Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Elisa Dultz | -- | 2183 | 2022-05-27 07:49:08 | | | |
| 2 | Nora Tang | -1 word(s) | 2182 | 2022-06-17 05:48:36 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Dultz, E.; , .; Medalia, O.; Onishchenko, E. Tour of the Nuclear Pore Complex Architecture. Encyclopedia. Available online: https://encyclopedia.pub/entry/23469 (accessed on 27 June 2026).
Dultz E, , Medalia O, Onishchenko E. Tour of the Nuclear Pore Complex Architecture. Encyclopedia. Available at: https://encyclopedia.pub/entry/23469. Accessed June 27, 2026.
Dultz, Elisa, , Ohad Medalia, Evgeny Onishchenko. "Tour of the Nuclear Pore Complex Architecture" Encyclopedia, https://encyclopedia.pub/entry/23469 (accessed June 27, 2026).
Dultz, E., , ., Medalia, O., & Onishchenko, E. (2022, May 27). Tour of the Nuclear Pore Complex Architecture. In Encyclopedia. https://encyclopedia.pub/entry/23469
Dultz, Elisa, et al. "Tour of the Nuclear Pore Complex Architecture." Encyclopedia. Web. 27 May, 2022.
Copy Citation
Nuclear pore complexes (NPCs) are the only transport channels that cross the nuclear envelope. Constructed from ~500–1000 nucleoporin proteins each, they are among the largest macromolecular assemblies in eukaryotic cells. Thanks to advances in structural analysis approaches, the construction principles and architecture of the NPC have recently been revealed at submolecular resolution. Although the overall structure and inventory of nucleoporins are conserved, NPCs exhibit significant compositional and functional plasticity even within single cells and surprising variability in their assembly pathways. Once assembled, NPCs remain seemingly unexchangeable in post-mitotic cells.
nuclear pore complex
nucleoporin
NPC
1. Inner Ring: The Flexible Core of the Nuclear Pore Complex
The architecture of the inner ring, with its symmetry along the nucleocytoplasmic axis, is highly conserved [1]. It coats the NE with eight spokes positioned around the central transport channel, each formed by three layers. Closest to the central channel, the innermost layer consists of the so-called channel nucleoporin heterotrimer (Nup49, Nup57, Nsp1), which projects intrinsically disordered FG-rich segments into the central NPC channel [2]. The outer, membrane-binding layer is composed of the α-solenoid/β-propeller domain paralogues Nup157/Nup170, which bind the NE via an amphipathic lipid packing sensor (ALPS) motif positioned in a loop between two β-propeller blades [3][4]. The paralogues Nup188/Nup192 have an NTR-like structure [5][6][7] and form the central layer between the membrane binding NUPs and the channel nucleoporin heterotrimer. The rigid layers are linked by flexible connectors: the flexible N-terminus of Nic96 ties the membrane binding layer to Nup188/Nup192 and the channel nucleoporin heterotrimer, and the short linear motifs (SliMs) in the membrane-interacting Nup53/Nup59 connect most of the inner ring NUPs [8][9][10][11][12][13]. While the NUPs within a single inner ring spoke have large interaction surfaces [9], recent structural models of the NPC propose that the interactions between spokes are minimal and instead are mostly mediated by natively disordered and flexible connector NUPs [8][9][10][11]. This flexibility allows the NPC to adjust its diameter depending on the physiological state of the cell [8][14][15][16][17][18], and the resulting spaces between spokes might solve the long-standing question of how transmembrane proteins can pass through the NPC. Intriguingly, Nup188/Nup192 (hsNup188/hsNup205; hs for Homo sapiens) not only share structural similarity with NTRs, their interaction with Nic96 (hsNup93) also resembles the interaction of the transport receptor importin-β and the importin-β binding domain (IBB) of its cargo complex [8][10][14][19]. This points to a common evolutionary origin of NUPs and transport receptors [5][6][7][8][20].
2. Symmetric Outer Rings: The Versatile Outer Coat of the Nuclear Pore Complex
The outer rings on the nuclear and cytoplasmic faces of the NPC are largely identical and made up of rigid subcomplexes known as Y complexes [21][22]. These building blocks are themselves composed of six conserved constituent proteins (Seh1 is not a conserved element of the Y complex in thermophilic fungi and Aspergillus nidulans [23][24][25]), which form a structure resembling the shape of the letter Y [26][27][28][29], and eight Y complexes assembled in a head-to-tail manner. S. cerevisiae has a single cytoplasmic and nucleoplasmic Y complex ring [8][30][31], human and Xenopus laevis NPCs carry two Y complex rings per side [4][19][21][32][33][34], and the green algae Chlamydomonas reinhardtii and fission yeast Schizosaccharomyces pombe exhibit an asymmetric distribution, with two nuclear and only one cytoplasmic Y complex rings [16][35]. Notably, the cytoplasmic Y complex ring of S. pombe only consists of the Y triskelion, breaking the head-to-tail arrangement observed in other species [16]. Surprisingly, the number of Y complex rings can vary even within the same cell: a subset of NPCs with two nucleoplasmic Y complex rings was recently observed in budding yeast ([8], further discussed below).
