Effects of Statins on Macrophages
An abundance of in vitro studies have reported paradoxical statin-mediated effects on inflammation (Table 2, Figure 2 and Figure 3), resulting from either blunting or enhancing pro-inflammatory signalling cascades. However, a limited number of studies have also reported that statins may alter the differentiation of macrophages rather than simply acting as regulators of inflammatory signalling pathways.
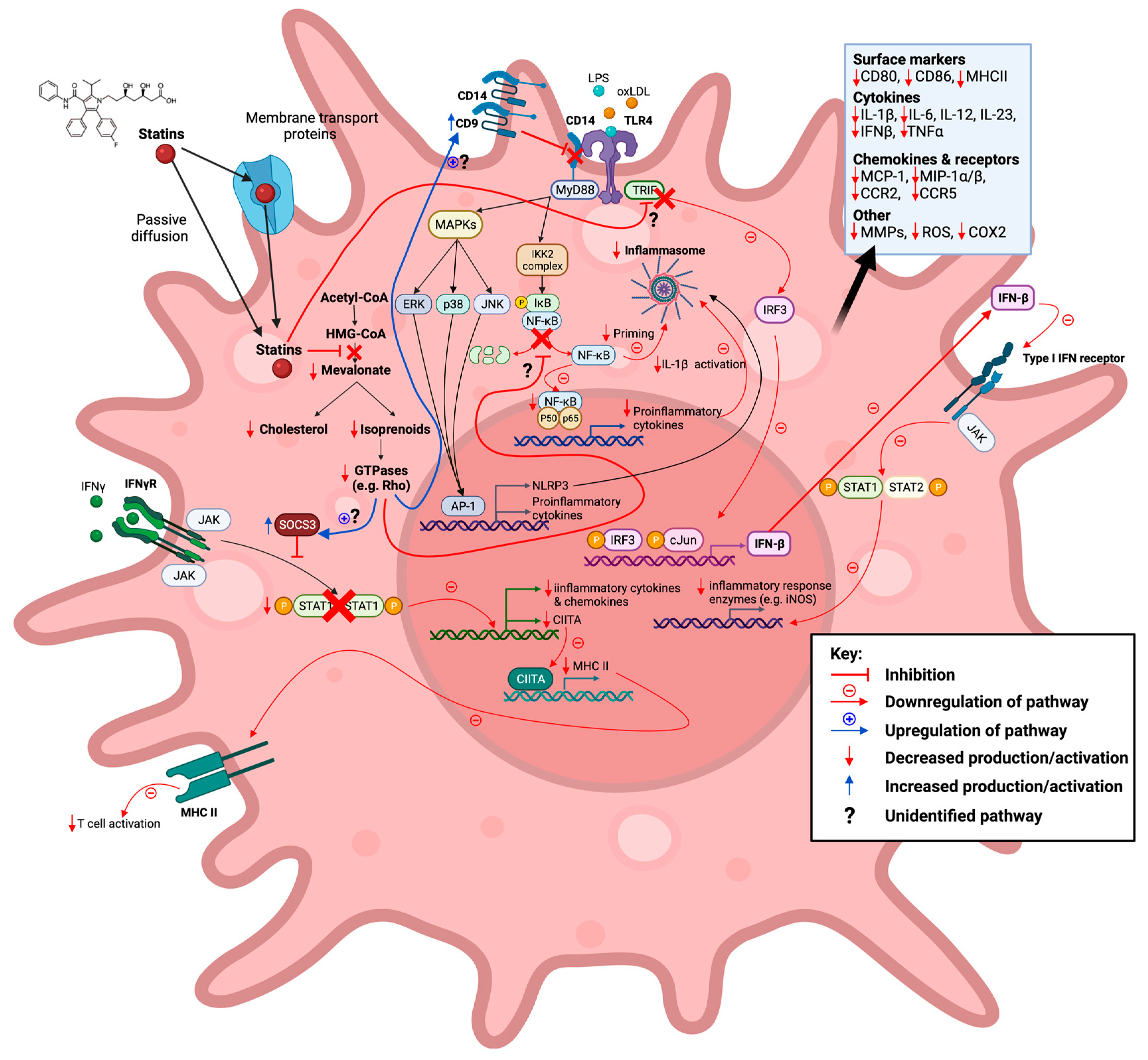
Figure 2. Statins inhibit the mevalonate pathway leading to both reduced cholesterol and isoprenoid biosynthesis, thereby also blocking farnesylation and geranylgeranylation of GTPases. Reduction in these downstream mevalonate intermediates is demonstrated to affect M1-associated macrophage inflammatory signalling pathways in vitro in an anti-inflammatory manner. This action of statins is seen in response to exogenous lipopolysaccharide (LPS), endogenous (interferon gamma (IFN-γ), and oxidized low-density lipoprotein (oxLDL) ligands. Created with BioRender.com, accessed on 7 March 2022.
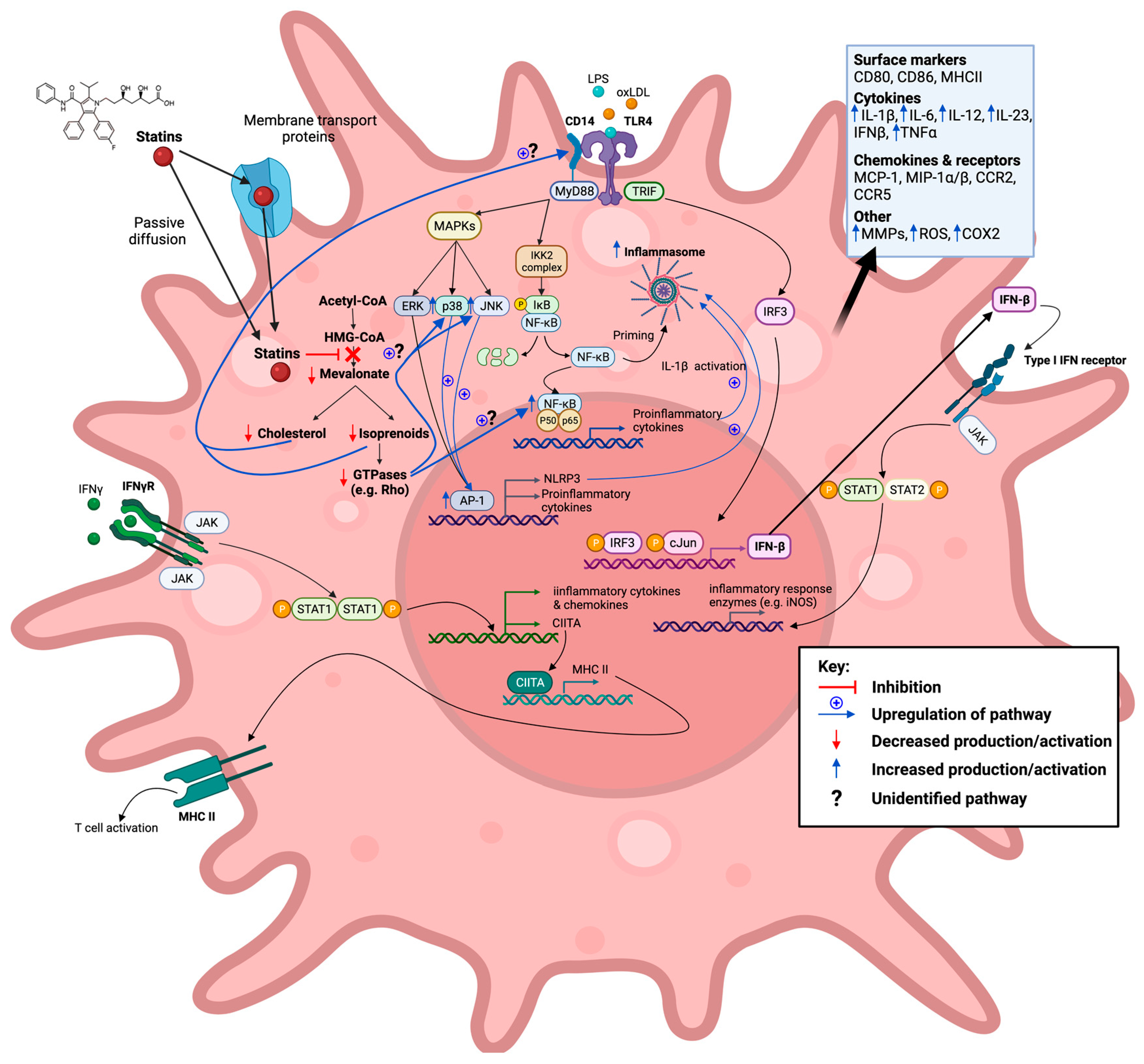
Figure 3. Statins inhibit the mevalonate pathway leading to both reduced cholesterol and isoprenoid biosynthesis, thereby also blocking farnesylation and geranylgeranylation of GTPases. Reduction in these downstream mevalonate intermediates is demonstrated to affect M1-associated macrophage inflammatory signalling pathways in vitro in a pro-inflammatory manner. This action of statins is seen in response to exogenous lipopolysaccharide (LPS) and oxidized low-density lipoprotein (oxLDL) ligands. Created with BioRender.com, accessed on 7 March 2022.
Table 2. The effects of statins on macrophages in vitro. Abbreviations: PBMC, peripheral blood mononuclear cells; BMDMs, bone-marrow derived macrophages; iNOS, inducible nitric oxide synthase; COX-2, cyclooxygenase-2; uPAR, urokinase plasminogen activator receptor; AdipoR, adiponectin receptors; mTOR, mechanistic target of rapamycin; mRNA, messenger ribonucleic acid; MWCNT, multi-walled carbon nanotubes; TF, tissue factor; GILZ, glucocorticoid-induced leucine zipper; TPA, 12-O-tetradecanoyl-phorbol-13-acetate; PMA, phorbol myristate acetate; acLDL, acetylated LDL; M-CSF, macrophage colony-stimulating factor; CC, cholesterol crystal; MSU, monosodium urate; SOD1, superoxide dismutase-1; AGE-RAGE, advanced glycation endproducts-receptor for advanced glycation endproducts; C/EBP, CCAAT/enhancer binding proteins; IP-10, interferon gamma-induced protein-10; agLDL, aggregated LDL; ETS-1, erythroblast transformation specific-1; KLF-2, Krüppel-like factor-2; ICAM-1, intercellular adhesion molecule-1.
