Maize (
Zea mays L.), also called corn, is one of the top three staple food crops, along with rice and wheat and is grown on more acreage than any other crop except wheat, with a current annual production of 1.16 billion metric tons (FAOSTAT 2020). Maize accounts for 20% of the calories consumed by humans daily in Africa and Mesoamerica
[1]. As the global population is expected to reach approximately 9.8 billion people by 2050, it is anticipated that worldwide maize production will need to increase to an additional 1.16 billion metric tons over the current maize harvest to meet global demand based on the current consumption rate. In addition, climate change will have a negative impact on maize production in the coming decades. For example, global warming will cut cereal crop yields by 43.5 million tons for every 1 °C increase in temperature
[2]. Further, climate change impacts, from rising temperatures to unpredictable rainfall patterns, will create favorable conditions for pathogens, allowing them to wreak havoc on crops, hence reducing grain yield and quality. Therefore, a 1.8% annual rate of genetic gain in grain yield will be required to meet the global demand for maize in the coming decades, in conjunction with improvements in disease resistance, agronomic performance and end-use quality. In addition to the staple food crop, maize is also used as animal feed (grain feed and silage), and a source for biofuel (e.g., bioethanol) and bio-based plastics. Approximately 40% of the maize produced in the United States is utilized in bioethanol production, and approximately 36% is fed to livestock (hogs, cattle and chickens)
[3].
Fungal diseases pose a significant threat to maize production worldwide, both in terms of grain yield and quality. The crop is challenged by over a dozen adapted fungal pathogens during the growing season, causing significant economic damage, including although not limited to
Fusarium verticilliodes (Fusarium ear and stalk rot),
F. graminearum (Gibberella ear rot),
Cercospora zeae-maydis (gray leaf spot),
Setosphaeria turcica (northern corn leaf blight),
Bipolaris maydis (southern corn leaf blight),
Cochliobolus carbonum (northern corn leaf spot),
Colletotrichum graminicola (anthracnose),
Diplodia maydis and D. Zeae (Diplodia ear rot)
, Ustilago maydis (common smut),
Puccinia polysora (southern corn rust),
P.
sorghi (common rust),
Sporisorium reilianum (head smut),
Rhizoctonia solani (Banded leaf and sheath blight),
Alternaria alternata (Alternaria leaf blight),
Kabatiella zeae (eyespot),
Sphacelotheca reiliana (head smut),
Physoderma maydis (Physoderma leaf spot),
Phyllachora maydis (tar spot),
Macrophomina phaseolina (Charcoal rot) and
Aspergillus flavus (Aspergillus ear rot)
[4][5]. Recently, researchers reported a new fungal pathogen
Didymella glomerata, causing Didymella leaf blight on maize in Panjin, Liaoning Province, China
[6]. Yield loss attributed to these pathogens has been estimated to be 10.6% in North America
[5] and 12% in Asia
[7].
C. graminicola (Ces.) G.W. Wilson is an ascomycete fungal pathogen that mainly infects above-ground parts of maize, causing anthracnose leaf blight (ALB;
Figure 1A) and anthracnose stalk rot (ASR;
Figure 1B) and top dieback. The pathogen was first reported to cause severe ASR in Ohio in 1963
[8]. However, anthracnose was not considered as an economically significant disease of maize until the early 1970s, when severe epidemics wiped out the sweet corn crop in the north-central and eastern United States
[9]. This sudden surge of anthracnose was perhaps attributed to the emergence of virulent races, a shift in cultural practices (e.g., widespread adoption of zero tillage or conservation tillage cropping system due to its environmental benefits) and the introduction of high-yielding cultivars lacking resistance to ALB/ASR
[10]. In the 1980s and 1990s, anthracnose became a major threat to maize production and reduced grain yield by up to 40% in the corn belt of the United States, equating to a loss of more than USD 1 billion
[11]. Today, ALB/ASR is considered as one of the top five most destructive diseases of maize in the United States and in Ontario, a major maize-producing province of Canada
[5]. Globally, ALB/ASR is one of the top 10 most devastating diseases of maize and is prevalent in 79 countries (
Figure 2).