The largely α-solenoid core of the Y complex is tethered to the NE by ALPS motifs in the β-propeller of Nup120 and Nup133 [36][37][38] and decorated by several species-specific β-propeller NUPs [39][40][41]. The α-solenoid/β-propeller architecture of the outer and inner ring NUPs is similar to the vesicle-coating protein complexes COPI and COPII, and the β-propeller protein Sec13 is a shared component of both NPCs and COPII complexes, suggesting a common evolutionary origin (reviewed in [42]). Although the eightfold rotational symmetry of the NPC is well established, deviations have been observed in X. laevis NPCs [43][44], which raises the question how the eightfold symmetry of the NPC is formed. Since the connections between inner ring spokes are flexible and the inner ring diameter can change drastically [14][15][16][17], it seems likely that the oligomerization of the Y complex ring plays a key role in establishing the correct stoichiometry of NPC subunits. However, the Y complex itself is not a rigid structure and has multiple hinge points [26], and additional constraints, such as, e.g., membrane interaction may thus be needed to determine the eightfold symmetry of the NPC.
The outer rings are connected to the inner ring by a set of paralogues with flexible linkers (Nup116, Nup100, Nup145N) [10][12][31][45], and the double rings in metazoan and C. reinhardtii NPCs are linked by an additional copy of hsNup155 or crNup155 (cr: C. reinhardtii), respectively [4][21][32][35]. In metazoan NPCs, the chromatin-binding NUP ELYS is associated with one short arm of the Y complex on the nucleoplasmic side [14][19][46][47][48].
Interestingly, recent biochemical characterization and higher-resolution electron microscopy (EM) maps of the NPC revealed that the importin-β-IBB-like complex hsNup205-hsNup93 is not only a part of the inner ring, but it can also be found in the outer rings of metazoan NPCs [11][14][19][34]. A characteristic question mark-shaped density can also be seen in EM maps from the double Y complex rings of S. cerevisiae and C. reinhardtii [8][35], and the presence of the hsNup205-hsNup93 heterodimer and its homologues may thus be conserved and important for the oligomerization of double Y rings.
3. Asymmetric Appendages: Functional Extensions of the Nuclear Pore Complex
The symmetry of the outer rings is broken by several subcomplexes that specifically bind to the cytoplasmic or nucleoplasmic ring. Identified by classical EM experiments, the cytoplasmic filaments and nuclear basket are the most prominent asymmetric components of the NPC [49][50][51]. The term cytoplasmic filaments is often used as a synonym for all NUPs that preferentially localize to the cytoplasmic side of the NPC. However, the main component of these elongated filaments protruding into the cytoplasm in metazoa is the largely disordered C-terminus of hsNup358, which harbors multiple Zinc-fingers and Ran-binding domains, and plays an important role in receptor-mediated transport and protein translation [11]. hsNup358 is specific to metazoa and stabilizes the cytoplasmic double Y ring [11][32].
The majority of the other cytoplasmic NUPs are conserved across species and form the so-called mRNA export platform. This extends to the center of the NPC [8][30][52][53][54] and plays a key role in mRNA export and remodeling [55]. Intriguingly, the mRNA export platform has high similarity with the channel nucleoporin heterotrimer at the center of the NPC, with Nsp1 being a shared component between the two. Further, the positioning of the hsNup93-hsNup205 heterodimer in the cytoplasmic outer ring and its biochemical interactions suggest that hsNup93 connects the cytoplasmic mRNA export platform in a similar way as the channel nucleoporin heterotrimer in the inner ring [11]. Interestingly, the mRNA export platforms in metazoa and yeast have different overall architectures. In yeast, the cytoplasmic coiled-coil NUPs form a single complex, whereas two parallel-orientated complexes are present in the X. laevis NPC [19]. This corresponds to the number of cytoplasmic Y rings in the two species. Intriguingly, the mRNA export platform is entirely absent in the more divergent eukaryote Trypanosoma brucei [1][56]. In contrast to the conserved Y complex and inner ring, the mRNA export platform might thus have specialized to meet the needs of the respective organism during evolution.
The nuclear basket was identified in early EM studies because of its characteristic elongated structure [49][50], but due to its flexible nature, it remains one of the least structurally characterized modules. The majority of the basket-like structure seen by classical EM analysis [57][58] likely stems from the large coiled-coil hsTPR (S. cerevisiae Mlp1/Mlp2) [59][60]. Although the stoichiometry of the nuclear basket coiled-coil NUPs is not entirely clear [31][61][62][63][64], up to eight basket-like filaments protruding into the nucleoplasm and tethering proteasomes to the NPC have been observed at single NPCs of C. reinhardtii [65].
So far, the best-resolved fragments of the nuclear basket are coiled-coil segments that likely belong to Mlp1/2 and bind to the nuclear Y complex [8], which is consistent with other EM and crosslinking data [30][31]. The inventory of the S. cerevisiae nuclear basket is completed by the mostly disordered Nup1, Nup2, and Nup60. Although these NUPs have evaded structural characterization, biochemical studies show that Nup1 and Nup60 (hsNup153) interact with the NE via an amphipathic helix (AH) [66][67][68]. Similar to the linker NUPs in other subcomplexes, Nup60 flexibly connects the nucleoplasmic Y complex ring with the Mlps and Nup2 (hsNup50) via SLiMs [66][69][70]. Further, Nup1, Nup2, and Nup60 contain FG repeats and, together with the Mlps, are important for export and quality control of mRNA (reviewed in [71]).
