 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Rafael Rincon Perez | -- | 2842 | 2022-05-30 15:37:31 | | | |
| 2 | Peter Tang | Meta information modification | 2842 | 2022-05-31 03:13:35 | | |
Video Upload Options
Multiple myeloma (MM) accounts for about 10% of hematological malignancies. It is a plasma cell malignancy that originates from the post-germinal lymphoid B-cell lineage, and is characterized by an uncontrolled clonal growth of plasma cells. The discovery of non-coding RNAs as key actors in multiple myeloma has broadened the molecular landscape of this disease, together with classical epigenetic factors such as methylation and acetylation. microRNAs and long non-coding RNAs comprise the majority of the described non-coding RNAs dysregulated in multiple myeloma, while circular RNAs are recently emerging as promising molecular targets.
1. Introduction
2. Methylation
3. Acetylation
4. Non-Coding RNAs
4.1. microRNAs
|
Activity/Pathway Affected |
miRNA |
Status 1 |
Target |
References |
|---|---|---|---|---|
|
Enhances PI3K/Akt pathway |
miR-20a |
|
EGR2, PTEN |
|
|
miR-21 |
|
PIAS3 |
[24] |
|
|
miR-25-3p |
|
PTEN |
[25] |
|
|
miR-221/222 |
|
PUMA, PTEN, CDKN1B, p27 |
[26] |
|
|
miR-410 |
|
KLF10 |
[27] |
|
|
Enhances mTOR pathway |
miR-19b |
|
TSC1 |
|
|
miR-135b, miR-642a |
|
DEPTOR |
[30] |
|
|
Related to a hypoxia phenotype |
miR-210 |
|
DIMT1 |
|
|
miR-1305 |
|
MDM2, IGF1, FGF2 |
[33] |
|
|
Disrupts PRC2 activity |
miR-124 |
|
EZH2 |
[34] |
|
Modulates microenvironment |
miR-146a |
|
Not described |
[35] |
|
miR-155 |
|
Not described |
[36] |
|
|
Promotes proliferation, circulating miRNAs |
miR-17-92 |
|
BIM |
[28] |
|
miR-221/222 |
|
[26] |
||
|
Circulating miRNA |
miR-1 |
|
Not described |
[37] |
|
miR-133a/b |
|
Not described |
[37] |
|
|
miR-135b |
|
HIF1A |
||
|
miR-146b |
|
Not described |
[40] |
|
|
miR-181a |
|
BCL2L11 |
||
|
miR-214 |
|
CD276 |
[43] |
|
|
Represses JAK/STAT pathway |
miR-125b |
|
IL6R, STAT3, MALAT1 |
|
|
miR-331-3p |
|
IL6R |
[46] |
|
|
miR-375 |
|
PDPK1 |
[47] |
|
|
miR-451 |
|
IL6R |
[48] |
|
|
let-7b-5p |
|
IGF1R |
[49] |
|
|
Regulates cyclin activity |
miR-26a |
|
CDK6 |
[50] |
|
miR-28-5p |
|
CCND1 |
[51] |
|
|
miR-30a-3p |
|
MAF |
[52] |
|
|
miR-338-3p |
|
CDK4 |
[53] |
|
|
miR-340-5p |
|
CCND1, NRAS |
[54] |
|
|
miR-196a/b |
|
CCND2 |
[37] |
|
|
Regulates proliferation |
miR-22 |
|
c-Myc |
[55] |
|
miR-29a |
|
c-Myc |
[56] |
|
|
miR-34a |
|
BCL2, CDK6, NOTCH1, c-Myc, MET, IL6R |
||
|
miR-193a |
|
MCL1 |
[59] |
|
|
miR-497 |
|
BCL2 |
[60] |
|
|
miR-767-5p |
|
MAPK4 |
[61] |
|
|
miR-874-3p |
|
HDAC1 |
[62] |
|
|
miR-1180 |
|
YAP |
[63] |
|
|
Prevents angiogenesis |
miR-15a/16 |
|
BCL2, VEGF, IL17 |
[64] |
|
Regulates acetylation |
miR-29b |
|
HDAC4, MCL1 |
[65] |
|
Regulates transcriptional activity |
miR-509-5p |
|
FOXP1 |
[66] |
|
miR-1271-5p |
|
SOX13, HGF |
||
|
Prevents hypoxia phenotype |
miR-199a-5p |
|
HIF1A, VEGFA |
[69] |
|
Prevents osteolytic activity |
miR-342 |
|
RUNX2 |
[70] |
|
miR-363 |
|
RUNX2 |
[70] |