Despite economic significance, molecular genetics underlying ASR and ALB lags behind other fungal diseases, such as ear and stalk rot caused by Fusarium spp.
2. Genetic Architecture of Anthracnose Resistance in Maize
In the absence of an adaptative resistance, genetic resistance in plants relies on the detection of conserved and variable pathogen molecules called, respectively, pathogen-associated molecular patterns (PAMPs) and effectors. The archetypical PAMPs include chitin (a major constituent of the fungal cell wall) and flagellin (a principal constituent of bacterial flagella). Effectors are proteinaceous and non-proteinaceous molecules that pathogens secrete into hosts to manipulate host metabolism and/or to disarm the immune system, thereby facilitating their colonization and proliferation. To detect PAMPs and effectors, plants possess a two-branched multilayered immune system. The first branch functions at the extracellular level and is called the PAMP-triggered immune (PTI) system, whereas the second branch, called the effector-triggered immune (ETI) system, acts at the intracellular level. The PTI system is activated when its components, transmembrane pattern recognition receptors (PRRs, e.g., receptor-like kinases and receptor-like proteins) detect PAMPs in the apoplast. Once the pathogens are able to disable the PTI system using their effector arsenal successfully, the ETI system comes into play, whereby nucleotide-binding leucine-rich repeat resistance proteins (NLRs, a major component of the ETI system) recognize variable effectors—that undermine both PTI and ETI systems and promote pathogen colonization—either directly through physical interactions
[12], or indirectly by monitoring the integrity of guarded host virulence targets (called guardees)
[13][14] or mimics thereof (called decoys)
[15] in the cytoplasm.
Z. mays (2
n = 20) is a diploid species, whose nuclear genome (2.13 Gb) encodes ~144 NLRs (
Figure 3). The number of NLRs in maize is relatively lower than those reported for other cereal crops, e.g.,
Oryza sativa (438),
Triticum aestivum (627) and