4. The Membrane Ring: An Enigmatic Girdle
Besides the membrane interactions of the inner and outer rings mediated by ALPS motifs, the NPC is also directly anchored in the NE by transmembrane NUPs. Because of their transmembrane regions, it is difficult to purify these proteins or distinguish them from the NE in EM studies, and the structure of the membrane ring is thus poorly characterized. In S. cerevisiae, there are four transmembrane NUPs, which are not as highly conserved as other components of the NPC [72] (reviewed in [73]). Only Ndc1 has a well-defined ortholog in metazoa [74][75], and is the only essential protein of this group. Ndc1 interacts with the inner ring NUPs Nup53/59 and Nup170 (in humans: hsNdc1, hsNup35, hsNup155, and additionally ALADIN) to form a membrane interaction hub that anchors the inner ring to the NE [8][13][14]. Pom152 and the human Gp210 are the only NUPs with structured domains in the NE lumen: both contain a series of luminal immunoglobulin repeats [76][77][78]. Despite the low primary sequence conservation and different membrane topology, the high structural similarity could hint at a common origin for both proteins. The immunoglobulin folds of Pom152 form a belt-like chain of beads around the NPC in the NE-lumen, which is anchored near the membrane interaction hub [8][16][31][77][78][79]. The belt-like luminal ring deforms together with changes in NPC diameter [8][16], which raises the possibility that it regulates the diameter of the NPC. However, neither Pom152 nor Gp210 is essential [80][81], and deletion of Gp210 does not lead to variation of the NPC diameter in cellulo [14]. Furthermore, the expression level of Gp210 in different cell lines varies widely [63][82], suggesting a more intricate role of the luminal ring than as just a mechanical girdle.
5. Linker Nucleoporins: An Invisible Thread Stitching the Nuclear Pore Complex Together
The NPC embodies two seemingly contradictory properties. On the one hand, it uses rigid building blocks with large interaction surfaces to form stable subcomplexes, such as the Y complex and the inner ring spokes, which confer a high degree of stability to the NPC core in post-mitotic cells [83][84][85][86][87] (reviewed in [88]). On the other hand, its structural flexibility allows for drastic changes in diameter [8][15][16][17] and likely enables a fast assembly and disassembly of the NPC in open mitosis [89]. How can these properties coexist in one structure? The emerging solution is a peculiar mode of association between the different NPC modules via intrinsically disordered NUPs. Homologues of the S. cerevisiae FG repeat NUPs Nup100, Nup145N and Nup116, and non-FG NUPs Nup53 and Nup59 are universally capable of linking several NPC elements each via SLiMs spread throughout their intrinsically disordered domains [2][12][90][91]. In this way, each of them can flexibly join several core subunits, akin to beads on a string. The electron densities observed next to the core NUPs in high-resolution NPC maps and chemical crosslinking data all point to SLiM-mediated connectivity of the NPC subunits [8][10][11][14][31]. Further, flexible connections could arise from the ability of some core NUPs to directly bind FG repeats [45]. These multivalent interactions might create a velcro-like effect that brings about both stability and structural plasticity (Figure 1).
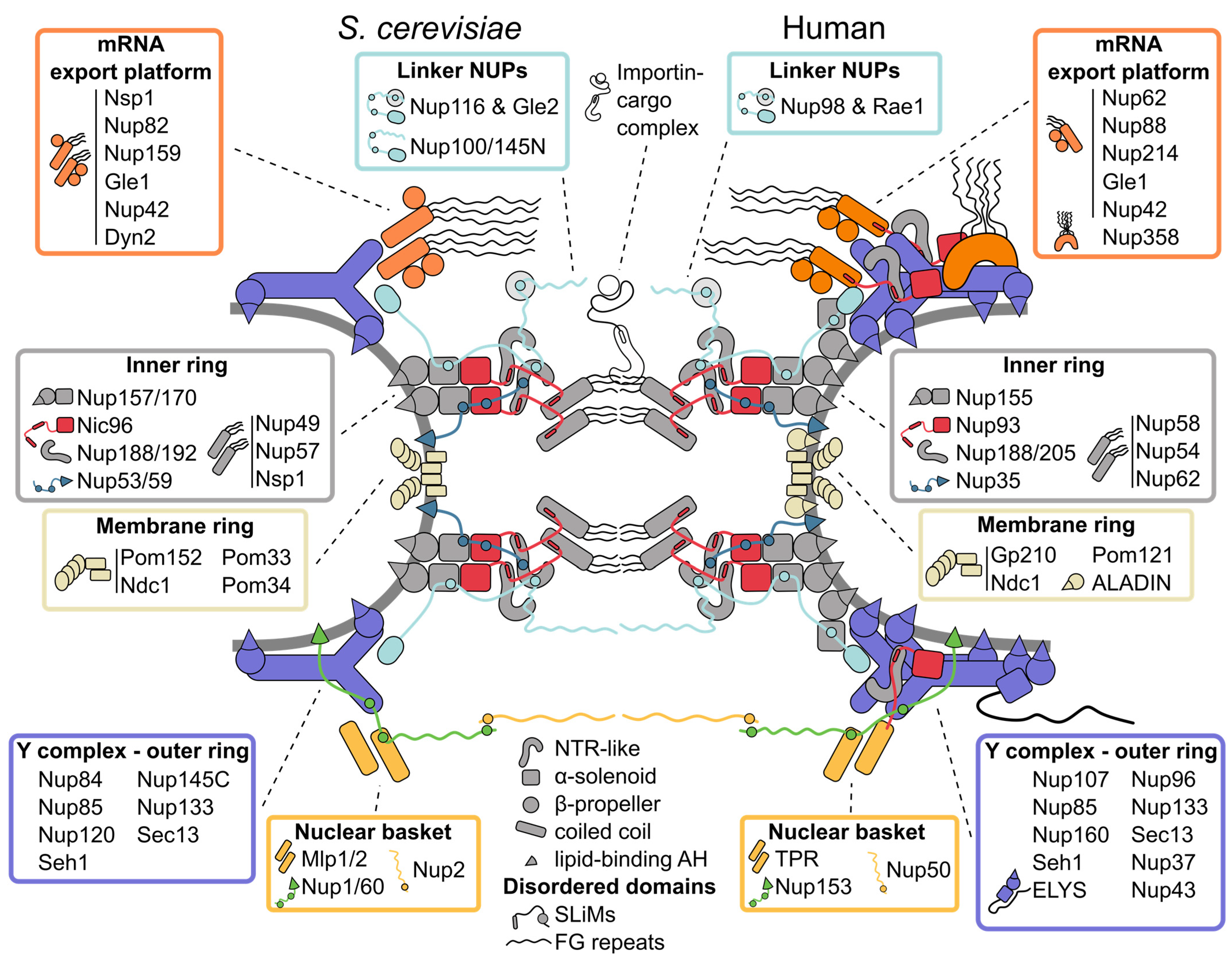
Figure 1. Inventory of the budding yeast and human nuclear pore complex. The nuclear pore complex (NPC) forms a channel connecting the nucleoplasm (bottom) with the cytoplasm (top) and is built of three concentric rings: the cytoplasmic outer ring, the inner ring, and the nucleoplasmic outer ring. The basic building blocks of the NPC are nucleoporins (NUPs), which are organized into several subcomplexes (boxes) and largely composed of only a few structural motifs (center bottom). The rigid subcomplexes are connected by disordered linkers. They contain short linear interaction motifs (SLiMs), which flexibly tie the NUPs and subcomplexes together. Multiple NUPs contain a lipid-binding amphipathic helix (AH) that helps tether the NPC to the lipid membrane. See text for details.