| Statin | Model | Summary | Inflammatory Effect |
Ref. | |
|---|---|---|---|---|---|
| Pro | Anti | ||||
| Atorvastatin | Human PBMC derived macrophages | Statins acted as inhibitors of the induction of MHC-II expression by IFN-γ due to suppression of CIITA transcription. Statins repressed MHC-II mediated T-cell activation. | ✓ | [57] | |
| Primary macrophages from B10.PL mice | Atorvastatin prevented IFN-γ induced MHC-II, CD40, CD80, and CD86 expression. | ✓ | [58] | ||
| RAW 264.7 macrophages | Atorvastatin inhibited LPS and IFN-γ-induced NO formation and iNOS induction—thought to be mediated through suppression of NF-κB activation and IFN-γ through STAT1. | ✓ | [59] | ||
| Murine peritoneal macrophages | Atorvastatin pretreatment enhanced TLR2 and TLR4 ligand-stimulated IL-6 and TNF production. | ✓ | [60] | ||
| RAW 264.7 macrophages | Enhanced LPS-mediated MMP-9 gene expression. | ✓ | [61] | ||
| RAW 264.7 macrophages | Atorvastatin pretreatment inhibited oxLDL-induced increase in COX-2, TNFα, and MCP-1 secretion. | ✓ | [62] | ||
| Murine BMDMs | Atorvastatin pretreatment exacerbated LPS-induced upregulation of Il-1b, IL-6, and NLRP3 transcript levels. | ✓ | [63] | ||
| Human PBMC derived macrophages | Statin treatment in combination with IL-4 during the macrophage differentiation phase led to increased M2 polarization via PPARγ activation. | ✓ | [64] | ||
| RAW 264.7 macrophages | Atorvastatin pretreatment inhibited LPS-induced IL-1β and TNFα production in RAW 264.7 macrophages through the enhancement of autophagy. Statin treatment was seen to attenuate NLRP3 inflammasome induction in response to LPS stimulation. Atorvastatin pretreatment inhibited the expression of IL-1β in response to LPS stimulation in peritoneal murine macrophages through autophagy activation, but not that of TNFα. | ✓ | [65] | ||
| Human PBMC derived macrophages | Atorvastatin reduced matrix degradation capability via reduced MMP-14 activation and uPAR localization to filipodia in LPS and IFN-γ stimulated macrophages. | ✓ | [66] | ||
| RAW 264.7 macrophages and J774 macrophages | Atorvastatin increased Rac1 GTP-loading in LPS stimulated macrophages, enhancing production of the proinflammatory cytokines IL-1β, TNFα, and IL-6. | ✓ | [67] | ||
| Human monocyte derived macrophages | Statin treatment during macrophage differentiation phase led to enhanced LPS-induced IL-1β and IL-6 secretion. | ✓ | [68] | ||
| THP1 derived macrophages | Statin treatment led to increased pro-inflammatory cytokine (IL-1β, TNFα, and IL-6) and AdipoR expression (also seen in combination with oxLDL stimulation); 24 h statin treatment resulted in increased IL-10 mRNA levels, whilst 72 h treatment resulted in decreased expression. | ✓ | [69] | ||
| Murine BMDMs | Statin-treated macrophages exhibited increased LPS-induced activation of NF-κB and IL-1β protein secretion in response to inflammasome stimulation. | ✓ | ✓ | [70] | |
| Murine BMDMs | Statin pretreatment exacerbated LPS-induced upregulation of IL-1β and NLRP3 transcript levels via p38 and mTOR. | ✓ | [71] | ||
| THP1 derived macrophages | Impaired MWCNT-elicited IL-1β secretion. | ✓ | [72] | ||
| Cerivastatin | Human PBMC derived macrophages | Cerivastatin treatment suppressed growth of macrophages expressing MMPs and TFs. | ✓ | [73] | |
| Rabbit foamy macrophages | Decreased protein expression and activity of MMP-1, MMP-2, and MMP-9. | ✓ | [74] | ||
| RAW-Blue™ cells and Murine BMDMs | Cerivastatin increased NF-κB/AP-1 activation in unstimulated and LPS-activated macrophages. LPS-induced TNF, IL-1β, and IL-6 expression was amplified. Expression of arginase-1 and GILZ was enhanced in unstimulated, LPS- and IL-4-activated macrophages. | ✓ | ✓ | [75] | |
| Fluvastatin | human PBMC derived macrophages | Fluvastatin decreased TF activity in both unstimulated and LPS-, or ac-LDL-stimulated macrophages, but enhanced IL-1β cytokine release. | ✓ | ✓ | [76] |
| Murine peritoneal macrophages and human PBMC derived macrophages | Simvastatin decreased MMP-9 protein secretion and inhibited TPA-induced enhanced MMP-9 release. | ✓ | [77] | ||
| RAW 264.7 macrophages | Fluvastatin inhibited LPS and IFN-γ-induced NO formation and iNOS induction.Thought to be mediated through suppression of NF-κB activation and IFN-γ through STAT1. | ✓ | [59] | ||
| RAW 264.7 macrophages | Fluvastatin upregulated macrophage Socs3 expression, resulting in low responsiveness to inflammatory signals (IFN-γ, IL-6, and M-CSF) due to lower activation of STAT1, STAT3, and STAT5. | ✓ | [78] | ||
| THP1 derived macrophages and THP1 derived acLDL loaded macrophages | Fluvastatin reduced both the expression, secretion, and proportion of active MMP-9 in PMA stimulated and acLDL-loaded THP1 derived macrophages. | ✓ | [79] | ||
| RAW 264.7 macrophages and murine BMDMs | Fluvastatin inhibited LPS-induced suppression of CD9, leading to reduced formation of CD14/TLR4 complexes and TNFα and MMP-9 release. | ✓ | [80] | ||
| Murine BMDMs | Fluvastatin pre-treatment exacerbated LPS-induced upregulation of IL-1b, IL-6, and NLRP3 transcript levels. Statin and LPS treatment of BMDMs harvested from NLRP3−/− mice synergistically enhanced IL-6 but did not affect IL-1β secretion. Statin treatment alone had no effect on the production of inflammatory mediators. | ✓ | [63] | ||
| Human monocyte derived macrophages | Statin treatment during macrophage differentiation phase led to enhanced LPS-induced IL-1β and IL-6 secretion. | ✓ | [68] | ||
| Murine BMDMs | Statin pretreatment exacerbated LPS-induced upregulation of IL-1b and NLRP3 transcript levels via p38 and mTOR. | ✓ | [71] | ||
| THP1 derived macrophages | Impaired MWCNT-elicited IL-1β secretion. | ✓ | [72] | ||
| Human PBMC derived macrophages | Decreased the activity of iNOS in M1 macrophages. | ✓ | [81] | ||
| Lovastatin | Rat peritoneal macrophages and microglia | Inhibited LPS-induced production of NO, TNFα, IL-1β, and IL-6 in rat primary microglia and macrophages. | ✓ | [82] | |
| Human PBMC derived macrophages | Statins acted as inhibitors of the induction of MHC-II expression by IFN-γ due to suppression of CIITA transcription. Statins repressed MHC-II mediated T-cell activation. | ✓ | [57] | ||
| RAW 264.7 macrophages | Lovastatin inhibited LPS and IFN-γ-induced NO formation and iNOS induction—thought to be mediated through suppression of NF-κB activation and IFN-γ through STAT1. | ✓ | [59] | ||
| RAW 264.7 macrophages | Lovastatin upregulated macrophage Socs3 expression, resulting in low responsiveness to inflammatory signals (IFN-γ, IL-6, and M-CSF) due to lower activation of STAT1, STAT3, and STAT5. | ✓ | [78] | ||
| Rabbit foamy macrophages | Decreased protein expression and activity of MMP-1, MMP-2, and MMP-9. | ✓ | [74] | ||
| RAW 264.7 macrophages | Lovastatin increased LPS-induced TNFα production. | ✓ | [83] | ||
| P388D1 macrophages | Statins increased production of MMP-12 in activated macrophage. | ✓ | [84] | ||
| RAW 264.7 macrophages | Lovastatin increased CD14 expression and enhanced LPS-induced membrane levels leading to greater TNFα production, but simultaneously suppressed soluble CD14. | ✓ | [85] | ||
| BMDMs from C57BL/6J mice and RAW 264.7 macrophages | Lovastatin blocked IFN-γ-induced Citta gene expression by inhibiting transcriptional events at Citta pIV, thereby suppressing MHC-II expression. | ✓ | [86] | ||
| RAW 264.7 macrophages | Lovastatin treatment induced NO release but did not affect pro-inflammatory cytokine levels in unstimulated cells. However, with LPS it synergistically enhanced IL-6, IL-12p40, IL-1β, and NO release. | ✓ | [87] | ||
| Murine BMDMs | Lovastatin pretreatment exacerbated LPS-induced upregulation of IL-1b, IL-6, and NLRP3 transcript levels. | ✓ | [63] | ||
| THP1 derived macrophages | Impaired MWCNT-elicited IL-1β secretion. | ✓ | [72] | ||
| Metavastatin | P388D1 cell line | Statins increased production of MMP-12 in activated macrophages. | ✓ | [84] | |
| U937 derived macrophages and RAW 264.7 macrophages | Metavastatin pretreatment significantly increased bacterial clearance, despite reducing oxidative burst and phagocytosis due to increased induction of extracellular traps. | ✓ | ✓ | [88] | |
| J774A.1 mouse macrophages | Increased levels of iNOS and killing of internalized S. pneumoniae. | ✓ | [89] | ||
| Pitavastatin | RAW 264.7 macrophages | Suppressed LPS-induced upregulation of MCP-1, iNOS, and IL-6 gene expression. | ✓ | [90] | |
| THP1 derived macrophages, and murine peritoneal macrophages and BMDMs (BALB/cCrSlc mice) | Pravastatin repressed mature IL-1β release elicited by MWCNT/CC/MSU exposure in THP1-derived macrophages, and LPS + MWCNT induced mature IL-1β release in peritoneal macrophages. Pravastatin pretreatment strongly enhanced mature IL-1β release in LPS + MWCNT exposed BMDMs. | ✓ | ✓ | [72] | |
| Pravastatin | Human PBMC derived macrophages | Statins acted as inhibitors of the induction of MHC-II expression by IFN-γ due to suppression of CIITA transcription. Statins repressed MHC-II mediated T-cell activation. | ✓ | [57] | |
| RAW 264.7 macrophages | Pravastatin inhibited LPS and IFN-γ-induced NO formation and iNOS induction—thought to be mediated through suppression of NF-κB activation and IFN-γ through STAT1. | ✓ | [59] | ||
| RAW 264.7 macrophages | Pravastatin upregulated macrophage Socs3 expression, resulting in low responsiveness to inflammatory signals (IFN-γ, IL-6, and M-CSF) due to lower activation of STAT1, STAT3, and STAT5. | ✓ | [78] | ||
| RAW 264.7 macrophages | Suppressed LPS-induced upregulation of MCP-1, iNOS, and IL-6 gene expression. | ✓ | [90] | ||
| Rosuvastatin | Human monocyte derived macrophages | Rosuvastatin reduced MMP-7 and MMP-9 production. | ✓ | [91] | |
| oxLDL induced THP1 foam cells | Rosuvastatin inhibited ox-LDL-induced reduction of SOD1 expression. | ✓ | [92] | ||
| THP1 derived macrophages | Rosuvastatin inhibited the AGE-RAGE axis and ROS production. | ✓ | [93] | ||
| RAW 264.7 macrophages and J774 macrophages | Rosuvastatin increased Rac1 GTP-loading in LPS-stimulated macrophages, enhancing production of the proinflammatory cytokines IL-1β, TNFα, and IL-6. | ✓ | [67] | ||
| Human monocyte derived macrophages | Statin treatment during macrophage differentiation phase led to enhanced LPS-induced IL-1β and IL-6 secretion | ✓ | [68] | ||
| THP1 derived macrophages | Statin treatment led to increased pro-inflammatory cytokine (IL-1β, TNFα, and IL-6) and AdipoR expression (also seen in combination with oxLDL stimulation); 24 h statin treatment resulted in increased IL-10 mRNA levels, whilst 72 h treatment resulted in decreased expression. | ✓ | [69] | ||
| THP1 derived macrophages | Inhibited foam cell formation and lessened the secretion of inflammatory cytokines (e.g., TNFα, IL-1β, and IL-6) from oxLDL-treated macrophages | ✓ | [94] | ||