Hordeum vulgare (224)
[16].
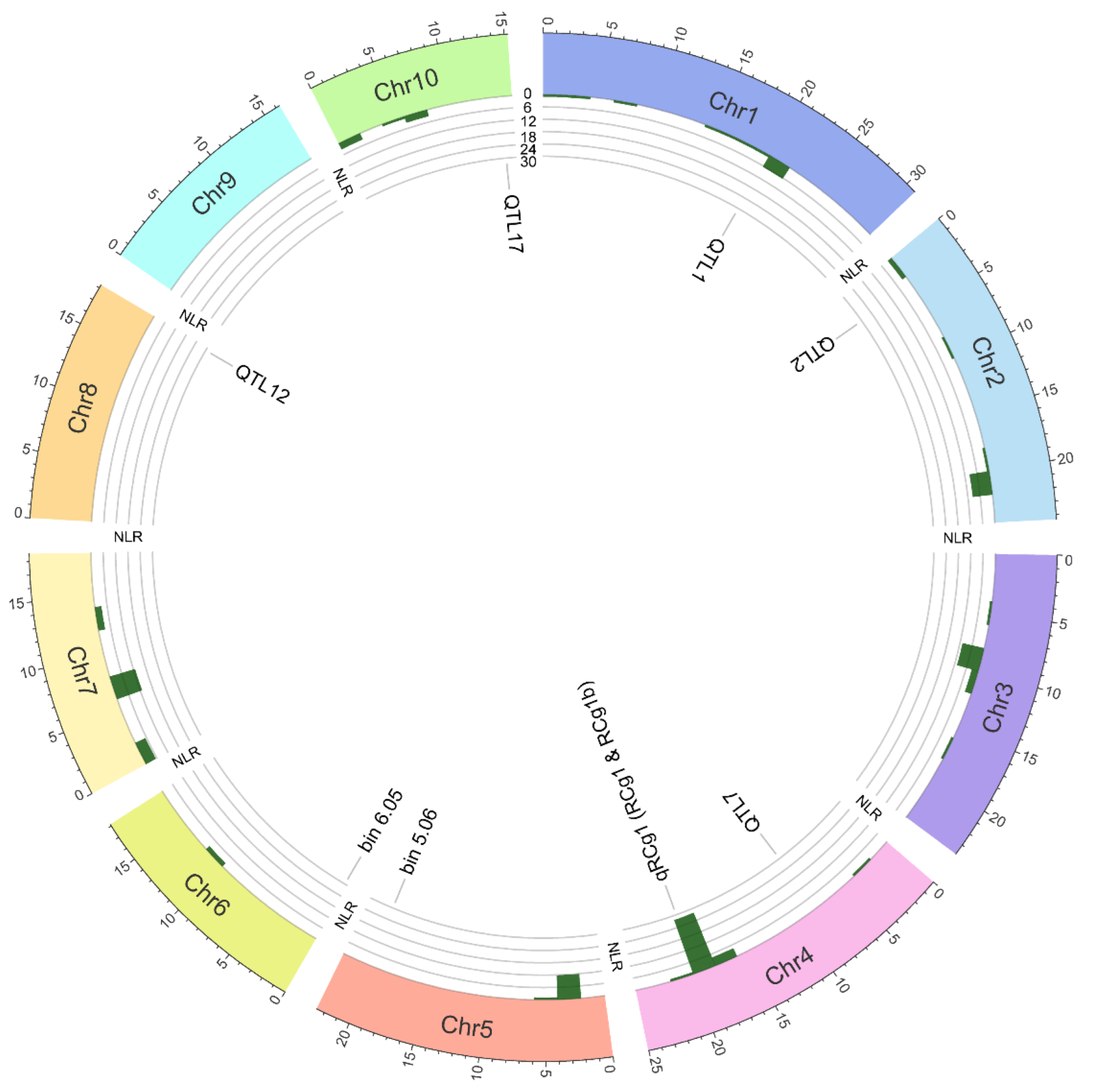 Figure 3.
Figure 3. Circos plot exemplifying the distribution of 144
NLRs (encoding nucleotide-binding leucine-rich repeat resistance proteins) and QTL (conferring resistance to anthracnose leaf blight and stalk rot) in the
Zea mays B73 genome. The outer track shows the
Z. mays ideogram, comprising ten chromosomes (Chr1 through Chr10). The middle track consists of five circular ticks (6
NLRs/tick); the bars inside the track exhibit the frequency distribution of
NLRs on the chromosomes. The inner track indicates the location of QTL based on their flanking marker positions listed in
Table 1. The B73 genome lacks the QTL
qRCg1 controlling resistance to anthracnose stalk rot; hence, its location on the B73 genome is relative to the marker UMC15a.
Perception of PAMPs by PRRs and effectors by NLRs triggers multilayered signaling, leading to, respectively, PTI and ETI responses in host tissues, such as oxidative (ROS) burst, Ca
2+ influx, upregulation of mitogen-activated protein kinases, activation of pathogenesis-related genes and phytohormone synthesis
[17]. The ETI system is very effective in guarding the plants against the biotrophic invaders that require living host tissue for nutrition and growth, or hemibiotrophic invaders, but not against necrotrophic invaders that kill and macerate living host tissues for nourishment and growth. In addition to the above-mentioned defense responses, ETI also induces localized cell death surrounding the infection site, called the hypersensitive cell death response (HR), which checks the ingress of the biotrophic and hemibiotrophic pathogens
[18].
C. graminicola is a hemibiotrophic pathogen; therefore, the ETI system may underlie anthracnose resistance in maize, i.e., anthracnose is a gene-for-gene disease in which the resistant maize genotypes and
C. graminicola recognize each other by their matching pairs of NLR protein(s) and effector(s) during the biotrophic phase of infection. If ALB/ASR is a gene-for-gene disease, it should be controlled by a single dominant resistance gene.