The interactions via short motifs are a prevailing theme also outside the NPC core. Short motif interactions contribute to the attachment of the mRNA export platform, central channel NUPs, the nuclear basket and transmembrane NUPs [11][12][13][66][67] (Figure 1). Interestingly, the interactions of the NPC with the NE rely on the same principle. Although transmembrane NUPs are one of the least evolutionarily conserved groups [72] (reviewed in [73]), short lipid-binding AHs found within multiple core and non-core NUPs are a conserved feature, often seen positioned along the lipid membrane in current NPC models [8][14]. This multitude of binding sites could stabilize the high curvature induced in the lipid membrane and establish a tight association of the NPC spokes to the pore membrane (reviewed in [92]).
Taken together, the short interaction motifs and intrinsically disordered domains emerge as key elements of NPC connectivity.
References
- DeGrasse, J.A.; DuBois, K.N.; Devos, D.; Siegel, T.N.; Sali, A.; Field, M.C.; Rout, M.P.; Chait, B.T. Evidence for a shared nuclear pore complex architecture that is conserved from the last common eukaryotic ancestor. Mol. Cell Proteom. 2009, 8, 2119–2130.
- Stuwe, T.; Bley, C.J.; Thierbach, K.; Petrovic, S.; Schilbach, S.; Mayo, D.J.; Perriches, T.; Rundlet, E.J.; Jeon, Y.E.; Collins, L.N.; et al. Architecture of the fungal nuclear pore inner ring complex. Science 2015, 350, 56–64.
- Whittle, J.R.R.; Schwartz, T.U. Architectural nucleoporins Nup157/170 and Nup133 are structurally related and descend from a second ancestral element. J. Biol. Chem. 2009, 284, 28442–28452.
- Lin, D.H.; Stuwe, T.; Schilbach, S.; Rundlet, E.J.; Perriches, T.; Mobbs, G.; Fan, Y.; Thierbach, K.; Huber, F.M.; Collins, L.N.; et al. Architecture of the symmetric core of the nuclear pore. Science 2016, 352, aaf1015.
- Andersen, K.R.; Onischenko, E.; Tang, J.H.; Kumar, P.; Chen, J.Z.; Ulrich, A.; Liphardt, J.T.; Weis, K.; Schwartz, T.U. Scaffold nucleoporins Nup188 and Nup192 share structural and functional properties with nuclear transport receptors. eLife 2013, 2, e00745.
- Flemming, D.; Devos, D.P.; Schwarz, J.; Amlacher, S.; Lutzmann, M.; Hurt, E. Analysis of the yeast nucleoporin Nup188 reveals a conserved S-like structure with similarity to karyopherins. J. Struct. Biol. 2012, 177, 99–105.
- Stuwe, T.; Lin, D.H.; Collins, L.N.; Hurt, E.; Hoelz, A. Evidence for an evolutionary relationship between the large adaptor nucleoporin Nup192 and karyopherins. Proc. Natl. Acad. Sci. USA 2014, 111, 2530–2535.
- Akey, C.W.; Singh, D.; Ouch, C.; Echeverria, I.; Nudelman, I.; Varberg, J.M.; Yu, Z.; Fang, F.; Shi, Y.; Wang, J.; et al. Comprehensive structure and functional adaptations of the yeast nuclear pore complex. Cell 2022, 185, 361–378.e25.
- Li, Z.; Chen, S.; Zhao, L.; Huang, G.; Pi, X.; Sun, S.; Wang, P.; Sui, S.F. Near-atomic structure of the inner ring of the Saccharomyces cerevisiae nuclear pore complex. Cell Res. 2022, 1–14.
- Petrovic, S.; Samanta, D.; Perriches, T.; Bley, C.J.; Thierbach, K.; Brown, B.; Nie, S.; Mobbs, G.W.; Stevens, T.A.; Liu, X.; et al. Architecture of the linker-scaffold in the nuclear pore. bioRxiv 2021.
- Bley, C.J.; Nie, S.; Mobbs, G.W.; Petrovic, S.; Gres, A.T.; Liu, X.; Mukherjee, S.; Harvey, S.; Huber, F.M.; Lin, D.H.; et al. Architecture of the cytoplasmic face of the nuclear pore. bioRxiv 2021.