| Simvastatin | Human monocyte derived macrophages | Simvastatin decreased superoxide production and therefore LDL oxidation | ✓ | [95] | |
| human PBMC derived macrophages | Simvastatin decreased TF activity in both unstimulated and LPS-stimulated/ac-LDL-stimulated macrophages. The suppression of TF activity induced by statin treatment was accompanied by a diminution in TF mRNA expression. | ✓ | [76] | ||
| Murine peritoneal macrophages | Simvastatin decreased MMP-9 protein secretion and inhibited TPA-induced enhanced MMP-9 release. | ✓ | [77] | ||
| Rabbit foamy macrophages | Decreased protein expression and activity of MMP-1, MMP-2, and MMP-9. | ✓ | [74] | ||
| Peritoneal murine macrophages and RAW 264.7 macrophages | Simvastatin pretreatment enhanced both IL-12p40 and TNFα LPS-induced mRNA expression and protein production by a mechanism involving the AP-1 and C/EBP transcription factors, but IP-10 levels were reduced. | ✓ | ✓ | [96] | |
| PBMC derived human macrophages | Simvastatin inhibited IFN-γ-induced upregulated mRNA expression of the chemokines MCP-1, MIP-1a, and MIP-1b and the chemokine receptors CCR1, CCR2, and CCR5. MCP-1 protein expression was also notably reduced. | ✓ | [97] | ||
| human primary monocyte derived macrophages | Statin administration significantly increased the secretion of IL-1β but had no significant effect on IL-8 or IL-6 and inhibited the secretion of TNFα. In combination with agLDL loading, statin treatment enhanced secretion of IL-1β and IL-8, but had no effect on TNFα or IL-6 secretion. | ✓ | ✓ | [98] | |
| BMDMs from C57BL/6J mice and RAW 264.7 macrophages | Simvastatin blocked IFN-γ-induced Citta gene expression by inhibiting transcriptional events at Citta pIV, thereby suppressing MHC-II expression. | ✓ | [86] | ||
| PBMC derived human macrophages and THP1 derived macrophages | Simvastatin treatment led to the downregulation of inflammatory signalling pathways, marked by a reduction in the gene expression of proinflammatory associated chemokines (MCP-1, MIP-1, and tissue factor) and transcription factors (NF-κB and ETS-1). The anti-inflammatory associated transcription factor KLF-2 had upregulated gene and protein expression. | ✓ | [99] | ||
| Murine peritoneal macrophages | Simvastatin pretreatment enhanced TLR2 and TLR4 ligand-stimulated IL-6 and TNF production. | ✓ | [60] | ||
| RAW 264.7 macrophages | Enhanced LPS-mediated MMP-9 gene expression. | ✓ | [61] | ||
| PBMC derived human macrophages, HL-60 derived macrophages and murine peritoneal macrophages (treated with simvastatin in vivo) | Simvastatin reduced phagocytosis and oxidative burst of IgG opsonized bacteria but enhanced the production of inflammatory mediators (TNFα and COX-2). No effect was seen on inflammatory mediators in response to non-opsonized bacteria, but impairment of phagocytosis remained. | ✓ | ✓ | [100] | |
| RAW 264.7 macrophages | Simvastatin pretreatment reduced basal and S. aureus-stimulated levels of C5aR and dampened macrophage sensitivity to membrane vesicles released from infected cells, decreasing TNFα production. | ✓ | [101] | ||
| RAW 264.7 macrophages and murine BMDMs | Simvastatin inhibited LPS induced suppression of CD9, leading to reduced formation of CD14/TLR4 complexes and TNFα and MMP-9 release. | ✓ | [80] | ||
| RAW 264.7 macrophages and murine BMDMs | Simvastatin pretreatment enhanced IL-12p40 and TNFα production in IFN-γ and L. monocytogenes stimulated macrophages. Statins suppressed MHC-II surface expression on IFN-γ-activated macrophages | ✓ | ✓ | [102] | |
| THP1 derived macrophages | Simvastatin pretreatment inhibited IFN-γ induced expression of MCP-1 and ICAM-1. | ✓ | [103] | ||
| Murine BMDMs and human PBMCs | Simvastatin enhanced LPS-stimulated pro-IL-1β (28 kDa form), which disrupted mature IL-1β inflammatory actions. | ✓ | [104] | ||
| Murine BMDMs | Simvastatin pretreatment exacerbated LPS-induced upregulation of IL-1b, IL-6, and NLRP3 transcript levels. | ✓ | [63] | ||
| Murine BMDMs | Simvastatin reduced parasite burden by enhancing oxidative burst and phagosome maturation. | ✓ | [105] | ||
| Raw 264.7 macrophages | Simvastatin repressed IL-1β secretion in response to H. pylori infection and increased autophagy. | ✓ | [106] | ||
| Human monocyte derived macrophages | Statin treatment during macrophage differentiation phase led to enhanced LPS-induced IL-1β and IL-6 secretion | ✓ | [68] | ||
| RAW-Blue™ cells and Murine BMDMs | Simvastatin increased NF-κB/AP-1 activation in unstimulated and LPS-activated macrophages. LPS-induced TNF, IL-1β, and IL-6 expression was amplified. Expression of arginase-1 and GILZ was enhanced in unstimulated, LPS-, and IL-4-activated macrophages. | ✓ | ✓ | [75] | |
2.1. Statins Modulate TLR Inflammatory Signalling Pathways
Cell surface TLRs, such as TLR1, TLR2, TLR4, TLR5, and TLR6, are key initiators of innate immune responses. They are predominantly involved in host defence mechanisms through their recognition of a diverse array of stimulatory signals related to microbial membrane components, such as lipids, lipoproteins, proteins, and LPS [107]. TLR engagement triggers a range of antimicrobial responses, including the production of reactive nitrogen and oxygen species, inflammatory cytokines, and matrix metalloproteinases (MMPs). However, alongside their responsiveness to exogenous ligands, TLRs also recognise endogenous ligands (e.g., oxLDL) released from damaged tissues or dead cells, thereby regulating sterile inflammatory processes [108]. Indeed, prolonged TLR activation has been associated with uncontrolled chronic inflammatory diseases, including atherosclerosis [109][110][111]. TLR4, in particular, is upregulated in atherosclerotic plaques and demonstrates increased expression as a result of ox-LDL exposure [112][113]. TLR4 signalling is mediated by the adaptor proteins myeloid differentiation primary response 88 (MyD88) and TIR-domain-containing adapter-inducing interferon-β (TRIF), which initiate two separate signal transduction pathways that culminate in the activation of a multitude of transcription factors [114][115], including members of the NF-κB [116] and IRF [117] families. MyD88-dependent signalling cascades include the activation of NF-κB and mitogen-activated protein kinase (MAPK) family members, such as extracellular signal-regulated kinase1/2, p38, and c-Jun N-terminal kinase (JNK), which, in turn, mediate the activation of AP-1 family transcription factors or the stabilization of mRNA to regulate inflammatory responses [107]. In contrast, TRIF-mediated TLR4 signalling occurs through the activation of IFN3 and STAT1, which induce the expression of IFN genes (e.g., IFN-B) and are also involved in late-phase NF-κB activation [116][118]. A number of accessory proteins, such as CD14 and CD36, are also suggested to play a role in macrophage inflammation cascades through their association with TLR4 [38].
2.1.1. Anti-Inflammatory Modulation of TLR Signalling Pathways
As noted, NF-κB, through its activation in the TLR4 signalling pathways, is a key regulator of both macrophage inflammatory responses to pathogens and their role in sterile inflammatory diseases. Multiple statins (atorvastatin [59], fluvastatin [59][76], lovastatin [59][82], pravastatin [59], and simvastatin [76][99]) have been shown to inhibit NF-κB activation. The effects of statins on NF-κB activation are suggested to be the result of statins’ inhibition of the mevalonate pathway, specifically the isoprenoid branch, as various studies have reported that the addition of mevalonate, FPP, and GGPP reverses their action on NF-κB [76][99]. The exact links between statins’ inhibitory action on both protein prenylation and NF-κB activation have yet to be fully elucidated, although it has recently been reported that statins attenuate the degradation of the NF- κB inhibitor protein IκB [119]. IκB degradation is reliant on the phosphorylation of the IKK2 complex, which may be regulated by Rac1 in macrophages [120]. The upregulated gene and protein expression of Krüppel-like factor 2 [99] (a potent regulator of pro-inflammatory activation) and SOD1 [92] (associated with increased antioxidant enzyme activity and decreased ROS production [121]) have also been reported to occur in statin-treated macrophages and may contribute to the suppression of NF-κB-driven signalling pathways. Statin-mediated inhibition of the IκB/NF-κB pathway has been shown to result in a global anti-inflammatory effect on macrophages, with mRNA and protein analysis revealing the attenuated expression of many pro-inflammatory associated mediators, including cytokines (TNFα, IL-1β, and IL-6) [82][99], chemokines (MCP-1 and MIP-1α/β) [99], and tissue factor (a membrane-bound glycoprotein that plays a prominent role in the extrinsic pathway of blood coagulation and fibrin deposition) [76], and NO production [59][81][82]. Importantly, the inhibitory effects of statin treatment on NF-κB-induced cytokine synthesis have also been seen when using the CVD-relevant endogenous ligand oxLDL and are associated with reduced macrophage oxLDL loading and foam cell formation [62][92][94][95].
Interestingly, statin-mediated inhibition of the MyD88/NF-κB pathway has also been implicated in reducing inflammatory responses through enhancing autophagy [65][106][122][123] via the Akt-mTORC1 axis [65][122], but there are conflicting thoughts on whether this results from the inhibition of the cholesterol or isoprenoid biosynthesis branch of the mevalonate pathway [106][122][123]. The increased autophagy resulting from statin treatment has been noted to restrict NLRP3 (NOD-, LRR-, and pyrin domain-containing protein 3) inflammasome activation and thus reduce pro-inflammatory cytokine release [65][106].
In addition to signalling through NF-κB-dependent pathways, which are thought to be induced predominantly by MyD88-signalling, it has been proposed that statins’ inhibitory effects on macrophage inflammatory responses result from a downstream suppression of TRIF-mediated signalling [90]. Pravastatin and pitavastatin treatment of TLR4-stimulated RAW264 macrophages have a strong inhibitory effect on the TRIF/IRF3/IFN-β pathway in macrophages. The reduction in IFN-β expression resulting from statin treatment led to decreased STAT1 phosphorylation and the attenuation of pro-inflammatory gene expression in macrophages, evidenced by the reduced secretion of MCP-1, NO, and IL-6. Unlike previous studies, the researchers could not identify whether this action was the result of mevalonate or isoprenoid inhibition by statins, as they noted that mevalonate itself also suppressed LPS-induced expression of IFN-β [90].