Over a dozen genetic sources of anthracnose resistance have been identified in maize: LB1, LB6, ECB8, L04-2, Pa91, T111 and LB-58 (ALB-resistant lines); and A556, MP305, H21, SP288, CI88A, FR16, S11, R177 and LB31 (ASR-resistant lines). Nevertheless, genetic resistance underlying anthracnose resistance is poorly understood due to the lack of genetic studies. The first study was undertaken over 40 years ago to determine the mode of inheritance of ALB and ASR resistance in maize. Lim and White
[19] generated forty-five F
1 diallel crosses originating from ten parental inbred lines (Pa91, T111 and R177 [resistant to ALB/ASR]; Mo940, Oh07B, C123 and Va26 [susceptible to ALB/ASR]; Mo17, B73 and H95 [intermediate lines]). Analysis of ALB and ASR reactions of F1 hybrids and parental lines thereof showed that ALB and ASR were not correlated traits and that the hybrids from resistant parents were more resistant than those involving intermediate or susceptible inbred lines. In addition, ALB and ASR resistance could be combined in hybrids by crossing an ALB-resistant inbred line (Pa91 or T111) with the ASR-resistant line (R177). This implies that distinct genes in maize control ALB and ASR resistance and that the resistance is likely polygenic and additive in gene action. The quantitative nature of ASR resistance was further corroborated by the analysis of reciprocal translocation testcross populations for ASR reactions. Carson and Hooker
[20] contrived 19 reciprocal translocation testcross populations and evaluated them for ASR reactions to locate chromosomal arms in the highly resistant Corn Belt inbred line A556 governing ASR resistance. These 19 reciprocal chromosomal translocation stocks were used to generate the testcross populations representing 15 out of the 20 chromosomal arms. Fourteen of the translocation stocks were in an M14 genetic background, whereas the remaining four were in the W23 genetic background. Both M14 and W23 inbred lines are highly susceptible to ASR. These nineteen stocks were crossed with A556; a susceptible tester C123 (ASR susceptible line) was also incorporated in the testcross to segregate ASR reactions and fertility/semisterility: (translocation stock ×A556) × C123. Mean differences in AR reactions between fertile and semisterile plants were used to determine which chromosomal arm confers resistance to ASR as full fertility was also a factor in enhanced resistance in addition to the chromosomal arms per se. Testcross population lines carrying the long arms of chromosomes 1, 4 and 8, and both arms of chromosome 6 showed increased ASR resistance, indicating that ASR is a polygenic trait, controlled at least by five genes. The polygenic nature of ALB was further confirmed by genetic mapping of ALB resistance in a biparental recombinant inbred line (RIL) population origination from a cross between ALB-resistant L
R04-2 and ALB-susceptible L
S95-1 inbred lines. Four out of seventeen QTL, one on chromosome 9 (QTL13;
Table 1) and three on chromosome 10 (QTL15 through 17;
Table 1 and
Figure 3) were the most stable and explained 27.7 to 54.3% of the variance in ALB severity in the RIL population
[21]. However, genes underlying the QTL controlling ALB resistance remain unknown.
A handful of studies suggest that anthracnose on maize is a gene-for-gene disease in which the resistance is controlled by a single dominant resistance gene or two dominant genes, one with major effect and the other with minor effect. Badu-Apraku et al.
[22] evaluated F
1, F
2, backcross and backcross-selfed plants originating from a cross between LB-58 and A632 maize inbred lines for ALB reactions. LB-58 shows HR following the
C. graminicola infection and thus is a highly resistant inbred line, whereas A632 is an ALB-susceptible line. All F
1 plants were resistant to ALB, suggesting that ALB is a dominant trait. The reactions to ALB in the F
2 population were segregated in a 3 (resistant) to 1 (susceptible) ratio both at the seedling and mature plant stages, indicating that ALB resistance in LB-58 is likely controlled by a single dominant resistance gene. The backcross progenies derived from the F
1 × LB-58 cross were all resistant, whereas the backcross progenies resulting from the F
1 × A632 cross were segregated for ALB reactions in a 1 (resistant): 1 (susceptible) ratio both at the seedling and mature plant stages, thereby conforming with a goodness-of-fit test for a single gene model (
p > 0.05) for the inheritance of resistance to ALB. Likewise, the segregation of the reactions of backcross-selfed populations (BC
1 and BC
2) to ALB also confirmed a single dominant resistance gene (
CgL) conferring resistance to ALB. The
CgL gene conditions HR (characterized by chlorotic flecks) during the
C. graminicola infection of LB-58, more likely at the biotrophic phase of infection, which restricts the pathogen from further colonization. Badu-Apraku et al.