1 Arrow up indicates overexpression of the miRNA, and arrow down indicates underexpression of the miRNA.
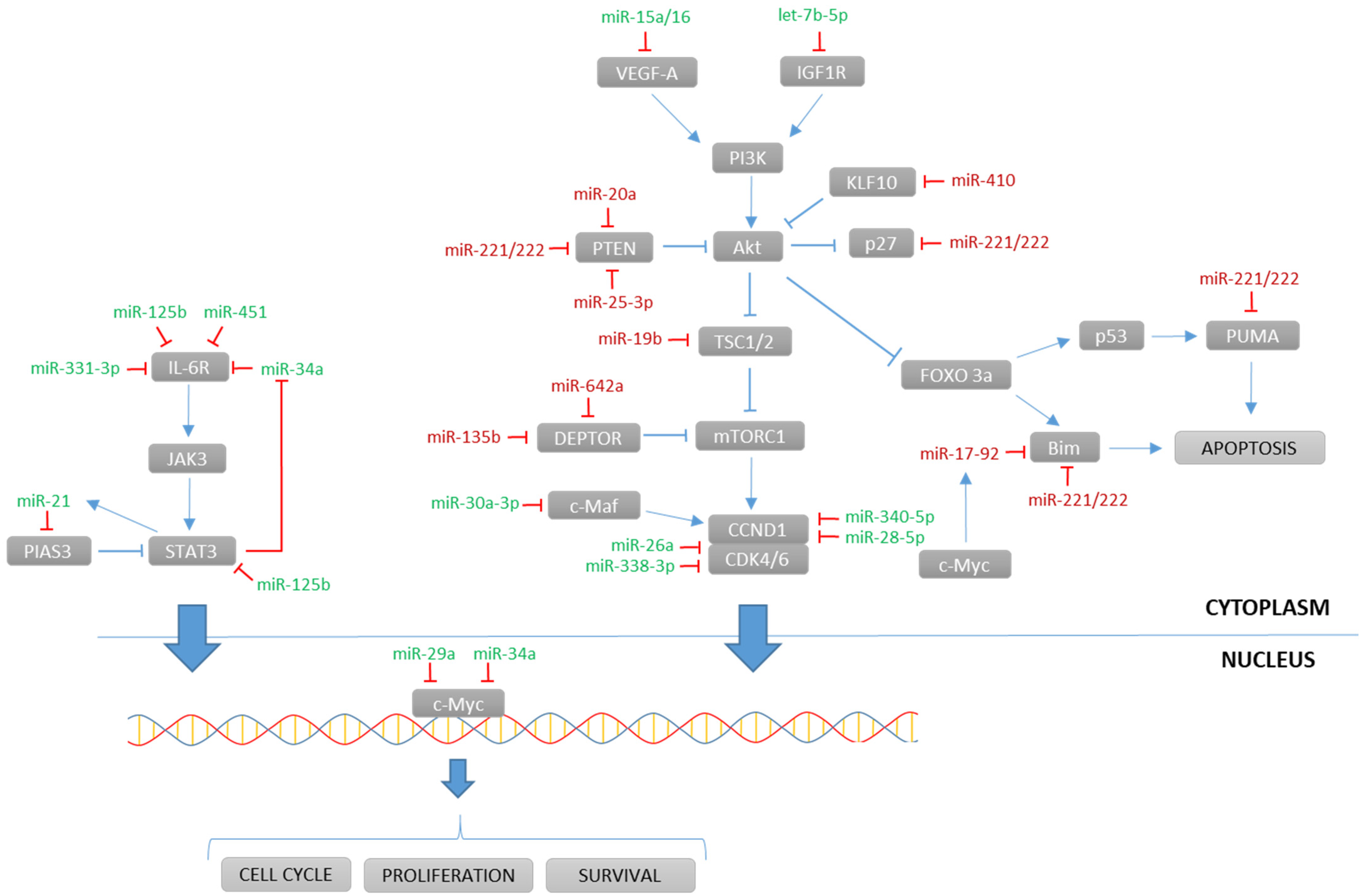
4.2. Long Non-Coding RNAs
|
lncRNA |
Status 1 |
Target |
Activity/Pathway Affected |
References |
|---|---|---|---|---|
|
ANGPLT1-3 |
|
miR-30a-3p |
ceRNA |
[52] |
|
BM742401 |
|
Not described |
Inhibit myeloma cell migration, biomarker |
[73] |
|
CRNDE |
|
miR-451 |
ceRNA |
|
|
DARS-AS1 |
|
RBM39 |
Enhances mTOR pathway, hypoxia phenotype |
[75] |
|
H19 |
|
miR-29b |
ceRNA, biomarker |
[76] |
|
HOTAIR |
|
Not described |
Enhances JAK/STAT pathway |
[77] |
|
MALAT1 |
|
HMGB1, miR-509-5p, miR-1271 |
Contributes to genomic stability, ceRNA, biomarker |
|
|
MEG3 |
|
miR-181a |
Promotes osteogenic differentiation, biomarker, ceRNA |
[80] |
|
MIAT |
|
miR-29b |
Inducible by bortezomib, ceRNA, biomarker |
[81] |
|
NEAT1 |
|
miR-214, miR-193a |
Downregulates genes involved in DNA repair, enhances Wnt/β-catenin pathway, ceRNA |
|
|
NR_046683 |
|
Not described |
Biomarker |
[84] |
|
OPI5-AS1 |
|
miR-410 |
ceRNA |
[27] |
|
PDIA3P |
|
c-Myc |
Regulates proliferation |
[85] |
|
RUNX2-AS1 |
|
RUNX2 pre-mRNA |
Promotes osteogenesis |
[86] |
|
SMILO |
|
Not described |
Regulates proliferation |
[87] |
|
SNHG16 |
|
miR-342 |
ceRNA |
[88] |
|
UCA1 |
|
miR-1271-5p, miR-331-3p |
ceRNA |
|
|
XLOC_013703 |
|
IKKA |
Represses NF-κB pathway |
[89] |


|
lncRNA |
miRNA |
Gene |
References |
|---|---|---|---|
|
ANGPLT1-3 |
miR-30a-3p |
MAF |
[52] |
|
CRNDE |
miR-451 |
IL6R |
|
|
H19 |
miR-29b |
HDAC4 and MCL1 |
|
|
MALAT1 |
miR-509-5p |
FOXP1 |
[66] |
|
miR-1271-5p |
SOX13 |
[67] |
|
|
MEG3 |
miR-181a |
BCL2L11 |
[80] |
|
MIAT |
miR-29b |
HDAC4 and MCL1 |
|
|
NEAT1 |
miR-214 |
CD276 |
[43] |
|
miR-193a |
MCL1 |
[59] |
|
|
OPI5-AS1 |
miR-410 |
KLF10 |
[27] |
|
PRAL |
miR-210 |
DIMT1 |
|
|
SNHG16 |
miR-342 |
RUNX2 |
[88] |
|
UCA1 |
miR-331-3p |
IL6R |
[54] |
|
miR-1271-5p |
SOX13 and HGF |
[68] |
4.3. Other ncRNAs
5. Conclusions
References
- Kyle, R.A.; Therneau, T.M.; Rajkumar, S.V.; Larson, D.R.; Plevak, M.F.; Offord, J.R.; Dispenzieri, A.; Katzmann, J.A.; Melton, L.J., 3rd. Prevalence of monoclonal gammopathy of undetermined significance. N. Engl. J. Med. 2006, 354, 1362–1369.