Statin treatment has also been reported to reduce the matrix degrading capacity of M1-like polarized macrophages through the modulation of matrix metalloproteinase (MMP) expression [66][74][77][79]. This is particularly relevant to CVD, as atherosclerotic lesions show enhanced MMP expression, and this is thought to contribute to the weakening of the vascular wall, aiding plaque rupture [124]. Atorvastatin co-incubation during the polarization of classically activated macrophages was found to reduce MMP-14 activation [66], which is thought to mediate the expression of other MMPs, such as MMP-9. MMP-9 is one of the most widely investigated MMPs and is known to be involved in inflammation (e.g., extracellular processing of IL-1β [125]) and fibrosis in CVD [126]. In line with this, various studies have reported that statin treatment decreases MMP-9 protein secretion, thereby reducing its activity [77][79]. Importantly, this effect was also seen in in vitro studies of foamy macrophages [74], which are abundant in atherosclerotic plaques. This effect of statins is thought to be dependent on their action as mevalonate inhibitors [66][77], and there is evidence that the uncoupling of JAK/STAT signalling plays a role [79]. However, it should be noted that most of the studies examining statin-mediated effects on MMP expression in macrophages have not investigated the potential underlying mechanisms, and the exact point in the TLR-signalling pathway that is impacted awaits clarification. Macrophage production of MMPs in the absence of statin treatment is regulated via both the NF-κB [127][128] and MAPK [129] pathways.
A final means by which statins are thought to blunt TLR4-induced macrophage inflammation is not via inhibition of its signalling cascade but rather via the enhancement of anti-inflammatory response elements. In this respect, it has been reported that fluvastatin and simvastatin upregulate CD9 expression in both RAW264.7 cells and murine bone-marrow derived macrophages (BMDMs) treated with LPS [80], consequently leading to reduced TNFα and MMP-9 production. CD9 is a recognised anti-inflammatory marker of macrophages [130] and negatively regulates LPS-induced macrophage activation by preventing the formation of CD14/TLR4 complexes [131]. Indeed, statin treatment no longer resulted in significant inhibition of TNFα and MMP-9 in BMDMs from CD9 knock-out mice, suggesting that statins’ anti-inflammatory effects are, to a degree, dependent on CD9 [80]. The upregulation of CD9 observed following statin treatment appears to be dependent on their inhibitory action on protein prenylation (Figure 4), specifically geranylgeranylation, as GGTI-298 (a geranylgeranyltransferase inhibitor), but not FTI-277 (a farnesyl transferase inhibitor) increased LPS-treated CD9 levels to a comparable degree. However, the precise mechanism by which decreased isoprenoid synthesis confers CD9 upregulation is currently unknown.
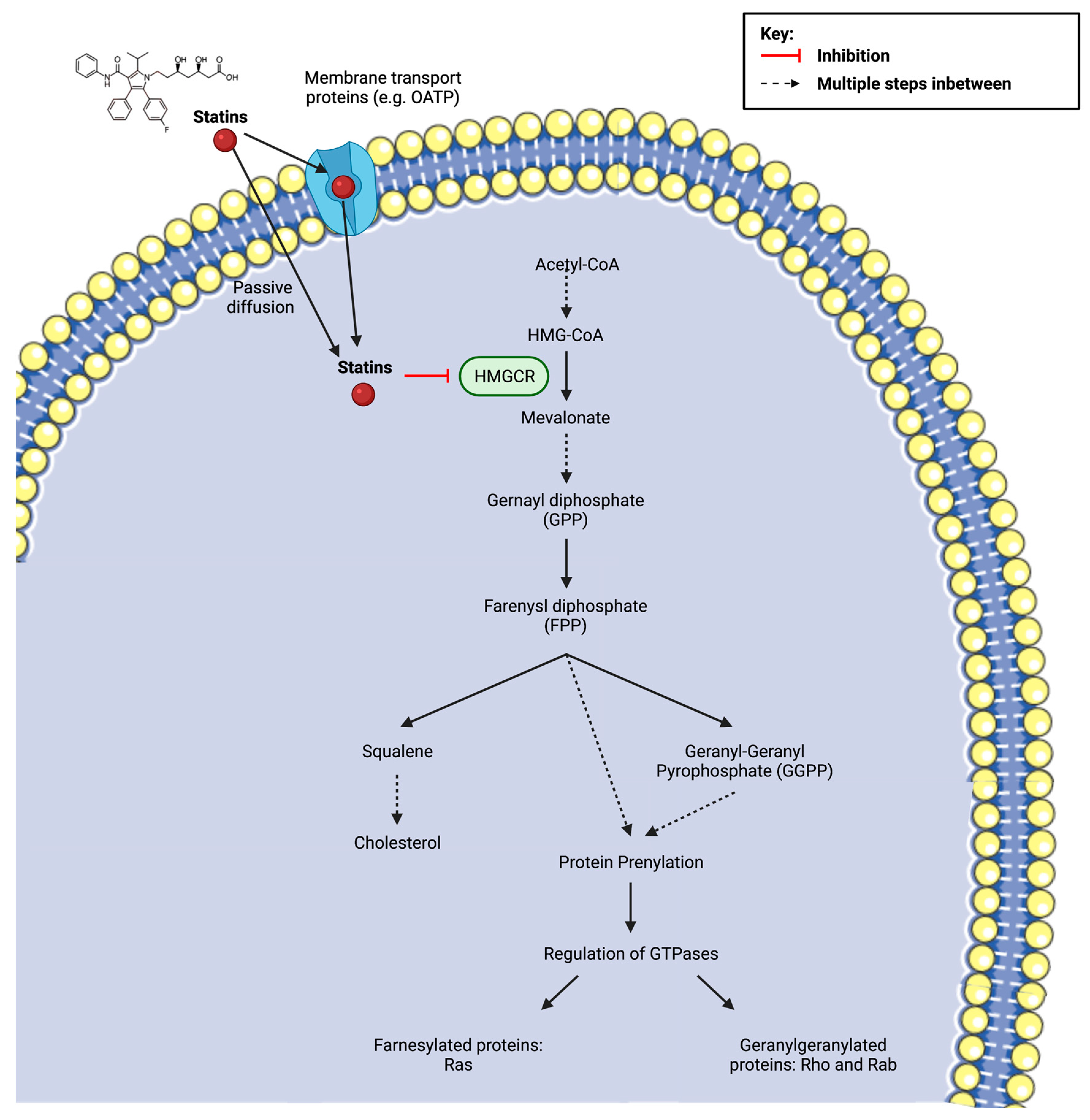
Figure 4. Statin inhibition of 3-hydroxy-3methylglutaryl coenzyme A (HMG-CoA) reductase (HMGCR) and the subsequent implications on downstream metabolites of the mevalonate pathway, including the synthesis of cholesterol and the isoprenoids farnesyl pyrophosphate (FPP) and geranylgeranyl pyrophosphate (GGPP). Protein prenylation, via isoprenoids, is essential for the activation of small guanosine triphosphate (GTP)-binding proteins (Ras, Rho, Rac). The cellular uptake of the drug depends on its solubility. Lipophilic statins are more likely to enter the cell via passive diffusion, whereas hydrophilic statins require protein transporters, such as organic anion transporting polypeptides (OATPs) in hepatocytes. Created with BioRender.com (accessed on 7 March 2022) and Smart.Servier.com (accessed on 7 March 2022).
2.1.2. Pro-Inflammatory Modulation of TLR Signalling Pathways
In contrast to the anti-inflammatory properties of statins described above, a growing number of in vitro studies are reporting that statins paradoxically enhance pro-inflammatory signalling in macrophages (Table 2). LPS-triggered TLR4 activation in macrophages activates both NF-κB and AP-1 transcription factors [132], which have both been implicated in statin-induced pro-inflammatory responses [71][96].
In one of the earliest studies [96] reporting pro-inflammatory effects, it was demonstrated that simvastatin pre-treatment enhanced LPS-induced IL-12p40 (a constituent of the bioactive cytokines IL-12 and IL-23) and TNFα mRNA expression and protein production by a mechanism involving AP-1 and C/EBP transcription factors. Specifically, statin treatment decreased c-FOS binding to the AP-1 promoter region (a negative regulator of the signalling system) whilst simultaneously enhancing JNK-mediated c-Jun phosphorylation, thereby stimulating the transcription of inflammatory genes. In keeping with this, atorvastatin and simvastatin pre-treatment is observed to enhance TLR2/TLR4 ligand-stimulated IL-6 and TNFα production [60], and various research groups have found statins to induce the activation of the MyD88 pathway transcription factor NF-κB [70][75] (alongside AP-1). There is evidence that these effects depend on the isoprenoid branch of the mevalonate pathway [96] and on Rho GTPases [70][83]. The molecular mechanisms connecting the effects of statins on GTPases and the increased expression of the AP-1 transcription factor remain poorly understood, but it has been suggested that Rho GTPase inactivation by the suppression of prenylation abolishes an inhibitory feedback loop in this pathway, thereby resulting in an enhanced upregulation of cytokine gene expression.
Statins have also been found to enhance pro-inflammatory macrophage responses by increasing NLRP3 inflammasome activation in a p38-dependent manner [71]. IL-1β is unique compared to most cytokines in that it requires post-translational modification via caspase-1 to reach its mature form, being originally translated as a 33 kDa inactive precursor (pro-IL-1β) [133]. Caspase-1, in turn, requires NLRP3 inflammasome activation to mediate this process [134]. Several studies have found that statins promote caspase-1 and NLRP3 activation and have shown that statin-stimulated IL-1β release is dependent on their enhanced activation [63][67][71]. Statin treatment is proposed to facilitate LPS-induced capase-1 and inflammasome stimulation via its disturbance of isoprenoid biosynthesis, as the effect was reversible with GGPP addition [67]. Furthermore, the deletion of geranylgeranyltransferase type 1 (GGTase-I; responsible for carrying out GTPase geranylgeranylation) in macrophages mimicked the effects of statins. Later studies by the group suggested that Rac1 mediates the hyperactivity to pro-inflammatory stimuli observed in statin-treated and GGTase-I-deficient macrophages because the deletion of Rac1 abolished the enhanced release of pro-inflammatory cytokines, whereas the deletion of other GTPases (RhoA and Cdc42) did not [67]. However, how statin-induced hyperactive Rac1 activation may drive the enhancement of LPS-stimulated p38 activation and thus increase pro-inflammatory IL-1β secretion has yet to be explored.
In consideration of the relevance of statins to atherosclerosis management, various research groups have also investigated the effects of statins on macrophage TLR-mediated cytokine responses using endogenous molecules (e.g., LDL and cholesterol crystals), with mixed findings. Lindholm and Nilsson reported that in combination with aggregated LDL (agLDL) loading, statin treatment enhanced secretion of IL-1β and IL-8 but had no effect on TNFα or IL-6 secretion in human primary monocyte-derived macrophages isolated from buffy coats [98]. Cui et al. also reported statin treatment to strongly enhance mature IL-1β release in murine BMDMs stimulated with a combination of LPS and cholesterol crystals but noted the opposite to be true in THP-1 derived macrophages [72]. Interestingly, despite the conflicting data between macrophage cell types, these effects were all reported to be isoprenoid dependent [72][98]. At present, it remains unclear which TLR-pathway signalling elements are affected by statin treatment in ox- and agLDL-stimulated macrophages but, given that (for reasons not completely understood) different TLR4 stimuli induce different cellular responses [135][136], future studies may find the involvement of signalling components outside of those noted in the LPS experiments.