[23] also discovered a single dominant resistance gene governing ASR resistance in the ASR-resistant inbred line LB-31. The researchers used a similar approach to determine the inheritance mode of ASR resistance in LB-31-derived populations. They created F
1, F
2 and backcross populations originating from a cross between LB-31 and ASR-susceptible inbred line B37. The F
1 plants were all resistant to ASR; the F
2 population was segregated for ASR reactions in a 3 (resistant) to 1 (susceptible) ratio; the progenies from F
1 × LB-31 were all resistant, whereas the progenies from F
1 × LB-31 were segregated for reactions to ASR in a 1 (resistant) to 1 (susceptible) ratio. The above observations imply that a single dominant gene (
CgR) controls ASR resistance in LB-31. However, none of the two genes (
CgL and
CgR) have been genetically or physically mapped. In a preliminary study, Carlson
[24] tracked the inheritance of ASR resistance in F
1 and F
1 × MP305 populations derived from ASR-resistant (MP305) and ASR-susceptible (A632) inbred lines. The majority of the lines in the populations had only one discolored internode due to ASR, similar to MP305; however, some individual plants had two to three discolored internodes, resembling A632, which showed a higher degree of susceptibility, i.e., the number of discolored internodes varied from two to five. The F
2 population showed a continuous distribution of ASR, albeit a high degree of skewness towards fewer discolored internodes. A two-gene model could explain the variation within the F
2 population; therefore, MP305 likely contains one major and one minor dominant resistance genes, which are likely closely linked. Toman and White
[25] delved into the inheritance of ASR resistance in the populations originating from the ASR-resistant DW1035 and ASR-susceptible FRB73 inbred lines. DW1035 is derived from a cross between MP305 and FRB73, followed by repeated backcrossing (five times) with selection for ASR resistance in each cycle
[20].
Jung et al.
[26] performed inheritance analyses of ASR resistance in F
1, F
2, F
3 and backcross populations originating from the DE811ASR × DE811 and DE811ASR × LH132 crosses. The inbred line DE811ASR is resistant to ASR and was developed from a cross between the recurrent parent DE811 and the resistant parent MP305, followed by backcrossing with DE811 three times (BC
3), whereas DE811 and LH132 are susceptible to ASR. Generation-means analysis of the populations for ASR (i.e., number of discolored internodes and number of internodes >75% discolored) showed that inheritance of ASR resistance is largely additive in the populations. The researchers were able to map a major QTL
qRcg1 on the long arm of chromosome 4 that conferred ASR resistance in F
2 and F
3 populations (
Table 1 and
Figure 3).
Table 1. QTL in maize conferring resistance to Anthracnose stalk rot and leaf blight caused by Colletotrichum graminicola.