- Laubach, J.P.; Richardson, P.G.; Anderson, K.C. The evolution and impact of therapy in multiple myeloma. Med. Oncol. 2010, 27 (Suppl. S1), 1–6.
- Cheung, H.H.; Lee, T.-L.; Rennert, O.M.; Chan, W.Y. DNA methylation of cancer genome. Birth Defects Res. Part C Embryo Today Rev. 2009, 87, 335–350.
- Dimopoulos, K.; Gimsing, P.; Grønbæk, K. The role of epigenetics in the biology of multiple myeloma. Blood Cancer J. 2014, 4, e207.
- Rasmussen, K.D.; Helin, K. Role of TET enzymes in DNA methylation, development, and cancer. Genes Dev. 2016, 30, 733–750.
- Tsiouplis, N.J.; Bailey, D.W.; Chiou, L.F.; Wissink, F.J.; Tsagaratou, A. TET-mediated epigenetic regulation in immune cell development and disease. Front. Cell Dev. Biol. 2020, 8, 623948.
- Walker, B.A.; Wardell, C.P.; Chiecchio, L.; Smith, E.M.; Boyd, K.D.; Neri, A.; Davies, F.E.; Ross, F.M.; Morgan, G.J. Aberrant global methylation patterns affect the molecular pathogenesis and prognosis of multiple myeloma. Blood 2011, 117, 553–562.
- Agirre, X.; Castellano, G.; Pascual, M.; Heath, S.; Kulis, M.; Segura, V.; Bergmann, A.; Esteve, A.; Merkel, A.; Raineri, E.; et al. Whole-epigenome analysis in multiple myeloma reveals DNA hypermethylation of B cell-specific enhancers. Genome Res. 2015, 25, 478–487.
- Kaiser, M.F.; Johnson, D.C.; Wu, P.; Walker, B.A.; Brioli, A.; Mirabella, F.; Wardell, C.P.; Melchor, L.; Davies, F.E.; Morgan, G.J. Global methylation analysis identifies prognostically important epigenetically inactivated tumor suppressor genes in multiple myeloma. Blood 2013, 122, 219–226.
- Martínez-Baños, D.; Sánchez-Hernández, B.; Jimenez, G.; Barrera-Lumbreras, G.; Barrales-Benítez, O. Global methylation and promoter-specific methylation of the P16, SOCS-1, E-cadherin, P73 and SHP-1 genes and their expression in patients with multiple myeloma during active disease and remission. Exp. Ther. Med. 2017, 13, 2442–2450.
- Chong, P.S.Y.; Chng, W.-J.; de Mel, S. STAT3: A promising therapeutic target in multiple myeloma. Cancers 2019, 11, 731.
- Kiziltepe, T.; Hideshima, T.; Catley, L.; Raje, N.; Yasui, H.; Shiraishi, N.; Okawa, Y.; Ikeda, H.; Vallet, S.; Pozzi, S.; et al. 5-azacytidine, a DNA methyltransferase inhibitor, induces ATR-mediated DNA double-strand break responses, apoptosis and synergistic cytotoxicity with doxorubicin and bortezomib against multiple myeloma cells. Mol. Cancer Ther. 2007, 6, 1718–1727.
- U.S National Institutes of Health-ClinicalTrials.Gov. Available online: www.clinicaltrials.gov (accessed on 1 September 2020).
- Dutta, R.; Tiu, B.; Sakamoto, K.M. CBP/p300 acetyltransferase activity in hematologic malignancies. Mol. Genet. Metab. 2016, 119, 37–43.
- Zhu, Y.X.; Shi, C.-X.; Bruins, L.A.; Wang, X.; Riggs, D.L.; Porter, B.; Ahmann, J.M.; de Campos, C.B.; Braggio, E.; Bergsagel, P.L.; et al. Identification of lenalidomide resistance pathways in myeloma and targeted resensitization using cereblon replacement, inhibition of STAT3 or targeting of IRF4. Blood Cancer J. 2019, 9, 19.