It has also been suggested that statin-mediated effects on TLR-inflammatory responses may not solely be the result of their action on its signalling pathway but may also result from an increase in membrane CD14 expression [85]. RAW 264.7 macrophage incubation with lovastatin both alone and in combination with LPS promoted increased CD14 mRNA and protein levels, resulting in greater LPS-induced TNFα secretion. Coincubation of lovastatin-treated macrophages with FPP, GGPP, or water-soluble cholesterol was seen to prevent LPS-induced TNFα levels, suggesting that statin effects on macrophage responses may be regulated at multiple levels.
2.2. Statins Modulate IFN-γR Inflammatory Signalling Pathways
Cytokines are major regulators of macrophage activation, and aberrant secretion is implicated in several disease states, including chronic inflammatory diseases such as atherosclerosis. IFN-γ, particularly, is known to play a role in atherosclerotic development, being highly expressed in lesions [137] and inducing foam cell formation [138] in macrophages via increased LDL uptake. IFN-γ exerts its biological activities by binding to a specific cell surface receptor, IFN-γR, which utilises the Jak-STAT pathway in its signal transduction (a recurring theme amongst members of the cytokine receptor superfamily). Through this mechanism, IFN-γ induces the expression of numerous genes that play a role in macrophage inflammatory responses, such as ROS production and communication between macrophages and other immune cells (e.g., T lymphocytes) via chemokine secretion and surface marker expression [139]. Notably, IFN-γ is also thought to participate in an amplification loop to increase immune system sensitivity, as it has been seen to enhance LPS-induced NF-κB activation and increase TLR expression, whilst in turn, TLR ligands, such as LPS, augment local IFN-γ induction [139].
2.2.1. Anti-Inflammatory Modulation of IFN-γR Signalling Pathways
In both human and mouse-derived macrophages, a variety of statins have been found to reduce IFN-γ-induced MHC-II expression through the downregulation of the class II transactivator (CIITA), thereby interfering with their ability to prompt T cell activation, indicative of an immunosuppressive impact [57][58][86][102]. Further examination of this effect provided some insight into the potential molecular basis, with Kwak et al. and Lee et al. finding that statins specifically decrease the expression of CIITA at the transcriptional level, after noting that CIITA mRNA destabilisation did not occur in the presence of simvastatin. The transcription of IFN-γ-inducible CIITA expression is controlled by a large regulatory region containing three independent promoters pI, pIII, and pIV, which, in turn, are controlled by distinct regulatory elements [140]. As Kwak et al. [57] had noted that constitutive MHC-II expression, which is controlled by pI and pIII, was not affected by statin treatment it was suggested that pIV may be involved. Lee et al. [86] therefore focused their investigation on this particular promoter region, discovering that its transcription factors STAT1 and IRF-1 were both downregulated. In addition to this, the team also documented that the addition of GGPP, but not cholesterol, abolished the statin-mediated reduction in IFN-γ-induced MHC-II expression, signifying again that the effect was likely to be dependent on statins’ action as isoprenoid inhibitors. They next tested the effects of two specific inhibitors of Ras superfamily protein prenylation: GGTI-298 and FTI-277. GGTI-298 was found to mimic the inhibitory actions of simvastatin on CIITA expression, but FTI-277 had no effect, indicating the specific involvement of geranylgeranylation. Furthermore, a Rac1-specific inhibitor was also shown to capture this effect, revealing its contribution to IFN-γ-induced STAT1 activation. Another potential factor leading to STAT1 suppression was suggested by Huang et al., who demonstrated that lovastatin and fluvastatin upregulate mRNA expression of the Socs-3 gene in macrophages [78]. SOCS proteins are known to negatively regulate cytokine signalling through their binding to the cytoplasmic domain of recognition receptors [141]. Regardless of the precise signalling mechanisms involved, the dampening of IFN-γ inflammatory stimulation via STAT1 inhibition has also been found to affect a number of other pro-inflammatory responses, including reduced mRNA expression of chemokines (monocyte chemotactic protein-1 (MCP-1) and macrophage inflammatory proteins-1 α and β (MIP-1α/β)) [90][97][103], chemokine receptors (CCRs—CCR1, CCR2, and CCR5) [97] and cytokines (IL-6), along with reduced NO production [59].
2.2.2. Pro-Inflammatory Modulation of IFN-γR Signalling Pathways
Interestingly, reports of statins enhancing pro-inflammatory signalling have not cited the involvement of IFN-γR pathways. Indeed, although simvastatin pre-treatment was found to enhance IL-12p40 and TNFα production in murine macrophages stimulated with both IFN-γ and L. monocytogenes infection [102], the researchers highlighted that this was most likely to be the result of TLR-mediated signalling pathways as they found that IFN-γ treatment alone in macrophages had no effect on pro-inflammatory cytokine production. Moreover, in agreement with anti-inflammatory reports, they noted a decreased surface expression of MHC-II. Another study by Linnenberger et al. agreed with this finding that statin treatment had no effect on macrophage stimulation by IFN-γ (despite enhancing LPS-induced expression of TNF, IL-1β, and IL-6) [75].
2.3. Statins Play Roles in Macrophage Differentiation
Alongside their effects on inflammatory signalling pathways, more recent studies have suggested that statins may directly alter the differentiation of macrophages in vitro. In one study, atorvastatin enhanced an IL-4-induced M2 phenotype via p38 MAPK-dependent PPARγ activation when added at the start of the differentiation process [64]. However, in other work, macrophages differentiated overnight in the presence of fluvastatin were more reactive to LPS stimulation than those that were not, characterised by a greater secretion of IL-1β and IL-6 and dependent on Rac1-geranylgeranylation [68]. Taken together, these studies suggest that macrophages differentiated in the presence of statins may be more immune-responsive to various stimuli and therefore can enhance either pro or anti-inflammatory functions depending on the particular stimulating agents they are exposed to.
References
- Cardiovascular Diseases (CVDs). Available online: https://www.who.int/news-room/fact-sheets/detail/cardiovascular-diseases-(cvds) (accessed on 21 February 2022).
- Frostegård, J. Immunity, Atherosclerosis and Cardiovascular Disease. BMC Med. 2013, 11, 117.
- Parisi, L.; Gini, E.; Baci, D.; Tremolati, M.; Fanuli, M.; Bassani, B.; Farronato, G.; Bruno, A.; Mortara, L. Macrophage Polarization in Chronic Inflammatory Diseases: Killers or Builders? J. Immunol. Res. 2018, 2018, 8917804.
- Bhatnagar, P.; Wickramasinghe, K.; Wilkins, E.; Townsend, N. Trends in the Epidemiology of Cardiovascular Disease in the UK. Heart 2016, 102, 1945–1952.
- Mensah, G.A.; Wei, G.S.; Sorlie, P.D.; Fine, L.J.; Rosenberg, Y.; Kaufmann, P.G.; Mussolino, M.E.; Hsu, L.L.; Addou, E.; Engelgau, M.M.; et al. Decline in Cardiovascular Mortality: Possible Causes and Implications. Circ. Res. 2017, 120, 366–380.
- Pirillo, A.; Casula, M.; Olmastroni, E.; Norata, G.D.; Catapano, A.L. Global Epidemiology of Dyslipidaemias. Nat. Rev. Cardiol. 2021, 18, 689–700.
- How Statin Drugs Protect the Heart—Johns Hopkins Medicine. Available online: https://www.hopkinsmedicine.org/health/wellness-and-prevention/how-statin-drugs-protect-the-heart (accessed on 21 February 2022).
- Silverman, M.G.; Ference, B.A.; Im, K.; Wiviott, S.D.; Giugliano, R.P.; Grundy, S.M.; Braunwald, E.; Sabatine, M.S. Association between Lowering LDL-C and Cardiovascular Risk Reduction among Different Therapeutic Interventions: A Systematic Review and Meta-Analysis. JAMA 2016, 316, 1289–1297.
- Collins, R.; Reith, C.; Emberson, J.; Armitage, J.; Baigent, C.; Blackwell, L.; Blumenthal, R.; Danesh, J.; Smith, G.D.; DeMets, D.; et al. Interpretation of the Evidence for the Efficacy and Safety of Statin Therapy. Lancet 2016, 388, 2532–2561.
- Lusis, A.J.; Mar, R.; Pajukanta, P. Genetics of Atherosclerosis. Annu. Rev. Genom. Hum. Genet. 2004, 5, 189–218.
- Pfizer Ireland Pharmaceuticals. Lipitor® (Atorvastatin Calcium) Highlights of Prescribing Information. 2009. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2009/020702s056lbl.pdf (accessed on 23 March 2022).
- Law, M.R.; Wald, N.J.; Rudnicka, A.R. Quantifying Effect of Statins on Low Density Lipoprotein Cholesterol, Ischaemic Heart Disease, and Stroke: Systematic Review and Meta-Analysis. BMJ 2003, 326, 1423.
- Duerden, M.; O’Flynn, N.; Qureshi, N. Cardiovascular Disease: Risk Assessment and Reduction, Including Lipid Modification Clinical Guideline. Br. J. Gen. Pract. 2015, 65, 378–380.
- Schachter, M. Chemical, Pharmacokinetic and Pharmacodynamic Properties of Statins: An Update. Fundam. Clin. Pharmacol. 2005, 19, 117–125.
- Bayer Corporation. BAYCOL ® (Cerivastatin Sodium) Highlights of Prescribing Information. 2000. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/nda/2000/20-740S008_Baycol_prntlbl.pdf (accessed on 23 March 2022).
- Adams, S.P.; Tiellet, N.; Alaeiilkhchi, N.; Wright, J.M.; Group, C.H. Cerivastatin for Lowering Lipids. Cochrane Database Syst. Rev. 2020, 1, CD012501.
- Novartis Pharmaceuticals Corporation. Lescol® XL (Fluvastatin Sodium) Highlights of Prescribing Information. 2012. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2012/021192s019lbl.pdf (accessed on 23 March 2022).
- Merck Sharp & Dohme Corp. MEVACOR® (Lovastatin) Highlights of Prescribing Information. 2012. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2012/019643s085lbl.pdf (accessed on 23 March 2022).
- Roy, M.; Kung, H.-J.; Ghosh, P.M. Statins and Prostate Cancer: Role of Cholesterol Inhibition vs. Prevention of Small GTP-Binding Proteins. Am. J. Cancer Res. 2011, 1, 542.
- Kowa Pharmaceuticals America Inc. LIVALO® (Pitavastatin) Highlights of Prescribing Information. 2019. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2019/022363s015lbl.pdf (accessed on 23 March 2022).
- Adams, S.P.; Alaeiilkhchi, N.; Wright, J.M. Pitavastatin for Lowering Lipids. Cochrane Database Syst. Rev. 2020, 6, CD012735.
- Bristol-Myers Squibb Company. PRAVACHOL® (Pravastatin Sodium) Highlights of Prescribing Information. 2011. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/nda/2011/019898Orig1s061.pdf (accessed on 23 March 2022).
- AstraZeneca. CRESTOR (Rosuvastatin Calcium) Highlights of Prescribing Information. 2010. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2010/021366s016lbl.pdf (accessed on 23 March 2022).