| Population |
Resistance Source |
Population |
QTL |
LG |
Linked Markers |
Marker Interval |
ASR/ALB |
Reference |
| DE811ASR × DE811 |
DE811ASR (MP305) |
RIL |
RCg1 |
4 |
UMC66a-UMC15a |
397.4–525.8 cM |
ASR |
[26] |
| DE811ASR × LH132 |
DE811ASR (MP305) |
RIL |
RCg1 |
4 |
UMC66a-UMC15a |
397.4–525.8 cM |
ASR |
[26] |
| DE811ASR × DE811 |
DE811ASR (MP305) |
NIL |
RCg1 |
4 |
MZA2591-PHI093 |
61.0–63.0 cM |
ASR |
[27][28] |
| S11 × DK8883 |
S11 |
F6:7 HIF |
bin 5.06 |
5 |
umc2216 |
518.4 cM |
ASR |
[29] |
| |
S11 |
F6:7 HIF |
bin 6.05 |
6 |
bngl2249 |
278.0 cM |
ASR |
[29] |
| LB58 × A632 |
LB58 |
BC |
CgL |
|
- |
- |
ALB |
[22] |
| LB31 × B37 |
LB31 |
RIL and BC |
CgR |
|
- |
|
ASR |
[23] |
| L04-2 × L95-1 |
L04-2 |
RIL |
QTL1 |
1 |
E32M48_308-E42M50_174 |
177.9–189.4 cM |
ALB |
[21] |
| |
L04-2 |
RIL |
QTL2 |
2 |
E35M56_680-E35M56_112 |
0.0–14.1 cM |
ALB |
|
| |
L04-2 |
RIL |
QTL3 |
3 |
E42M51_162-E42M50_76 |
0.0–7.6 cM |
ALB |
|
| |
L04-2 |
RIL |
QTL4 |
3 |
E32M48_167-E32M59_104 |
51.0–61.4 cM |
ALB |
|
| |
L04-2 |
RIL |
QTL5 |
4 |
E35M60_87-E32M60_185 |
0.0–10.4 cM |
ALB |
|
| |
L04-2 |
RIL |
QTL6 |
4 |
E32M52_73-E44M51_84 |
15.3–34.8 cM |
ALB |
|
| |
L04-2 |
RIL |
QTL7 |
4 |
Umc1511-E32M53_434 |
88.1–119.3 cM |
ALB |
|
| |
L04-2 |
RIL |
QTL8 |
4 |
E32M53_434-E44M51_135 |
119.3–137.7 cM |
ALB |
|
| |
L04-2 |
RIL |
QTL9 |
5 |
E32M48_532-E32M50_139 |
242.0–244.3 cM |
ALB |
|
| |
L04-2 |
RIL |
QTL10 |
8 |
E35M60_80-E32M50_100 |
0.0–23.9 cM |
ALB |
|
| |
L04-2 |
RIL |
QTL11 |
8 |
E32M60_94-E32M50_248 |
57.3–74.1 cM |
ALB |
|
| |
L04-2 |
RIL |
QTL12 |
8 |
E35M60_86-Phi015 |
85.5–107.5 cM |
ALB |
|
| |
L04-2 |
RIL |
QTL13S |
9 |
E32M48_562-E32M48_97 |
126.4–157.7 cM |
ALB |
|
| |
L04-2 |
RIL |
QTL14 |
9 |
E32M51_314-E35M56_174 |
179.1–201.1 cM |
ALB |
|
| |
L04-2 |
RIL |
QTL15S |
10 |
E32M49_698-E32M59_207 |
28.3–58.7 cM |
ALB |
|
| |
L04-2 |
RIL |
QTL16S |
10 |
E32M50_118-E44M56_81 |
85.4–109.1 cM |
ALB |
|
| |
L04-2 |
RIL |
QTL17S |
10 |
E32M59_76-Umc1084 |
161.6–191.7 cM |
ALB |
 +1 credit
+1 credit
 Figure 1. Symptoms caused by Colletotrichum graminicola on the susceptible Zea mays inbred line B73. (A) Gray to brown oval necrotic lesions on the B73 leaves are the typical symptom of anthracnose leaf blight. The blight lesions contain dot-like black structures called microsclerotia, which serve as the primary source of inoculum in the next growing season. (B) Discoloration of the pith (rotting pith) is the typical symptom of anthracnose stalk rot. The rotten pith also leads to bleaching of the upper part of the maize plants (top dieback). (C) Heathy pith.