- Falkenberg, K.J.; Johnstone, R.W. Histone deacetylases and their inhibitors in cancer, neurological diseases and immune disorders. Nat. Rev. Drug Discov. 2014, 13, 673–691.
- Bradner, J.E.; West, N.; Grachan, M.L.; Greenberg, E.F.; Haggarty, S.J.; Warnow, T.; Mazitschek, R. Chemical phylogenetics of histone deacetylases. Nat. Chem. Biol. 2010, 6, 238–243.
- Imai, Y.; Hirano, M.; Kobayashi, M.; Futami, M.; Tojo, A. HDAC inhibitors exert anti-myeloma effects through multiple modes of action. Cancers 2019, 11, 475.
- Hideshima, T.; Bradner, J.E.; Wong, J.; Chauhan, D.; Richardson, P.; Schreiber, S.L.; Anderson, K.C. Small-molecule inhibition of proteasome and aggresome function induces synergistic antitumor activity in multiple myeloma. Proc. Natl. Acad. Sci. USA 2005, 102, 8567–8572.
- Gupta, S.C.; Tripathi, Y.N. Potential of long non-coding RNAs in cancer patients: From biomarkers to therapeutic targets. Int. J. Cancer 2017, 140, 1955–1967.
- Su, M.; Xiao, Y.; Ma, J.; Tang, Y.; Tian, B.; Zhang, Y.; Li, X.; Wu, Z.; Yang, D.; Zhou, Y.; et al. Circular RNAs in cancer: Emerging functions in hallmarks, stemness, resistance and roles as potential biomarkers. Mol. Cancer 2019, 18, 90.
- Wang, T.; Tao, W.; Zhang, L.; Li, S. Oncogenic role of microRNA-20a in human multiple myeloma. Onco Targets Ther. 2017, 10, 4465–4474.
- Jiang, Y.; Chang, H.; Chen, G. Effects of microRNA-20a on the proliferation, migration and apoptosis of multiple myeloma via the PTEN/PI3K/AKT signaling pathway. Oncol. Lett. 2018, 15, 10001–10007.
- Xiong, Q.; Zhong, Q.; Zhang, J.; Yang, M.; Li, C.; Zheng, P.; Bi, L.-J.; Ge, F. Identification of novel miR-21 target proteins in multiple myeloma cells by quantitative proteomics. J. Proteome Res. 2012, 11, 2078–2090.
- Zi, Y.; Zhang, Y.; Wu, Y.; Zhang, L.; Yang, R.; Huang, Y. Downregulation of microRNA-25-3p inhibits the proliferation and promotes the apoptosis of multiple myeloma cells via targeting the PTEN/PI3K/AKT signaling pathway. Int. J. Mol. Med. 2021, 47, 8.
- Song, J.; Ouyang, Y.; Che, J.; Li, X.; Zhao, Y.; Yang, K.; Zhao, X.; Chen, Y.; Fan, C.; Yuan, W. Potential value of miR-221/222 as diagnostic, prognostic, and therapeutic biomarkers for diseases. Front. Immunol. 2017, 8, 56.
- Yang, N.; Chen, J.; Zhang, H.; Wang, X.; Yao, H.; Peng, Y.; Zhang, W. LncRNA OIP5-AS1 loss-induced microRNA-410 accumulation regulates cell proliferation and apoptosis by targeting KLF10 via activating PTEN/PI3K/AKT pathway in multiple myeloma. Cell Death Dis. 2017, 8, e2975.
- Chen, L.; Li, C.; Zhang, R.; Gao, X.; Qu, X.; Zhao, M.; Qiao, C.; Xu, J.; Li, J. miR-17-92 cluster microRNAs confers tumorigenicity in multiple myeloma. Cancer Lett. 2011, 309, 62–70.
- Wang, N.; Liang, X.; Yu, W.; Zhou, S.; Fang, M. Differential expression of microRNA-19b promotes proliferation of cancer stem cells by regulating the TSC1/mTOR signaling pathway in multiple myeloma. Cell. Physiol. Biochem. 2018, 50, 1804–1814.
- Quwaider, D.; Corchete, L.A.; Misiewicz-Krzeminska, I.; Sarasquete, M.E.; Pérez, J.J.; Krzeminski, P.; Puig, N.; Mateos, M.V.; García-Sanz, R.; Herrero, A.B.; et al. DEPTOR maintains plasma cell differentiation and favorably affects prognosis in multiple myeloma. J. Hematol. Oncol. 2017, 10, 92.
- Saba, F.; Soleimani, M.; Abroun, S. New role of hypoxia in pathophysiology of multiple myeloma through miR-210. EXCLI J. 2018, 17, 647–662.
- Ikeda, S.; Kitadate, A.; Abe, F.; Saitoh, H.; Michishita, Y.; Hatano, Y.; Kawabata, Y.; Kitabayashi, A.; Teshima, K.; Kume, M.; et al. Hypoxia-inducible microRNA-210 regulates the DIMT1-IRF4 oncogenic axis in multiple myeloma. Cancer Sci. 2017, 108, 641–652.
- Lee, J.Y.; Ryu, D.; Lim, S.W.; Ryu, K.J.; Choi, M.E.; Yoon, S.E.; Kim, K.; Park, C.; Kim, S.J. Exosomal miR-1305 in the oncogenic activity of hypoxic multiple myeloma cells: A biomarker for predicting prognosis. J. Cancer 2021, 12, 2825–2834.