- Marck & Co., Inc. ZOCOR (Simvastatin) Highlights of Prescribing Information. 2010. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2010/019766s078lbl.pdf (accessed on 23 March 2022).
- Furberg, C.D.; Pitt, B. Withdrawal of Cerivastatin from the World Market. Curr. Control. Trials Cardiovasc. Med. 2001, 2, 205.
- Endo, A. The Origin of the Statins. Atheroscler. Suppl. 2004, 5, 125–130.
- Lippi, G.; Mattiuzzi, C.; Cervellin, G. Statins Popularity: A Global Picture. Br. J. Clin. Pharmacol. 2019, 85, 1614–1615.
- Wang, Y.; Nichol, M.B.; Yan, B.P.Y.; Wu, J.; Tomlinson, B.; Lee, V.W.Y. Descriptive Analysis of Real-World Medication Use Pattern of Statins and Antiplatelet Agents among Patients with Acute Coronary Syndrome in Hong Kong and the USA. BMJ Open 2019, 9, e024937.
- Zhou, Q.; Liao, J.K. Pleiotropic Effects of Statins—Basic Research and Clinical Perspectives. Circ. J. 2010, 74, 818–826.
- Climent, E.; Benaiges, D.; Pedro-Botet, J. Hydrophilic or Lipophilic Statins? Front. Cardiovasc. Med. 2021, 8, 687585.
- Moroni, F.; Ammirati, E.; Norata, G.D.; Magnoni, M.; Camici, P.G. The Role of Monocytes and Macrophages in Human Atherosclerosis, Plaque Neoangiogenesis, and Atherothrombosis. Mediators Inflamm. 2019, 2019, 7434376.
- Moore, K.J.; Sheedy, F.J.; Fisher, E.A. Macrophages in Atherosclerosis: A Dynamic Balance. Nat. Rev. Immunol. 2013, 13, 709.
- Beck-Joseph, J.; Lehoux, S. Molecular Interactions Between Vascular Smooth Muscle Cells and Macrophages in Atherosclerosis. Front. Cardiovasc. Med. 2021, 8, 737934.
- Anlamlert, W.; Lenbury, Y.; Bell, J. Modeling Fibrous Cap Formation in Atherosclerotic Plaque Development: Stability and Oscillatory Behavior. Adv. Differ. Equ. 2017, 2017, 195.
- Mosser, D.M.; Hamidzadeh, K.; Goncalves, R. Macrophages and the Maintenance of Homeostasis. Cell. Mol. Immunol. 2020, 18, 579–587.
- Shapouri-Moghaddam, A.; Mohammadian, S.; Vazini, H.; Taghadosi, M.; Esmaeili, S.A.; Mardani, F.; Seifi, B.; Mohammadi, A.; Afshari, J.T.; Sahebkar, A. Macrophage Plasticity, Polarization, and Function in Health and Disease. J. Cell. Physiol. 2018, 233, 6425–6440.
- Murray, P.J.; Allen, J.E.; Biswas, S.K.; Fisher, E.A.; Gilroy, D.W.; Goerdt, S.; Gordon, S.; Hamilton, J.A.; Ivashkiv, L.B.; Lawrence, T.; et al. Macrophage Activation and Polarization: Nomenclature and Experimental Guidelines. Immunity 2014, 41, 14–20.
- Viola, A.; Munari, F.; Sánchez-Rodríguez, R.; Scolaro, T.; Castegna, A. The Metabolic Signature of Macrophage Responses. Front. Immunol. 2019, 10, 1462.
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-ΚB Signaling in Inflammation. Sig. Transduct Target Ther. 2017, 2, 17023.
- Bode, J.G.; Ehlting, C.; Häussinger, D. The Macrophage Response towards LPS and Its Control through the P38MAPK–STAT3 Axis. Cell. Signal. 2012, 24, 1185–1194.
- Yamamoto, M.; Sato, S.; Hemmi, H.; Hoshino, K.; Kaisho, T.; Sanjo, H.; Takeuchi, O.; Sugiyama, M.; Okabe, M.; Takeda, K.; et al. Role of Adaptor TRIF in the MyD88-Independent Toll-like Receptor Signaling Pathway. Science 2003, 301, 640–643.
- Von Knethen, A.; Callsen, D.; Brüne, B. NF-KappaB and AP-1 Activation by Nitric Oxide Attenuated Apoptotic Cell Death in RAW 264.7 Macrophages. Mol. Biol. Cell 1999, 10, 361–372.
- Bonecchi, R.; Garlanda, C.; Mantovani, A.; Riva, F. Cytokine Decoy and Scavenger Receptors as Key Regulators of Immunity and Inflammation. Cytokine 2016, 87, 37.
- Wynn, T.A.; Vannella, K.M. Review Macrophages in Tissue Repair, Regeneration, and Fibrosis. Immunity 2016, 44, 450–462.
- Mosser, D.M.; Edwards, J.P. Exploring the Full Spectrum of Macrophage Activation. Nat. Rev. Immunol. 2008, 8, 958–969.
- Chávez-Galán, L.; Olleros, M.L.; Vesin, D.; Garcia, I. Much More than M1 and M2 Macrophages, There Are Also CD169+ and TCR+ Macrophages. Front. Immunol. 2015, 6, 263.
- Nahrendorf, M.; Swirski, F.K. Abandoning M1/M2 for a Network Model of Macrophage Function. Circ. Res. 2016, 119, 414.
- Stöger, J.L.; Gijbels, M.J.J.; van der Velden, S.; Manca, M.; van der Loos, C.M.; Biessen, E.A.L.; Daemen, M.J.A.P.; Lutgens, E.; de Winther, M.P.J. Distribution of Macrophage Polarization Markers in Human Atherosclerosis. Atherosclerosis 2012, 225, 461–468.
- Chinetti-Gbaguidi, G.; Baron, M.; Bouhlel, M.A.; Vanhoutte, J.; Copin, C.; Sebti, Y.; Derudas, B.; Mayi, T.; Bories, G.; Tailleux, A.; et al. Human Atherosclerotic Plaque Alternative Macrophages Display Low Cholesterol Handling but High Phagocytosis Because of Distinct Activities of the PPARγ and LXRα Pathways. Circ. Res. 2011, 108, 985–995.
- Feig, J.E.; Parathath, S.; Rong, J.X.; Mick, S.L.; Vengrenyuk, Y.; Grauer, L.; Young, S.G.; Fisher, E.A. Reversal of Hyperlipidemia with a Genetic Switch Favorably Affects the Content and Inflammatory State of Macrophages in Atherosclerotic Plaques. Circulation 2011, 123, 989–998.
- Hewing, B.; Parathath, S.; Mai, C.K.; Fiel, M.I.; Guo, L.; Fisher, E.A. Rapid Regression of Atherosclerosis with MTP Inhibitor Treatment. Atherosclerosis 2013, 227, 125–129.
- Rayner, K.J.; Sheedy, F.J.; Esau, C.C.; Hussain, F.N.; Temel, R.E.; Parathath, S.; Van Gils, J.M.; Rayner, A.J.; Chang, A.N.; Suarez, Y.; et al. Antagonism of MiR-33 in Mice Promotes Reverse Cholesterol Transport and Regression of Atherosclerosis. J. Clin. Investig. 2011, 121, 2921–2931.
- Varnava, A.M.; Mills, P.G.; Davies, M.J. Relationship Between Coronary Artery Remodeling and Plaque Vulnerability. Circulation 2002, 105, 939–943.
- Poh, A.R.; Ernst, M. Targeting Macrophages in Cancer: From Bench to Bedside. Front. Oncol. 2018, 8, 49.
- Ma, W.T.; Gao, F.; Gu, K.; Chen, D.K. The Role of Monocytes and Macrophages in Autoimmune Diseases: A Comprehensive Review. Front. Immunol. 2019, 10, 1140.
- Cheng, Y.; Marion, T.N.; Cao, X.; Wang, W.; Cao, Y. Park 7: A Novel Therapeutic Target for Macrophages in Sepsis-Induced Immunosuppression. Front. Immunol. 2018, 9, 2632.
- Kwak, B.; Mulhaupt, F.; Myit, S.; Mach, F. Statins as a Newly Recognized Type of Immunomodulator. Nat. Med. 2000, 6, 1399–1402.
- Youssef, S.; Stüve, O.; Patarroyo, J.O.; Ruiz, P.J.; Radosevich, J.L.; Mi Hur, E.; Bravo, M.; Mitchell, D.J.; Sobel, R.A.; Stelnman, L.; et al. The HMG-CoA Reductase Inhibitor, Atorvastatin, Promotes a Th2 Bias and Reverses Paralysis in Central Nervous System Autoimmune Disease. Nature 2002, 420, 78–84.
- Huang, K.-C.; Chen, C.-W.; Chen, J.-C.; Lin, W.-W. HMG-CoA Reductase Inhibitors Inhibit Inducible Nitric Oxide Synthase Gene Expression in Macrophages. J. Biomed. Sci. 2003, 10, 396–405.
- Helmers, A.J.; Gowda, D.C.; Kain, K.C.; Liles, W.C. Statins Fail to Improve Outcome in Experimental Cerebral Malaria and Potentiate Toll-Like Receptor-Mediated Cytokine Production by Murine Macrophages. Am. J. Trop. Med. Hyg. 2009, 81, 631–637.
- Lee, D.K.; Park, E.J.; Kim, E.K.; Jin, J.; Kim, J.S.; Shin, I.J.; Kim, B.Y.; Lee, H.; Kim, D.E. Atorvastatin and Simvastatin, but Not Pravastatin, Up-Regulate LPS-Induced MMP-9 Expression in Macrophages by Regulating Phosphorylation of ERK and CREB. Cell. Physiol. Biochem. 2012, 30, 499–511.
- Shao, Q.; Shen, L.H.; Hu, L.A.; Pu, J.; Jing, Q.; He, B. Atorvastatin Suppresses Inflammatory Response Induced by OxLDL through Inhibition of ERK Phosphorylation, IκBα Degradation, and COX-2 Expression in Murine Macrophages. J. Cell. Biochem. 2012, 113, 611–618.
- Henriksbo, B.D.; Lau, T.C.; Cavallari, J.F.; Denou, E.; Chi, W.; Lally, J.S.; Crane, J.D.; Duggan, B.M.; Foley, K.P.; Fullerton, M.D.; et al. Fluvastatin Causes NLRP3 Inflammasome-Mediated Adipose Insulin Resistance. Diabetes 2014, 63, 3742–3747.
- Zhang, O.; Zhang, J. Atorvastatin Promotes Human Monocyte Differentiation toward Alternative M2 Macrophages through P38 Mitogen-Activated Protein Kinase-Dependent Peroxisome Proliferator-Activated Receptor γ Activation. Int. Immunopharmacol. 2015, 26, 58–64.
- Han, F.; Xiao, Q.Q.; Peng, S.; Che, X.Y.; Jiang, L.S.; Shao, Q.; He, B. Atorvastatin Ameliorates LPS-Induced Inflammatory Response by Autophagy via AKT/MTOR Signaling Pathway. J. Cell. Biochem. 2018, 119, 1604–1615.
- Hohensinner, P.J.; Baumgartner, J.; Ebenbauer, B.; Thaler, B.; Fischer, M.B.; Huber, K.; Speidl, W.S.; Wojta, J. Statin Treatment Reduces Matrix Degradation Capacity of Proinflammatory Polarized Macrophages. Vascul. Pharmacol. 2018, 110, 49–54.