Figure 1. Symptoms caused by Colletotrichum graminicola on the susceptible Zea mays inbred line B73. (A) Gray to brown oval necrotic lesions on the B73 leaves are the typical symptom of anthracnose leaf blight. The blight lesions contain dot-like black structures called microsclerotia, which serve as the primary source of inoculum in the next growing season. (B) Discoloration of the pith (rotting pith) is the typical symptom of anthracnose stalk rot. The rotten pith also leads to bleaching of the upper part of the maize plants (top dieback). (C) Heathy pith.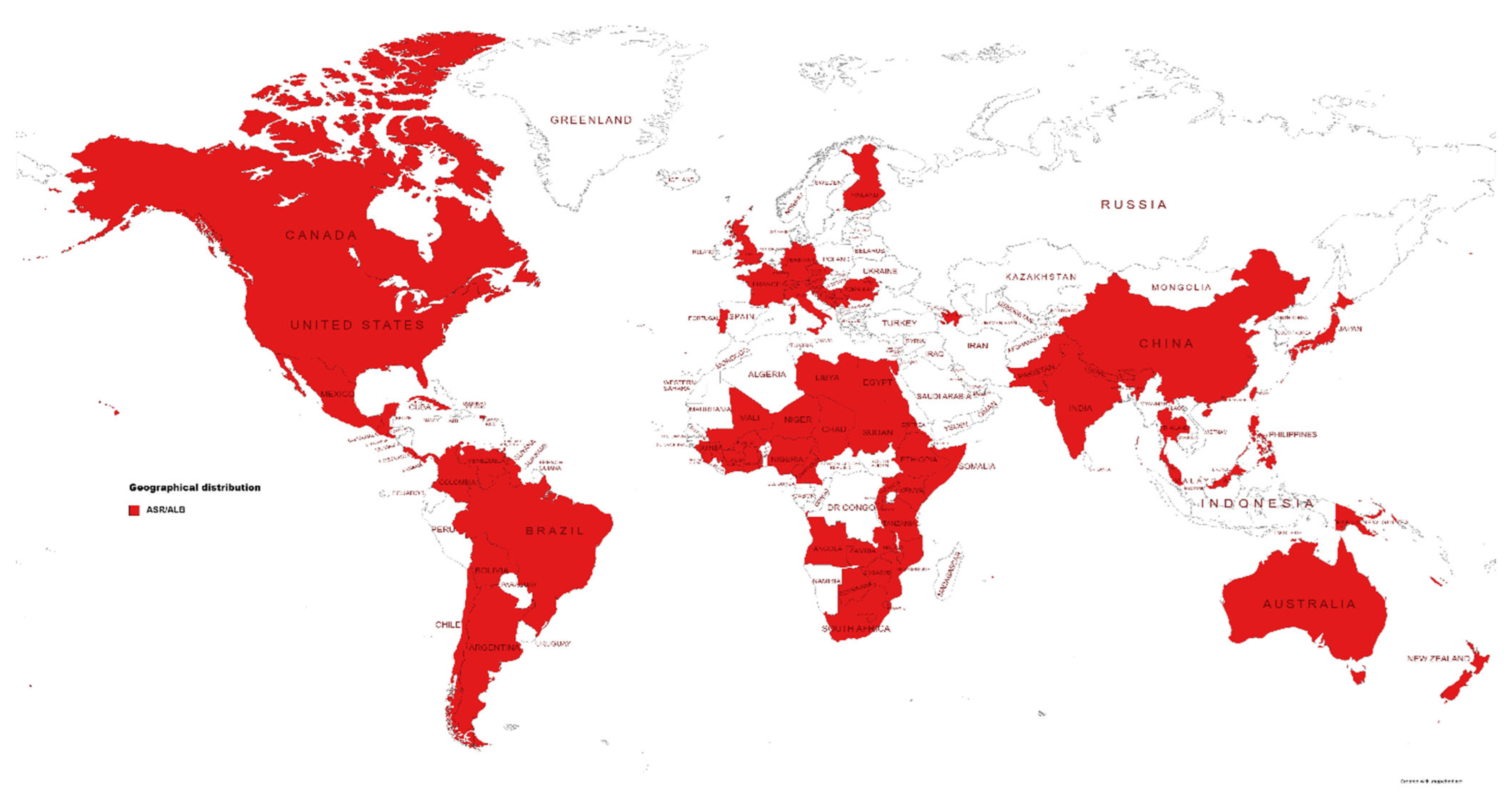 Figure 2. Global distribution of anthracnose leaf blight (ALB) and anthracnose stalk rot (ASR).
Figure 2. Global distribution of anthracnose leaf blight (ALB) and anthracnose stalk rot (ASR).