- Takanlu, J.S.; Fard, A.A.; Mohammdi, S.; Rad, S.M.A.H.; Abroun, S.; Nikbakht, M. Indirect tumor inhibitory effects of microRNA-124 through targeting EZH2 in the multiple myeloma cell line. Cell J. 2020, 22, 23–29.
- De Veirman, K.; Wang, J.; Xu, S.; Leleu, X.; Himpe, E.; Maes, K.; De Bruyne, E.; Van Valckenborgh, E.; Vanderkerken, K.; Menu, E.; et al. Induction of miR-146a by multiple myeloma cells in mesenchymal stromal cells stimulates their pro-tumoral activity. Cancer Lett. 2016, 377, 17–24.
- Gao, X.; Zhou, J.; Wang, J.; Dong, X.; Chang, Y.; Jin, Y. Mechanism of exosomal miR-155 derived from bone marrow mesenchymal stem cells on stemness maintenance and drug resistance in myeloma cells. J. Orthop. Surg. Res. 2021, 16, 637.
- Gutierrez, N.C.; Sarasquete, M.E.; Misiewicz-Krzeminska, I.; Delgado, M.; De Las Rivas, J.; Ticona, F.V.; Fermiñán, E.; Martín-Jiménez, P.; Chillon, C.; Risueño, A.; et al. Deregulation of microRNA expression in the different genetic subtypes of multiple myeloma and correlation with gene expression profiling. Leukemia 2010, 24, 629–637.
- Hao, M.; Zang, M.; Zhao, L.; Deng, S.; Xu, Y.; Qi, F.; An, G.; Qin, Y.; Sui, W.; Li, F.; et al. Serum high expression of miR-214 and miR-135b as novel predictor for myeloma bone disease development and prognosis. Oncotarget 2016, 7, 19589–19600.
- Umezu, T.; Tadokoro, H.; Azuma, K.; Yoshizawa, S.; Ohyashiki, K.; Ohyashiki, J.H. Exosomal miR-135b shed from hypoxic multiple myeloma cells enhances angiogenesis by targeting factor-inhibiting HIF-1. Blood 2014, 124, 3748–3757.
- Bao, Y.; Wei, M.; Ji, X. microRNA-146b overexpression associates with deteriorated clinical characteristics, increased International Staging System stage, cacoethic chromosome abnormality, and unfavorable prognosis in multiple myeloma patients. J. Clin. Lab. Anal. 2020, 34, e23168.
- Peng, J.; Thakur, A.; Zhang, S.; Dong, Y.; Wang, X.; Yuan, R.; Zhang, K.; Guo, X. Expressions of miR-181a and miR-20a in RPMI8226 cell line and their potential as biomarkers for multiple myeloma. Tumor Biol. 2015, 36, 8545–8552.
- Papadimitriou, M.-A.; Papanota, A.-M.; Adamopoulos, P.G.; Pilala, K.-M.; Liacos, C.-I.; Malandrakis, P.; Mavrianou-Koutsoukou, N.; Patseas, D.; Eleutherakis-Papaiakovou, E.; Gavriatopoulou, M.; et al. miRNA-seq and clinical evaluation in multiple myeloma: miR-181a overexpression predicts short-term disease progression and poor post-treatment outcome. Br. J. Cancer 2022, 126, 79–90.
- Gao, Y.; Fang, P.; Li, W.-J.; Zhang, J.; Wang, G.-P.; Jiang, D.-F.; Chen, F.-P. LncRNA NEAT1 sponges miR-214 to regulate M2 macrophage polarization by regulation of B7-H3 in multiple myeloma. Mol. Immunol. 2020, 117, 20–28.
- Gao, D.; Xiao, Z.; Li, H.-P.; Han, D.-H.; Zhang, Y.-P. The mechanism study of miR-125b in occurrence and progression of multiple myeloma. Cancer Med. 2018, 7, 134–145.
- Misso, G.; Zarone, M.R.; Lombardi, A.; Grimaldi, A.; Cossu, A.M.; Ferri, C.; Russo, M.; Vuoso, D.C.; Luce, A.; Kawasaki, H.; et al. miR-125b upregulates miR-34a and sequentially activates stress adaption and cell death mechanisms in multiple myeloma. Mol. Ther.-Nucleic Acids 2019, 16, 391–406.
- Li, J.-L.; Liu, X.-L.; Guo, S.-F.; Yang, Y.; Zhu, Y.-L.; Li, J.-Z. Long noncoding RNA UCA1 regulates proliferation and apoptosis in multiple myeloma by targeting miR-331-3p/IL6R axis for the activation of JAK2/STAT3 pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 9238–9250.
- Tatekawa, S.; Chinen, Y.; Ri, M.; Narita, T.; Shimura, Y.; Matsumura-Kimoto, Y.; Tsukamoto, T.; Kobayashi, T.; Kawata, E.; Uoshima, N.; et al. Epigenetic repression of miR-375 is the dominant mechanism for constitutive activation of the PDPK1/RPS6KA3 signalling axis in multiple myeloma. Br. J. Haematol. 2017, 178, 534–546.
- David, A.; Zocchi, S.; Talbot, A.; Choisy, C.; Ohnona, A.; Lion, J.; Cuccuini, W.; Soulier, J.; Arnulf, B.; Bories, J.-C.; et al. The long non-coding RNA CRNDE regulates growth of multiple myeloma cells via an effect on IL6 signalling. Leukemia 2021, 35, 1710–1721.