- Akula, M.K.; Ibrahim, M.X.; Ivarsson, E.G.; Khan, O.M.; Kumar, I.T.; Erlandsson, M.; Karlsson, C.; Xu, X.; Brisslert, M.; Wang, D.; et al. Protein Prenylation Restrains Innate Immunity by Inhibiting Rac1 Effector Interactions. Nat. Commun. 2019, 10, 3975.
- Fu, H.; Alabdullah, M.; Großmann, J.; Spieler, F.; Abdosh, R.; Lutz, V.; Kalies, K.; Knöpp, K.; Rieckmann, M.; Koch, S.; et al. The Differential Statin Effect on Cytokine Production of Monocytes or Macrophages Is Mediated by Differential Geranylgeranylation-Dependent Rac1 Activation. Cell Death Dis. 2019, 10, 880.
- Gasbarrino, K.; Hafiane, A.; Zheng, H.; Daskalopoulou, S.S. Intensive Statin Therapy Compromises the Adiponectin-AdipoR Pathway in the Human Monocyte-Macrophage Lineage. Stroke 2019, 50, 3609–3617.
- Healy, A.; Berus, J.M.; Christensen, J.L.; Lee, C.; Mantsounga, C.; Dong, W.; Watts, J.P.; Assali, M.; Ceneri, N.; Nilson, R.; et al. Statins Disrupt Macrophage Rac1 Regulation Leading to Increased Atherosclerotic Plaque Calcification. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 714–732.
- Henriksbo, B.D.; Tamrakar, A.K.; Phulka, J.S.; Barra, N.G.; Schertzer, J.D. Statins Activate the NLRP3 Inflammasome and Impair Insulin Signaling via P38 and MTOR. Am. J. Physiol.-Endocrinol. Metab. 2020, 319, E110–E116.
- Cui, H.; Soga, K.; Tamehiro, N.; Adachi, R.; Hachisuka, A.; Hirose, A.; Kondo, K.; Nishimaki-Mogami, T. Statins Repress Needle-like Carbon Nanotube- or Cholesterol Crystal-Stimulated IL-1β Production by Inhibiting the Uptake of Crystals by Macrophages. Biochem. Pharmacol. 2021, 188, 114580.
- Aikawa, M.; Rabkin, E.; Sugiyama, S.; Voglic, S.J.; Fukumoto, Y.; Furukawa, Y.; Shiomi, M.; Schoen, F.J.; Libby, P. An HMG-CoA Reductase Inhibitor, Cerivastatin, Suppresses Growth of Macrophages Expressing Matrix Metalloproteinases and Tissue Factor in Vivo and in Vitro. Circulation 2001, 103, 276–283.
- Luan, Z.; Chase, A.J.; Newby, A.C. Statins Inhibit Secretion of Metalloproteinases-1, -2, -3, and -9 from Vascular Smooth Muscle Cells and Macrophages. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 769–775.
- Linnenberger, R.; Hoppstädter, J.; Wrublewsky, S.; Ampofo, E.; Kiemer, A.K. Statins and Bempedoic Acid: Different Actions of Cholesterol Inhibitors on Macrophage Activation. Int. J. Mol. Sci. 2021, 22, 12480.
- Colli, S.; Eligini, S.; Lalli, M.; Camera, M.; Paoletti, R.; Tremoli, E. Vastatins Inhibit Tissue Factor in Cultured Human Macrophages. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 265–272.
- Bellosta, S.; Via, D.; Canavesi, M.; Pfister, P.; Fumagalli, R.; Paoletti, R.; Bernini, F. HMG-CoA Reductase Inhibitors Reduce MMP-9 Secretion by Macrophages. Arterioscler. Thromb. Vasc. Biol. 1998, 18, 1671–1678.
- Huang, K.C.; Chen, C.W.; Chen, J.C.; Lin, W.W. Statins Induce Suppressor of Cytokine Signaling-3 in Macrophages. FEBS Lett. 2003, 555, 385–389.
- Rival, Y.; Benéteau, N.; Chapuis, V.; Taillandier, T.; Lestienne, F.; Dupont-Passelaigue, E.; Patoiseau, J.F.; Colpaert, F.C.; Junquéro, D. Cardiovascular Drugs Inhibit MMP-9 Activity from Human THP-1 Macrophages. DNA Cell Biol. 2004, 23, 283–292.
- Jin, Y.; Tachibana, I.; Takeda, Y.; He, P.; Kang, S.; Suzuki, M.; Kuhara, H.; Tetsumoto, S.; Tsujino, K.; Minami, T.; et al. Statins Decrease Lung Inflammation in Mice by Upregulating Tetraspanin CD9 in Macrophages. PLoS ONE 2013, 8, e73706.
- Kauerova, S.; Bartuskova, H.; Muffova, B.; Janousek, L.; Fronek, J.; Petras, M.; Poledne, R.; Lesna, I.K. Statins Directly Influence the Polarization of Adipose Tissue Macrophages: A Role in Chronic Inflammation. Biomedicines 2021, 9, 211.
- Pahan, K.; Sheikh, F.G.; Namboodiri, M.S.; Singh, I. Lovastatin and Phenylacetate Inhibit the Induction of Nitric Oxide Synthase and Cytokines in Rat Primary Astrocytes, Microglia, and Macrophages. J. Clin. Investig. 1997, 100, 2671–2679.
- Monick, M.M.; Powers, L.S.; Butler, N.S.; Hunninghake, G.W. Inhibition of Rho Family GTPases Results in Increased TNF-α Production After Lipopolysaccharide Exposure. J. Immunol. 2003, 171, 2625–2630.
- Arikan, M.C.; Shapiro, S.D.; Mariani, T.J. Induction of Macrophage Elastase (MMP-12) Gene Expression by Statins. J. Cell. Physiol. 2005, 204, 139–145.
- Frey, T.; De Maio, A. Increased Expression of CD14 in Macrophages after Inhibition of the Cholesterol Biosynthetic Pathway by Lovastatin. Mol. Med. 2007, 13, 592–604.
- Lee, S.J.; Qin, H.; Benveniste, E.N. The IFN-γ-Induced Transcriptional Program of the CIITA Gene Is Inhibited by Statins. Eur. J. Immunol. 2008, 38, 2325.
- Zanin, V.; Marcuzzi, A.; Kleiner, G.; Piscianz, E.; Monasta, L.; Zacchigna, S.; Crovella, S.; Zauli, G. Lovastatin Dose-Dependently Potentiates the Pro-Inflammatory Activity of Lipopolysaccharide Both In Vitro and In Vivo. J. Cardiovasc. Transl. Res. 2013, 6, 981.
- Chow, O.A.; Von Köckritz-Blickwede, M.; Bright, A.T.; Hensler, M.E.; Zinkernagel, A.S.; Cogen, A.L.; Gallo, R.L.; Monestier, M.; Wang, Y.; Glass, C.K.; et al. Statins Enhance Formation of Phagocyte Extracellular Traps. Cell Host Microbe 2010, 8, 445–454.
- Yang, Z.; Huang, Y.C.T.; Koziel, H.; de Crom, R.; Ruetten, H.; Wohlfart, P.; Thomsen, R.W.; Kahlert, J.A.; Sørensen, H.T.; Jozefowski, S.; et al. Female Resistance to Pneumonia Identifies Lung Macrophage Nitric Oxide Synthase-3 as a Therapeutic Target. ELife 2014, 3, e03711.
- Abe, M.; Matsuda, M.; Kobayashi, H.; Miyata, Y.; Nakayama, Y.; Komuro, R.; Fukuhara, A.; Shimomura, I. Effects of Statins on Adipose Tissue Inflammation: Their Inhibitory Effect on MyD88-Independent IRF3/IFN-β Pathway in Macrophages. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 871–877.
- Furman, C.; Copin, C.; Kandoussi, M.; Davidson, R.; Moreau, M.; McTaggiart, F.; Chapman, M.J.; Fruchart, J.C.; Rouis, M. Rosuvastatin Reduces MMP-7 Secretion by Human Monocyte-Derived Macrophages: Potential Relevance to Atherosclerotic Plaque Stability. Atherosclerosis 2004, 174, 93–98.
- Verreth, W.; De Keyzer, D.; Davey, P.C.; Geeraert, B.; Mertens, A.; Herregods, M.C.; Smith, G.; Desjardins, F.; Balligand, J.L.; Holvoet, P. Rosuvastatin Restores Superoxide Dismutase Expression and Inhibits Accumulation of Oxidized LDL in the Aortic Arch of Obese Dyslipidemic Mice. Br. J. Pharmacol. 2007, 151, 347–355.
- Ishibashi, Y.; Matsui, T.; Takeuchi, M.; Yamagishi, S. Rosuvastatin Blocks Advanced Glycation End Products-Elicited Reduction of Macrophage Cholesterol Efflux by Suppressing NADPH Oxidase Activity via Inhibition of Geranylgeranylation of Rac-1. Horm. Metab. Res. 2011, 43, 619–624.
- Suh, J.S.; Lee, S.H.; Fouladian, Z.; Lee, J.Y.; Kim, T.; Kang, M.K.; Lusis, A.J.; Boström, K.I.; Kim, R.H.; Park, N.H. Rosuvastatin Prevents the Exacerbation of Atherosclerosis in Ligature-Induced Periodontal Disease Mouse Model. Sci. Rep. 2020, 10, 6383.
- Giroux, L.M.; Davignon, J.; Naruszewicz, M. Simvastatin Inhibits the Oxidation of Low-Density Lipoproteins by Activated Human Monocyte-Derived Macrophages. Biochim. Biophys. Acta-Lipids Lipid Metab. 1993, 1165, 335–338.
- Matsumoto, M.; Einhaus, D.; Gold, E.S.; Aderem, A. Simvastatin Augments Lipopolysaccharide-Induced Proinflammatory Responses in Macrophages by Differential Regulation of the c-Fos and c-Jun Transcription Factors. J. Immunol. 2004, 172, 7377–7384.
- Veillard, N.R.; Braunersreuther, V.; Arnaud, C.; Burger, F.; Pelli, G.; Steffens, S.; Mach, F. Simvastatin Modulates Chemokine and Chemokine Receptor Expression by Geranylgeranyl Isoprenoid Pathway in Human Endothelial Cells and Macrophages. Atherosclerosis 2006, 188, 51–58.
- Lindholm, M.W.; Nilsson, J. Simvastatin Stimulates Macrophage Interleukin-1β Secretion through an Isoprenylation-Dependent Mechanism. Vascul. Pharmacol. 2007, 46, 91–96.
- Tuomisto, T.T.; Lumivuori, H.; Kansanen, E.; Häkkinen, S.K.; Turunen, M.P.; Van Thienen, J.V.; Horrevoets, A.J.; Levonen, A.L.; Ylä-Herttuala, S. Simvastatin Has an Anti-Inflammatory Effect on Macrophages via Upregulation of an Atheroprotective Transcription Factor, Kruppel-like Factor 2. Cardiovasc. Res. 2008, 78, 175–184.