- Fischer, J.; Teimer, R.; Amlacher, S.; Kunze, R.; Hurt, E. Linker Nups connect the nuclear pore complex inner ring with the outer ring and transport channel. Nat. Struct. Mol. Biol. 2015, 22, 774–781.
- Onischenko, E.; Stanton, L.H.; Madrid, A.S.; Kieselbach, T.; Weis, K. Role of the Ndc1 interaction network in yeast nuclear pore complex assembly and maintenance. J. Cell Biol. 2009, 185, 475–491.
- Mosalaganti, S.; Obarska-Kosinska, A.; Siggel, M.; Turonova, B.; Zimmerli, C.E.; Buczak, K.; Schmidt, F.H.; Margiotta, E.; Mackmull, M.-T.; Hagen, W.; et al. Artificial intelligence reveals nuclear pore complexity. bioRxiv 2021.
- Schuller, A.P.; Wojtynek, M.; Mankus, D.; Tatli, M.; Kronenberg-Tenga, R.; Regmi, S.G.; Dip, P.V.; Lytton-Jean, A.K.R.; Brignole, E.J.; Dasso, M.; et al. The cellular environment shapes the nuclear pore complex architecture. Nature 2021, 598, 667–671.
- Zimmerli, C.E.; Allegretti, M.; Rantos, V.; Goetz, S.K.; Obarska-Kosinska, A.; Zagoriy, I.; Halavatyi, A.; Hummer, G.; Mahamid, J.; Kosinski, J.; et al. Nuclear pores dilate and constrict in cellulo. Science 2021, 374, eabd9776.
- Zila, V.; Margiotta, E.; Turonova, B.; Muller, T.G.; Zimmerli, C.E.; Mattei, S.; Allegretti, M.; Borner, K.; Rada, J.; Muller, B.; et al. Cone-shaped HIV-1 capsids are transported through intact nuclear pores. Cell 2021, 184, 1032–1046.e18.
- Liashkovich, I.; Meyring, A.; Kramer, A.; Shahin, V. Exceptional structural and mechanical flexibility of the nuclear pore complex. J. Cell Physiol. 2011, 226, 675–682.
- Tai, L.; Zhu, Y.; Ren, H.; Huang, X.; Zhang, C.; Sun, F. 8 Å structure of the outer rings of the Xenopus laevis nuclear pore complex obtained by cryo-EM and AI. Protein Cell 2022, 1–18.
- Sampathkumar, P.; Kim, S.J.; Upla, P.; Rice, W.J.; Phillips, J.; Timney, B.L.; Pieper, U.; Bonanno, J.B.; Fernandez-Martinez, J.; Hakhverdyan, Z.; et al. Structure, dynamics, evolution, and function of a major scaffold component in the nuclear pore complex. Structure 2013, 21, 560–571.
- Bui, K.H.; von Appen, A.; DiGuilio, A.L.; Ori, A.; Sparks, L.; Mackmull, M.T.; Bock, T.; Hagen, W.; Andres-Pons, A.; Glavy, J.S.; et al. Integrated structural analysis of the human nuclear pore complex scaffold. Cell 2013, 155, 1233–1243.
- Maimon, T.; Elad, N.; Dahan, I.; Medalia, O. The human nuclear pore complex as revealed by cryo-electron tomography. Structure 2012, 20, 998–1006.
- Kelley, K.; Knockenhauer, K.E.; Kabachinski, G.; Schwartz, T.U. Atomic structure of the Y complex of the nuclear pore. Nat. Struct. Mol. Biol. 2015, 22, 425–431.
- Thierbach, K.; von Appen, A.; Thoms, M.; Beck, M.; Flemming, D.; Hurt, E. Protein interfaces of the conserved Nup84 complex from Chaetomium thermophilum shown by crosslinking mass spectrometry and electron microscopy. Structure 2013, 21, 1672–1682.
- Liu, H.L.; De Souza, C.P.; Osmani, A.H.; Osmani, S.A. The three fungal transmembrane nuclear pore complex proteins of Aspergillus nidulans are dispensable in the presence of an intact An-Nup84-120 complex. Mol. Biol. Cell 2009, 20, 616–630.
- Kampmann, M.; Blobel, G. Three-dimensional structure and flexibility of a membrane-coating module of the nuclear pore complex. Nat. Struct. Mol. Biol. 2009, 16, 782–788.
- Lutzmann, M.; Kunze, R.; Buerer, A.; Aebi, U.; Hurt, E. Modular self-assembly of a Y-shaped multiprotein complex from seven nucleoporins. EMBO J. 2002, 21, 387–397.
- Nordeen, S.A.; Turman, D.L.; Schwartz, T.U. Yeast Nup84-Nup133 complex structure details flexibility and reveals conservation of the membrane anchoring ALPS motif. Nat. Commun. 2020, 11, 6060.
- Siniossoglou, S.; Lutzmann, M.; Santos-Rosa, H.; Leonard, K.; Mueller, S.; Aebi, U.; Hurt, E. Structure and assembly of the Nup84p complex. J. Cell Biol. 2000, 149, 41–54.
- Allegretti, M.; Zimmerli, C.E.; Rantos, V.; Wilfling, F.; Ronchi, P.; Fung, H.K.H.; Lee, C.W.; Hagen, W.; Turonova, B.; Karius, K.; et al. In-cell architecture of the nuclear pore and snapshots of its turnover. Nature 2020, 586, 796–800.
- Kim, S.J.; Fernandez-Martinez, J.; Nudelman, I.; Shi, Y.; Zhang, W.; Raveh, B.; Herricks, T.; Slaughter, B.D.; Hogan, J.A.; Upla, P.; et al. Integrative structure and functional anatomy of a nuclear pore complex. Nature 2018, 555, 475–482.