- Xu, H.; Liu, C.; Zhang, Y.; Guo, X.; Liu, Z.; Luo, Z.; Chang, Y.; Liu, S.; Sun, Z.; Wang, X. Let-7b-5p regulates proliferation and apoptosis in multiple myeloma by targeting IGF1R. Acta Biochim. Biophys. Sin. 2014, 46, 965–972.
- Xu, Y.-Y.; Song, Y.-Q.; Huang, Z.-M.; Zhang, H.-B.; Chen, M. microRNA-26a inhibits multiple myeloma cell growth by suppressing cyclin-dependent kinase 6 expression. Kaohsiung J. Med. Sci. 2019, 35, 277–283.
- Li, Z.; Wong, K.Y.; Chan, G.C.-F.; Chim, C.S. Epigenetic silencing of LPP/miR-28 in multiple myeloma. J. Clin. Pathol. 2018, 71, 253–258.
- Nian, F.; Zhu, J.; Chang, H. Long non-coding RNA ANGPTL1-3 promotes multiple myeloma bortezomib resistance by sponging miR-30a-3p to activate c-Maf expression. Biochem. Biophys. Res. Commun. 2019, 514, 1140–1146.
- Cao, Y.; Shi, X.; Liu, Y.; Xu, R.; Ai, Q. microRNA-338-3p inhibits proliferation and promotes apoptosis of multiple myeloma cells through targeting cyclin-dependent kinase 4. Oncol. Res. 2018, 27, 117–124.
- Li, Z.; Wong, K.Y.; Calin, G.A.; Chng, W.-J.; Chan, G.C.-F.; Chim, C.S. Epigenetic silencing of miR-340-5p in multiple myeloma: Mechanisms and prognostic impact. Clin. Epigenetics 2019, 11, 71.
- Caracciolo, D.; Riillo, C.; Juli, G.; Scionti, F.; Todoerti, K.; Polerà, N.; Grillone, K.; Fiorillo, L.; Arbitrio, M.; Di Martino, M.T.; et al. miR-22 modulates lenalidomide activity by counteracting MYC addiction in multiple myeloma. Cancers 2021, 13, 4365.
- Saha, M.N.; Abdi, J.; Yang, Y.; Chang, H. miRNA-29a as a tumor suppressor mediates PRIMA-1Met-induced anti-myeloma activity by targeting c-Myc. Oncotarget 2016, 7, 7149–7160.
- Di Martino, M.T.; Leone, E.; Amodio, N.; Foresta, U.; Lionetti, M.; Pitari, M.R.; Cantafio, M.E.G.; Gullà, A.; Conforti, F.; Morelli, E.; et al. Synthetic miR-34a mimics as a novel therapeutic agent for multiple myeloma: In Vitro and In Vivo evidence. Clin. Cancer Res. 2012, 18, 6260–6270.
- Misso, G.; Di Martino, M.T.; De Rosa, G.; Farooqi, A.A.; Lombardi, A.; Campani, V.; Zarone, M.R.; Gullà, A.; Tagliaferri, P.; Tassone, P.; et al. miR-34: A new weapon against cancer? Mol. Ther.-Nucleic Acids 2014, 3, e194.
- Wu, Y.; Wang, H. LncRNA NEAT1 promotes dexamethasone resistance in multiple myeloma by targeting miR-193a/MCL1 pathway. J. Biochem. Mol. Toxicol. 2018, 32, e22008.
- Tian, F.; Zhan, Y.; Zhu, W.; Li, J.; Tang, M.; Chen, X.; Jiang, J. microRNA-497 inhibits multiple myeloma growth and increases susceptibility to bortezomib by targeting Bcl-2. Int. J. Mol. Med. 2019, 43, 1058–1066.
- Feng, Y.; Zhang, L.; Wu, J.; Khadka, B.; Fang, Z.; Gu, J.; Tang, B.; Xiao, R.; Pan, G.; Liu, J. CircRNA circ_0000190 inhibits the progression of multiple myeloma through modulating miR-767-5p/MAPK4 pathway. J. Exp. Clin. Cancer Res. 2019, 38, 54.
- Tian, F.-Q.; Chen, Z.-R.; Zhu, W.; Tang, M.-Q.; Li, J.-H.; Zhang, X.-C.; Jiang, J.; Cheng, X.-H. Inhibition of hsa_circ_0003489 shifts balance from autophagy to apoptosis and sensitizes multiple myeloma cells to bortezomib via miR-874-3p/HDAC1 axis. J. Gene Med. 2021, 23, e3329.
- Chen, F.; Wang, X.; Fu, S.; Wang, S.; Fu, Y.; Zhang, J.; Liu, Z. circular RNA circ-CDYL sponges miR-1180 to elevate yes-associated protein in multiple myeloma. Exp. Biol. Med. 2020, 245, 925–932.
- Li, Y.; Zhang, B.; Li, W.; Wang, L.; Yan, Z.; Li, H.; Yao, Y.; Yao, R.; Xu, K.; Li, Z. miR-15a/16 regulates the growth of myeloma cells, angiogenesis and antitumor immunity by inhibiting Bcl-2, VEGF-A and IL-17 expression in multiple myeloma. Leuk. Res. 2016, 49, 73–79.