- Benati, D.; Ferro, M.; Savino, M.T.; Ulivieri, C.; Schiavo, E.; Nuccitelli, A.; Pasini, F.L.; Baldari, C.T. Opposite Effects of Simvastatin on the Bactericidal and Inflammatory Response of Macrophages to Opsonized S. Aureus. J. Leukoc. Biol. 2010, 87, 433–442.
- Burns, E.M.; Smelser, L.K.; Then, J.E.; Stankiewicz, T.E.; Kushdilian, M.; McDowell, S.A.; Bruns, H.A. Short Term Statin Treatment Improves Survival and Differentially Regulates Macrophage-Mediated Responses to Staphylococcus Aureus. Curr. Pharm. Biotechnol. 2013, 14, 233–241.
- Parihar, S.P.; Guler, R.; Lang, D.M.; Suzuki, H.; Marais, A.D.; Brombacher, F. Simvastatin Enhances Protection against Listeria Monocytogenes Infection in Mice by Counteracting Listeria-Induced Phagosomal Escape. PLoS ONE 2013, 8, e75490.
- Alkorashy, M. Anti-Inflammatory Actions of Statins in Human Macrophages. Maarab Alkorashy and Dipak P. Ramji. Atherosclerosis 2014, 235, e266.
- Davaro, F.; Forde, S.D.; Garfield, M.; Jiang, Z.; Halmen, K.; Tamburro, N.D.; Kurt-Jones, E.; Fitzgerald, K.A.; Golenbock, D.T.; Wang, D. 3-Hydroxyl-3-Methylglutaryl Coenzyme A (HMG-CoA) Reductase Inhibitor (Statin)-Induced 28-KDa Interleukin-1β Interferes with Mature IL-1β Signaling. J. Biol. Chem. 2014, 289, 16214–16222.
- Parihar, S.P.; Hartley, M.A.; Hurdayal, R.; Guler, R.; Brombacher, F. Topical Simvastatin as Host-Directed Therapy against Severity of Cutaneous Leishmaniasis in Mice. Sci. Rep. 2016, 6, 33458.
- Liao, W.C.; Huang, M.Z.; Wang, M.L.; Lin, C.J.; Lu, T.L.; Lo, H.R.; Pan, Y.J.; Sun, Y.C.; Kao, M.C.; Lim, H.J.; et al. Statin Decreases Helicobacter Pylori Burden in Macrophages by Promoting Autophagy. Front. Cell. Infect. Microbiol. 2017, 6, 203.
- Kawasaki, T.; Kawai, T. Toll-like Receptor Signaling Pathways. Front. Immunol. 2014, 5, 461.
- Yu, L.; Wang, L.; Chen, S. Endogenous Toll-like Receptor Ligands and Their Biological Significance. J. Cell. Mol. Med. 2010, 14, 2592.
- Andreakos, E.; Foxwell, B.; Feldmann, M. Is Targeting Toll-like Receptors and Their Signaling Pathway a Useful Therapeutic Approach to Modulating Cytokine-Driven Inflammation? Immunol. Rev. 2004, 202, 250–265.
- Mullick, A.E.; Tobias, P.S.; Curtiss, L.K. Modulation of Atherosclerosis in Mice by Toll-like Receptor 2. J. Clin. Investig. 2005, 115, 3149–3156.
- Mullick, A.E.; Tobias, P.S.; Curtiss, L.K. Toll-like Receptors and Atherosclerosis. Immunol. Res. 2006, 34, 193–209.
- Li, H.; Sun, B. Toll-like Receptor 4 in Atherosclerosis. J. Cell. Mol. Med. 2007, 11, 88.
- Michelsen, K.S.; Doherty, T.M.; Shah, P.K.; Arditi, M. Role of Toll-like Receptors in Atherosclerosis. Circ. Res. 2004, 95, e96–e97.
- Iwasaki, A.; Medzhitov, R. Toll-like Receptor Control of the Adaptive Immune Responses. Nat. Immunol. 2004, 5, 987–995.
- Takeda, K.; Akira, S. Toll-like Receptors in Innate Immunity. Int. Immunol. 2005, 17, 1–14.
- Kawai, T.; Akira, S. Signaling to NF-KB by Toll-like Receptors. Trends Mol. Med. 2007, 13, 460–469.
- Hertzog, P.J.; O’Neill, L.A.; Hamilton, J.A. The Interferon in TLR Signaling: More than Just Antiviral. Trends Immunol. 2003, 24, 534–539.
- Kagan, J.C.; Su, T.; Horng, T.; Chow, A.; Akira, S.; Medzhitov, R. TRAM Couples Endocytosis of Toll-like Receptor 4 to the Induction of Interferon-β. Nat. Immunol. 2008, 9, 361.
- Hilgendorff, A.; Muth, H.; Parviz, B.; Staubitz, A.; Haberbosch, W.; Tillmanns, H.; Hölschermann, H. Statins Differ in Their Ability to Block NF-KappaB Activation in Human Blood Monocytes. Int. J. Clin. Pharmacol. Ther. 2003, 41, 397–401.
- Chen, B.C.; Kang, J.C.; Lu, Y.T.; Hsu, M.J.; Liao, C.C.; Chiu, W.T.; Yeh, F.L.; Lin, C.H. Rac1 Regulates Peptidoglycan-Induced Nuclear Factor-ΚB Activation and Cyclooxygenase-2 Expression in RAW 264.7 Macrophages by Activating the Phosphatidylinositol 3-Kinase/Akt Pathway. Mol. Immunol. 2009, 46, 1179–1188.
- Hwang, J.; Jin, J.; Jeon, S.; Moon, S.H.; Park, M.Y.; Yum, D.Y.; Kim, J.H.; Kang, J.E.; Park, M.H.; Kim, E.J.; et al. SOD1 Suppresses Pro-Inflammatory Immune Responses by Protecting against Oxidative Stress in Colitis. Redox Biol. 2020, 37, 101760.
- Bruiners, N.; Dutta, N.K.; Guerrini, V.; Salamon, H.; Yamaguchi, K.D.; Karakousis, P.C.; Gennaro, M.L. The Anti-Tubercular Activity of Simvastatin Is Mediated by Cholesterol-Driven Autophagy via the AMPK-MTORC1-TFEB Axis. J. Lipid Res. 2020, 61, 1617.
- Wei, Y.M.; Li, X.; Xu, M.; Abais, J.M.; Chen, Y.; Riebling, C.R.; Boini, K.M.; Li, P.L.; Zhang, Y. Enhancement of Autophagy by Simvastatin through Inhibition of Rac1-MTOR Signaling Pathway in Coronary Arterial Myocytes. Cell. Physiol. Biochem. 2013, 31, 925–937.
- Watanabe, N.; Ikeda, U. Matrix Metalloproteinases and Atherosclerosis. Curr. Atheroscler. Rep. 2004, 6, 112–120.
- Esnault, S.; Kelly, E.A.; Johnson, S.H.; Delain, L.P.; Haedt, M.J.; Noll, A.L.; Sandbo, N.; Jarjour, N.N. Matrix Metalloproteinase-9-Dependent Release of IL-1 β by Human Eosinophils. Mediators Inflamm. 2019, 2019, 7479107.
- Yabluchanskiy, A.; Ma, Y.; Iyer, R.P.; Hall, M.E.; Lindsey, M.L. Matrix Metalloproteinase-9: Many Shades of Function in Cardiovascular Disease. Physiology 2013, 28, 391.
- Rhee, J.W.; Lee, K.W.; Kim, D.; Lee, Y.; Jeon, O.H.; Kwon, H.J.; Kim, D.S. NF-KappaB-Dependent Regulation of Matrix Metalloproteinase-9 Gene Expression by Lipopolysaccharide in a Macrophage Cell Line RAW 264.7. J. Biochem. Mol. Biol. 2007, 40, 88–94.
- Chase, A.J.; Bond, M.; Crook, M.F.; Newby, A.C. Role of Nuclear Factor-ΚB Activation in Metalloproteinase-1, -3, and -9 Secretion by Human Macrophages in Vitro and Rabbit Foam Cells Produced in Vivo. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 765–771.
- Yu, X.; Lin, S.G.; Huang, X.R.; Bacher, M.; Leng, L.; Bucala, R.; Lan, H.Y. Macrophage Migration Inhibitory Factor Induces MMP-9 Expression in Macrophages via the MEK-ERK MAP Kinase Pathway. J. Interferon Cytokine Res. 2007, 27, 103–109.
- Brosseau, C.; Colas, L.; Magnan, A.; Brouard, S. CD9 Tetraspanin: A New Pathway for the Regulation of Inflammation? Front. Immunol. 2018, 9, 2316.
- Suzuki, M.; Tachibana, I.; Takeda, Y.; He, P.; Minami, S.; Iwasaki, T.; Kida, H.; Goya, S.; Kijima, T.; Yoshida, M.; et al. Tetraspanin CD9 Negatively Regulates Lipopolysaccharide-Induced Macrophage Activation and Lung Inflammation. J. Immunol. 2009, 182, 6485–6493.
- Guha, M.; Mackman, N. LPS Induction of Gene Expression in Human Monocytes. Cell. Signal. 2001, 13, 85–94.
- Lopez-Castejon, G.; Brough, D. Understanding the Mechanism of IL-1β Secretion. Cytokine Growth Factor Rev. 2011, 22, 189.
- Franchi, L.; Eigenbrod, T.; Muñoz-Planillo, R.; Nuñez, G. The Inflammasome: A Caspase-1 Activation Platform Regulating Immune Responses and Disease Pathogenesis. Nat. Immunol. 2009, 10, 241.
- Chilton, P.M.; Embry, C.A.; Mitchell, T.C. Effects of Differences in Lipid A Structure on TLR4 Pro-Inflammatory Signaling and Inflammasome Activation. Front. Immunol. 2012, 3, 154.
- Miller, Y.I.; Choi, S.H.; Wiesner, P.; Bae, Y.S. The SYK Side of TLR4: Signalling Mechanisms in Response to LPS and Minimally Oxidized LDL. Br. J. Pharmacol. 2012, 167, 990.
- McLaren, J.E.; Ramji, D.P. Interferon Gamma: A Master Regulator of Atherosclerosis. Cytokine Growth Factor Rev. 2009, 20, 125–135.
- Yu, X.H.; Zhang, J.; Zheng, X.L.; Yang, Y.H.; Tang, C.K. Interferon-γ in Foam Cell Formation and Progression of Atherosclerosis. Clin. Chim. Acta 2015, 441, 33–43.
- Schroder, K.; Hertzog, P.J.; Ravasi, T.; Hume, D.A. Interferon-γ: An Overview of Signals, Mechanisms and Functions. J. Leukoc. Biol. 2004, 75, 163–189.
- Reith, W.; LeibundGut-Landmann, S.; Waldburger, J.M. Regulation of MHC Class II Gene Expression by the Class II Transactivator. Nat. Rev. Immunol. 2005, 5, 793–806.
- Ramgolam, V.S.; Markovic-Plese, S. Regulation of Suppressors of Cytokine Signaling as a Therapeutic Approach in Autoimmune Diseases, with an Emphasis on Multiple Sclerosis. J. Signal Transduct. 2011, 2011, 635721.
More
Information
Subjects:
Immunology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
993
Revisions:
2 times
(View History)
Update Date:
08 Jun 2022
Table of Contents




Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No