- von Appen, A.; Kosinski, J.; Sparks, L.; Ori, A.; DiGuilio, A.L.; Vollmer, B.; Mackmull, M.T.; Banterle, N.; Parca, L.; Kastritis, P.; et al. In situ structural analysis of the human nuclear pore complex. Nature 2015, 526, 140–143.
- Eibauer, M.; Pellanda, M.; Turgay, Y.; Dubrovsky, A.; Wild, A.; Medalia, O. Structure and gating of the nuclear pore complex. Nat. Commun. 2015, 6, 7532.
- Huang, G.; Zhang, Y.; Zhu, X.; Zeng, C.; Wang, Q.; Zhou, Q.; Tao, Q.; Liu, M.; Lei, J.; Yan, C.; et al. Structure of the cytoplasmic ring of the Xenopus laevis nuclear pore complex by cryo-electron microscopy single particle analysis. Cell Res. 2020, 30, 520–531.
- Mosalaganti, S.; Kosinski, J.; Albert, S.; Schaffer, M.; Strenkert, D.; Salome, P.A.; Merchant, S.S.; Plitzko, J.M.; Baumeister, W.; Engel, B.D.; et al. In situ architecture of the algal nuclear pore complex. Nat. Commun. 2018, 9, 2361.
- Berke, I.C.; Boehmer, T.; Blobel, G.; Schwartz, T.U. Structural and functional analysis of Nup133 domains reveals modular building blocks of the nuclear pore complex. J. Cell Biol. 2004, 167, 591–597.
- Drin, G.; Casella, J.F.; Gautier, R.; Boehmer, T.; Schwartz, T.U.; Antonny, B. A general amphipathic alpha-helical motif for sensing membrane curvature. Nat. Struct. Mol. Biol. 2007, 14, 138–146.
- Leksa, N.C.; Brohawn, S.G.; Schwartz, T.U. The structure of the scaffold nucleoporin Nup120 reveals a new and unexpected domain architecture. Structure 2009, 17, 1082–1091.
- Brohawn, S.G.; Leksa, N.C.; Spear, E.D.; Rajashankar, K.R.; Schwartz, T.U. Structural evidence for common ancestry of the nuclear pore complex and vesicle coats. Science 2008, 322, 1369–1373.
- Debler, E.W.; Ma, Y.; Seo, H.S.; Hsia, K.C.; Noriega, T.R.; Blobel, G.; Hoelz, A. A fence-like coat for the nuclear pore membrane. Mol. Cell 2008, 32, 815–826.
- Hsia, K.C.; Stavropoulos, P.; Blobel, G.; Hoelz, A. Architecture of a coat for the nuclear pore membrane. Cell 2007, 131, 1313–1326.
- Field, M.C.; Rout, M.P. Pore timing: The evolutionary origins of the nucleus and nuclear pore complex. F1000Res 2019, 8.
- Hinshaw, J.E.; Milligan, R.A. Nuclear pore complexes exceeding eightfold rotational symmetry. J. Struct. Biol. 2003, 141, 259–268.
- Loschberger, A.; Franke, C.; Krohne, G.; van de Linde, S.; Sauer, M. Correlative super-resolution fluorescence and electron microscopy of the nuclear pore complex with molecular resolution. J. Cell Sci. 2014, 127, 4351–4355.
- Onischenko, E.; Tang, J.H.; Andersen, K.R.; Knockenhauer, K.E.; Vallotton, P.; Derrer, C.P.; Kralt, A.; Mugler, C.F.; Chan, L.Y.; Schwartz, T.U.; et al. Natively Unfolded FG Repeats Stabilize the Structure of the Nuclear Pore Complex. Cell 2017, 171, 904–917.e19.
- Franz, C.; Walczak, R.; Yavuz, S.; Santarella, R.; Gentzel, M.; Askjaer, P.; Galy, V.; Hetzer, M.; Mattaj, I.W.; Antonin, W. MEL-28/ELYS is required for the recruitment of nucleoporins to chromatin and postmitotic nuclear pore complex assembly. EMBO Rep. 2007, 8, 165–172.
- Rasala, B.A.; Orjalo, A.V.; Shen, Z.; Briggs, S.; Forbes, D.J. ELYS is a dual nucleoporin/kinetochore protein required for nuclear pore assembly and proper cell division. Proc. Natl. Acad. Sci. USA 2006, 103, 17801–17806.
- Rasala, B.A.; Ramos, C.; Harel, A.; Forbes, D.J. Capture of AT-rich chromatin by ELYS recruits POM121 and NDC1 to initiate nuclear pore assembly. Mol. Biol. Cell 2008, 19, 3982–3996.
- Jarnik, M.; Aebi, U. Toward a more complete 3-D structure of the nuclear pore complex. J. Struct. Biol. 1991, 107, 291–308.
- Goldberg, M.W.; Allen, T.D. The nuclear pore complex: Three-dimensional surface structure revealed by field emission, in-lens scanning electron microscopy, with underlying structure uncovered by proteolysis. J. Cell Sci. 1993, 106 Pt 1, 261–274.
- Watson, M.L. Further observations on the nuclear envelope of the animal cell. J. Biophys Biochem. Cytol. 1959, 6, 147–156.
- Fernandez-Martinez, J.; Kim, S.J.; Shi, Y.; Upla, P.; Pellarin, R.; Gagnon, M.; Chemmama, I.E.; Wang, J.; Nudelman, I.; Zhang, W.; et al. Structure and Function of the Nuclear Pore Complex Cytoplasmic mRNA Export Platform. Cell 2016, 167, 1215–1228.e25.