- Kwon, J.J.; Factora, T.D.; Dey, S.; Kota, J. A systematic review of miR-29 in cancer. Mol. Ther.-Oncolytics 2019, 12, 173–194.
- Gu, Y.; Xiao, X.; Yang, S. LncRNA MALAT1 acts as an oncogene in multiple myeloma through sponging miR-509-5p to modulate FOXP1 expression. Oncotarget 2017, 8, 101984–101993.
- Liu, N.; Feng, S.; Li, H.; Chen, X.; Bai, S.; Liu, Y. Long non-coding RNA MALAT1 facilitates the tumorigenesis, invasion and glycolysis of multiple myeloma via miR-1271-5p/SOX13 axis. J. Cancer Res. Clin. Oncol. 2020, 146, 367–379.
- Yang, Y.; Chen, L. Downregulation of lncRNA UCA1 facilitates apoptosis and reduces proliferation in multiple myeloma via regulation of the miR-1271-5p/HGF axis. J. Chin. Med. Assoc. 2019, 82, 699–709.
- Raimondi, L.; Amodio, N.; Di Martino, M.T.; Altomare, E.; Leotta, M.; Caracciolo, D.; Gullà, A.; Neri, A.; Taverna, S.; D’Aquila, P.; et al. Targeting of multiple myeloma-related angiogenesis by miR-199a-5p mimics: In Vitro and In Vivo anti-tumor activity. Oncotarget 2014, 5, 3039–3054.
- Gowda, P.S.; Wildman, B.J.; Trotter, T.N.; Xu, X.; Hao, X.; Hassan, M.Q.; Yang, Y. Runx2 suppression by miR-342 and miR-363 inhibits multiple myeloma progression. Mol. Cancer Res. 2018, 16, 1138–1148.
- Inomata, M.; Tagawa, H.; Guo, Y.-M.; Kameoka, Y.; Takahashi, N.; Sawada, K. microRNA-17-92 down-regulates expression of distinct targets in different B-cell lymphoma subtypes. Blood 2009, 113, 396–402.
- Qi, X.; Zhang, D.-H.; Wu, N.; Xiao, J.-H.; Wang, X.; Ma, W. ceRNA in cancer: Possible functions and clinical implications. J. Med. Genet. 2015, 52, 710–718.
- Li, Z.; Kumar, S.; Jin, D.-Y.; Calin, G.A.; Chng, W.-J.; Siu, K.-L.; Poon, M.W.; Chim, C.S. Epigenetic silencing of long non-coding RNA BM742401 in multiple myeloma: Impact on prognosis and myeloma dissemination. Cancer Cell Int. 2020, 20, 403.
- Meng, Y.-B.; He, X.; Huang, Y.-F.; Wu, Q.-N.; Zhou, Y.-C.; Hao, D.-J. Long noncoding RNA CRNDE promotes multiple myeloma cell growth by suppressing miR-451. Oncol. Res. 2017, 25, 1207–1214.
- Tong, J.; Xu, X.; Zhang, Z.; Ma, C.; Xiang, R.; Liu, J.; Xu, W.; Wu, C.; Li, J.; Zhan, F.; et al. Hypoxia-induced long non-coding RNA DARS-AS1 regulates RBM39 stability to promote myeloma malignancy. Haematologica 2020, 105, 1630–1640.
- Pan, Y.; Zhang, Y.; Liu, W.; Huang, Y.; Shen, X.; Jing, R.; Pu, J.; Wang, X.; Ju, S.; Cong, H.; et al. LncRNA H19 overexpression induces bortezomib resistance in multiple myeloma by targeting MCL-1 via miR-29b-3p. Cell Death Dis. 2019, 10, 106.
- Guan, R.; Wang, W.; Fu, B.; Pang, Y.; Lou, Y.; Li, H. Increased lncRNA HOTAIR expression promotes the chemoresistance of multiple myeloma to dexamethasone by regulating cell viability and apoptosis by mediating the JAK2/STAT3 signaling pathway. Mol. Med. Rep. 2019, 20, 3917–3923.
- Hu, Y.; Lin, J.; Fang, H.; Fang, J.; Li, C.; Chen, W.; Liu, S.; Ondrejka, S.; Gong, Z.; Reu, F.; et al. Targeting the MALAT1/PARP1/LIG3 complex induces DNA damage and apoptosis in multiple myeloma. Leukemia 2018, 32, 2250–2262.
- Handa, H.; Kuroda, Y.; Kimura, K.; Masuda, Y.; Hattori, H.; Alkebsi, L.; Matsumoto, M.; Kasamatsu, T.; Kobayashi, N.; Tahara, K.-I.; et al. Long non-coding RNA MALAT1 is an inducible stress response gene associated with extramedullary spread and poor prognosis of multiple myeloma. Br. J. Haematol. 2017, 179, 449–460.
- Benetatos, L.; Dasoula, A.; Hatzimichael, E.; Georgiou, I.; Syrrou, M.; Bourantas, K.L. Promoter hypermethylation of the MEG3 (DLK1/MEG3) imprinted gene in multiple myeloma. Clin. Lymphoma Myeloma 2008, 8, 171–175.
- Fu, Y.; Liu, X.; Zhang, F.; Jiang, S.; Liu, J.; Luo, Y. Bortezomib-inducible long non-coding RNA myocardial infarction associated transcript is an oncogene in multiple myeloma that suppresses miR-29b. Cell Death Dis. 2019, 10, 319.