- Walther, T.C.; Pickersgill, H.S.; Cordes, V.C.; Goldberg, M.W.; Allen, T.D.; Mattaj, I.W.; Fornerod, M. The cytoplasmic filaments of the nuclear pore complex are dispensable for selective nuclear protein import. J. Cell Biol. 2002, 158, 63–77.
- Vallotton, P.; Rajoo, S.; Wojtynek, M.; Onischenko, E.; Kralt, A.; Derrer, C.P.; Weis, K. Mapping the native organization of the yeast nuclear pore complex using nuclear radial intensity measurements. Proc. Natl. Acad. Sci. USA 2019, 116, 14606–14613.
- Lin, D.H.; Hoelz, A. The Structure of the Nuclear Pore Complex (An Update). Annu. Rev. Biochem. 2019, 88, 725–783.
- Obado, S.O.; Brillantes, M.; Uryu, K.; Zhang, W.; Ketaren, N.E.; Chait, B.T.; Field, M.C.; Rout, M.P. Interactome Mapping Reveals the Evolutionary History of the Nuclear Pore Complex. PLoS Biol. 2016, 14, e1002365.
- Goldberg, M.W.; Allen, T.D. High resolution scanning electron microscopy of the nuclear envelope: Demonstration of a new, regular, fibrous lattice attached to the baskets of the nucleoplasmic face of the nuclear pores. J. Cell Biol. 1992, 119, 1429–1440.
- Kiseleva, E.; Allen, T.D.; Rutherford, S.; Bucci, M.; Wente, S.R.; Goldberg, M.W. Yeast nuclear pore complexes have a cytoplasmic ring and internal filaments. J. Struct. Biol. 2004, 145, 272–288.
- Krull, S.; Thyberg, J.; Bjorkroth, B.; Rackwitz, H.R.; Cordes, V.C. Nucleoporins as components of the nuclear pore complex core structure and Tpr as the architectural element of the nuclear basket. Mol. Biol. Cell 2004, 15, 4261–4277.
- Niepel, M.; Molloy, K.R.; Williams, R.; Farr, J.C.; Meinema, A.C.; Vecchietti, N.; Cristea, I.M.; Chait, B.T.; Rout, M.P.; Strambio-De-Castillia, C. The nuclear basket proteins Mlp1p and Mlp2p are part of a dynamic interactome including Esc1p and the proteasome. Mol. Biol. Cell 2013, 24, 3920–3938.
- Li, Y.; Aksenova, V.; Tingey, M.; Yu, J.; Ma, P.; Arnaoutov, A.; Chen, S.; Dasso, M.; Yang, W. Distinct roles of nuclear basket proteins in directing the passage of mRNA through the nuclear pore. Proc. Natl. Acad. Sci. USA 2021, 118.
- Mi, L.; Goryaynov, A.; Lindquist, A.; Rexach, M.; Yang, W. Quantifying nucleoporin stoichiometry inside single nuclear pore complexes in vivo. Sci. Rep. 2015, 5, 9372.
- Ori, A.; Banterle, N.; Iskar, M.; Andres-Pons, A.; Escher, C.; Khanh Bui, H.; Sparks, L.; Solis-Mezarino, V.; Rinner, O.; Bork, P.; et al. Cell type-specific nuclear pores: A case in point for context-dependent stoichiometry of molecular machines. Mol. Syst. Biol. 2013, 9, 648.
- Rajoo, S.; Vallotton, P.; Onischenko, E.; Weis, K. Stoichiometry and compositional plasticity of the yeast nuclear pore complex revealed by quantitative fluorescence microscopy. Proc. Natl. Acad. Sci. USA 2018, 115, E3969–E3977.
- Albert, S.; Schaffer, M.; Beck, F.; Mosalaganti, S.; Asano, S.; Thomas, H.F.; Plitzko, J.M.; Beck, M.; Baumeister, W.; Engel, B.D. Proteasomes tether to two distinct sites at the nuclear pore complex. Proc. Natl. Acad. Sci. USA 2017, 114, 13726–13731.
- Cibulka, J.; Bisaccia, F.; Radisavljevic, K.; Gudino Carrillo, R.M.; Kohler, A. Assembly principle of a membrane-anchored nuclear pore basket scaffold. Sci. Adv. 2022, 8, eabl6863.
- Mészáros, N.; Cibulka, J.; Mendiburo, M.J.; Romanauska, A.; Schneider, M.; Köhler, A. Nuclear pore basket proteins are tethered to the nuclear envelope and can regulate membrane curvature. Dev. Cell 2015, 33, 285–298.
- Vollmer, B.; Lorenz, M.; Moreno-Andres, D.; Bodenhofer, M.; De Magistris, P.; Astrinidis, S.A.; Schooley, A.; Flotenmeyer, M.; Leptihn, S.; Antonin, W. Nup153 Recruits the Nup107-160 Complex to the Inner Nuclear Membrane for Interphasic Nuclear Pore Complex Assembly. Dev. Cell 2015, 33, 717–728.
- Hase, M.E.; Cordes, V.C. Direct interaction with nup153 mediates binding of Tpr to the periphery of the nuclear pore complex. Mol. Biol. Cell 2003, 14, 1923–1940.
- Makise, M.; Mackay, D.R.; Elgort, S.; Shankaran, S.S.; Adam, S.A.; Ullman, K.S. The Nup153-Nup50 protein interface and its role in nuclear import. J. Biol. Chem. 2012, 287, 38515–38522.
- Bensidoun, P.; Zenklusen, D.; Oeffinger, M. Choosing the right exit: How functional plasticity of the nuclear pore drives selective and efficient mRNA export. Wiley Interdiscip. Rev. RNA 2021, 12, e1660.