- Taiana, E.; Favasuli, V.; Ronchetti, D.; Todoerti, K.; Pelizzoni, F.; Manzoni, M.; Barbieri, M.; Fabris, S.; Silvestris, I.; Cantafio, M.E.G.; et al. Long non-coding RNA NEAT1 targeting impairs the DNA repair machinery and triggers anti-tumor activity in multiple myeloma. Leukemia 2020, 34, 234–244.
- Geng, W.; Guo, X.; Zhang, L.; Ma, Y.; Wang, L.; Liu, Z.; Ji, H.; Xiong, Y. Resveratrol inhibits proliferation, migration and invasion of multiple myeloma cells via NEAT1-mediated Wnt/beta-catenin signaling pathway. Biomed. Pharmacother. 2018, 107, 484–494.
- Dong, H.; Jiang, S.; Fu, Y.; Luo, Y.; Gui, R.; Liu, J. Upregulation of lncRNA NR_046683 serves as a prognostic biomarker and potential drug target for multiple myeloma. Front. Pharmacol. 2019, 10, 45.
- Yang, X.; Ye, H.; He, M.; Zhou, X.; Sun, N.; Guo, W.; Lin, X.; Huang, H.; Lin, Y.; Yao, R.; et al. LncRNA PDIA3P interacts with c-Myc to regulate cell proliferation via induction of pentose phosphate pathway in multiple myeloma. Biochem. Biophys. Res. Commun. 2018, 498, 207–213.
- Li, B.; Xu, H.; Han, H.; Song, S.; Zhang, X.; Ouyang, L.; Qian, C.; Hong, Y.; Qiu, Y.; Zhou, W.; et al. Exosome-mediated transfer of lncRUNX2-AS1 from multiple myeloma cells to MSCs contributes to osteogenesis. Oncogene 2018, 37, 5508–5519.
- Carrasco-Leon, A.; Ezponda, T.; Meydan, C.; Valcarcel, L.V.; Ordoñez, R.; Kulis, M.; Garate, L.; Miranda, E.; Segura, V.; Guruceaga, E.; et al. Characterization of complete lncRNAs transcriptome reveals the functional and clinical impact of lncRNAs in multiple myeloma. Leukemia 2021, 35, 1438–1450.
- Yang, X.; Huang, H.; Wang, X.; Liu, H.; Liu, H.; Lin, Z. Knockdown of lncRNA SNHG16 suppresses multiple myeloma cell proliferation by sponging miR-342-3p. Cancer Cell Int. 2020, 20, 38.
- Pu, J.; Huang, H.; Su, J.; Yuan, J.; Cong, H.; Wang, X.; Ju, S. Decreased expression of long noncoding RNA XLOC_013703 promotes cell growth via NF-kappaB pathway in multiple myeloma. IUBMB Life 2019, 71, 1240–1251.
- Li, B.; Hong, J.; Hong, M.; Wang, Y.; Yu, T.; Zang, S.; Wu, Q. piRNA-823 delivered by multiple myeloma-derived extracellular vesicles promoted tumorigenesis through re-educating endothelial cells in the tumor environment. Oncogene 2019, 38, 5227–5238.
- Yan, H.; Wu, Q.-L.; Sun, C.-Y.; Ai, L.-S.; Deng, J.; Zhang, L.; Chen, L.; Chu, Z.-B.; Tang, B.; Wang, K.; et al. piRNA-823 contributes to tumorigenesis by regulating de novo DNA methylation and angiogenesis in multiple myeloma. Leukemia 2015, 29, 196–206.
- Taulli, R.; Pandolfi, P.P. “Snorkeling” for missing players in cancer. J. Clin. Investig. 2012, 122, 2765–2768.
- Chu, L.; Su, M.Y.; Maggi, L.B., Jr.; Lu, L.; Mullins, C.; Crosby, S.; Huang, G.; Chng, W.J.; Vij, R.; Tomasson, M.H. Multiple myeloma-associated chromosomal translocation activates orphan snoRNA ACA11 to suppress oxidative stress. J. Clin. Investig. 2012, 122, 2793–2806.
- Oliveira, V.; Mahajan, N.; Bates, M.L.; Tripathi, C.; Kim, K.Q.; Zaher, H.S.; Maggi, L.B., Jr.; Tomasson, M.H. The snoRNA target of t(4;14) in multiple myeloma regulates ribosome biogenesis. FASEB BioAdvances 2019, 1, 404–414.
- Zhou, Y.; Goodenbour, J.M.; Godley, L.A.; Wickrema, A.; Pan, T. High levels of tRNA abundance and alteration of tRNA charging by bortezomib in multiple myeloma. Biochem. Biophys. Res. Commun. 2009, 385, 160–164.
- Zhou, F.; Wang, D.; Wei, W.; Chen, H.; Shi, H.; Zhou, N.; Wu, L.; Peng, R. Comprehensive profiling of circular RNA expressions reveals potential diagnostic and prognostic biomarkers in multiple myeloma. BMC Cancer 2020, 20, 40.
- Gao, M.; Li, C.; Xiao, H.; Dong, H.; Jiang, S.; Fu, Y.; Gong, L. hsa_circ_0007841: A novel potential biomarker and drug resistance for multiple myeloma. Front. Oncol. 2019, 9, 1261.