- Neumann, N.; Lundin, D.; Poole, A.M. Comparative genomic evidence for a complete nuclear pore complex in the last eukaryotic common ancestor. PLoS ONE 2010, 5, e13241.
- Field, M.C.; Koreny, L.; Rout, M.P. Enriching the pore: Splendid complexity from humble origins. Traffic 2014, 15, 141–156.
- Chadrin, A.; Hess, B.; San Roman, M.; Gatti, X.; Lombard, B.; Loew, D.; Barral, Y.; Palancade, B.; Doye, V. Pom33, a novel transmembrane nucleoporin required for proper nuclear pore complex distribution. J. Cell Biol. 2010, 189, 795–811.
- Stavru, F.; Hulsmann, B.B.; Spang, A.; Hartmann, E.; Cordes, V.C.; Gorlich, D. NDC1: A crucial membrane-integral nucleoporin of metazoan nuclear pore complexes. J. Cell Biol. 2006, 173, 509–519.
- Tcheperegine, S.E.; Marelli, M.; Wozniak, R.W. Topology and functional domains of the yeast pore membrane protein Pom152p. J. Biol. Chem. 1999, 274, 5252–5258.
- Upla, P.; Kim, S.J.; Sampathkumar, P.; Dutta, K.; Cahill, S.M.; Chemmama, I.E.; Williams, R.; Bonanno, J.B.; Rice, W.J.; Stokes, D.L.; et al. Molecular Architecture of the Major Membrane Ring Component of the Nuclear Pore Complex. Structure 2017, 25, 434–445.
- Hao, Q.; Zhang, B.; Yuan, K.; Shi, H.; Blobel, G. Electron microscopy of Chaetomium pom152 shows the assembly of ten-bead string. Cell Discov. 2018, 4, 56.
- Zhang, Y.; Li, S.; Zeng, C.; Huang, G.; Zhu, X.; Wang, Q.; Wang, K.; Zhou, Q.; Yan, C.; Zhang, W.; et al. Molecular architecture of the luminal ring of the Xenopus laevis nuclear pore complex. Cell Res. 2020, 30, 532–540.
- Stavru, F.; Nautrup-Pedersen, G.; Cordes, V.C.; Gorlich, D. Nuclear pore complex assembly and maintenance in POM121- and gp210-deficient cells. J. Cell Biol. 2006, 173, 477–483.
- Wozniak, R.W.; Blobel, G.; Rout, M.P. POM152 is an integral protein of the pore membrane domain of the yeast nuclear envelope. J. Cell Biol. 1994, 125, 31–42.
- Olsson, M.; Scheele, S.; Ekblom, P. Limited expression of nuclear pore membrane glycoprotein 210 in cell lines and tissues suggests cell-type specific nuclear pores in metazoans. Exp. Cell Res. 2004, 292, 359–370.
- D’Angelo, M.A.; Raices, M.; Panowski, S.H.; Hetzer, M.W. Age-dependent deterioration of nuclear pore complexes causes a loss of nuclear integrity in postmitotic cells. Cell 2009, 136, 284–295.
- Rabut, G.; Doye, V.; Ellenberg, J. Mapping the dynamic organization of the nuclear pore complex inside single living cells. Nat. Cell Biol. 2004, 6, 1114–1121.
- Savas, J.N.; Toyama, B.H.; Xu, T.; Yates, J.R., 3rd; Hetzer, M.W. Extremely long-lived nuclear pore proteins in the rat brain. Science 2012, 335, 942.
- Toyama, B.H.; Arrojo, E.D.R.; Lev-Ram, V.; Ramachandra, R.; Deerinck, T.J.; Lechene, C.; Ellisman, M.H.; Hetzer, M.W. Visualization of long-lived proteins reveals age mosaicism within nuclei of postmitotic cells. J. Cell Biol. 2019, 218, 433–444.
- Toyama, B.H.; Savas, J.N.; Park, S.K.; Harris, M.S.; Ingolia, N.T.; Yates, J.R., 3rd; Hetzer, M.W. Identification of long-lived proteins reveals exceptional stability of essential cellular structures. Cell 2013, 154, 971–982.
- Knockenhauer, K.E.; Schwartz, T.U. The nuclear pore complex as a flexible and dynamic gate. Cell 2016, 164, 1162–1171.
- Dultz, E.; Zanin, E.; Wurzenberger, C.; Braun, M.; Rabut, G.; Sironi, L.; Ellenberg, J. Systematic kinetic analysis of mitotic dis- and reassembly of the nuclear pore in living cells. J. Cell Biol. 2008, 180, 857–865.
- Amlacher, S.; Sarges, P.; Flemming, D.; van Noort, V.; Kunze, R.; Devos, D.P.; Arumugam, M.; Bork, P.; Hurt, E. Insight into structure and assembly of the nuclear pore complex by utilizing the genome of a eukaryotic thermophile. Cell 2011, 146, 277–289.
- Teimer, R.; Kosinski, J.; von Appen, A.; Beck, M.; Hurt, E. A short linear motif in scaffold Nup145C connects Y-complex with pre-assembled outer ring Nup82 complex. Nat. Commun. 2017, 8, 1107.
- Hamed, M.; Antonin, W. Dunking into the Lipid Bilayer: How Direct Membrane Binding of Nucleoporins Can Contribute to Nuclear Pore Complex Structure and Assembly. Cells 2021, 10, 3601.
More
Information
Subjects:
Cell Biology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
946
Revisions:
2 times
(View History)
Update Date:
17 Jun 2022
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No