Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Ryza Priatama | -- | 4567 | 2022-05-26 09:52:40 | | | |
| 2 | Conner Chen | Meta information modification | 4567 | 2022-05-27 08:07:38 | | | | |
| 3 | Conner Chen | Meta information modification | 4567 | 2022-05-27 11:05:28 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Priatama, R.; Lee, Y.K.; , .; Park, S.; Park, S.J. Plasma Treatment for Seed Germination and Plant Growth. Encyclopedia. Available online: https://encyclopedia.pub/entry/23405 (accessed on 05 July 2026).
Priatama R, Lee YK, , Park S, Park SJ. Plasma Treatment for Seed Germination and Plant Growth. Encyclopedia. Available at: https://encyclopedia.pub/entry/23405. Accessed July 05, 2026.
Priatama, Ryza, Young Koung Lee, , Seungil Park, Soon Ju Park. "Plasma Treatment for Seed Germination and Plant Growth" Encyclopedia, https://encyclopedia.pub/entry/23405 (accessed July 05, 2026).
Priatama, R., Lee, Y.K., , ., Park, S., & Park, S.J. (2022, May 26). Plasma Treatment for Seed Germination and Plant Growth. In Encyclopedia. https://encyclopedia.pub/entry/23405
Priatama, Ryza, et al. "Plasma Treatment for Seed Germination and Plant Growth." Encyclopedia. Web. 26 May, 2022.
Copy Citation
Low-temperature atmospheric pressure plasma has been used in various fields such as plasma medicine, agriculture, food safety and storage, and food manufacturing. In the field of plasma agriculture, plasma treatment improves seed germination, plant growth, and resistance to abiotic and biotic stresses, allows pesticide removal, and enhances biomass and yield.
low temperature
plasma seed germination
plasma agriculture
molecular mechanism
cold atmospheric plasma
1. Introduction
In recent years, low temperature plasma (LTP), also known as non-thermal plasma (NTP) or cold atmospheric plasma (CAP), has been widely applied in biology. It has broad applications in the field of biology, including seed germination, cultivation, surface sterilization, microorganism decontamination, food manufacturing and processing, wound healing, and food storage. In the field of agriculture, plasma agriculture or plasma farming involves comprehensive application of plasma to process from pre-cultivation until the product reaches the kitchen table. In plant sciences, studies on plasma treatment have been focused on exploring the possible applications, standardization of treatment, and characterization of plasma effects in terms of plant biochemistry [1]. Recently, molecular mechanisms underlying the effects of plasma on seed germination and plant growth have been explored at the cellular level, including gene expression analysis, transcriptome profiling, protein expression analysis, and epigenetics [2][3].
Seed germination is essential for the sustainability and survival of plant species. Germination is a complex process that begins with water imbibition, which triggers physiological processes, ultimately leading to the breaking of seed dormancy [4]. A rapid and uniform germination is one of the key factors for optimum crop cultivation [5]. In agriculture, numerous studies have been conducted to develop methods to promote seed germination that would eventually have a positive impact on crop growth, plant biomass, and yield. Among them, seed priming is a well-established technique for enhancing seed germination efficiency and plant growth [6][7][8]. Seed priming reduces the time required for germination and improves vigor resulting in high crop yields. The direct treatment of seeds with plasma changes the shape of the seed coat resulting in the induction of seed germination, reduced germination time, improved disease resistance, and rapid growth and development [9][10]. In addition, the production of water with altered chemical composition by plasma treatment confers antimicrobial properties, which enable uniform seed germination and induction of germination by its active components, such as reactive species [11]. This positive effect of plasma treatment resulting in seed sterilization, combined with reduced chemical and pesticide usage for pre-cultivation process, is an effective way to reduce the harmful effects of chemicals and pesticides on the environment [12][13].
Plasma treatment has been investigated in seeds of various plant species and ecotypes. In Arabidopsis (a model plant), direct plasma jet treatment promotes seed germination [14]. Moreover, irrigation of Arabidopsis with plasma-activated water (PAW, or plasma treated water, PTW) results in increased germination and an increased number of leaves and flowers [14]. Recently, research on horticultural crops and Arabidopsis is being actively conducted. It was reported that NTP enhances seed germination efficiency, improves surface sterilization of seeds, and promotes long-term growth by increasing seedling biomass, the production of antioxidants and plant hormones and the expression of genes related to defense and drought stress [15][16][17].
In radish, the germination rate was increased, and its growth was promoted when plasma was applied both indirectly—seeds irrigated with PAW—and directly [18][19]. In tomato and pepper, a combined plasma approach was used, in which seeds were exposed for 10 min (min.) to plasma and irrigated with PAW, and the germination rate and plant growth were found to be increased compared with that in samples treated with water [20]. Recently, in ginseng, plasma treatment caused an increase in germination rate and sterilization, and also inhibited the growth of microorganisms exhibiting antifungal activity, especially showing an inhibitory effect on root rot [21]. Therefore, direct and indirect plasma treatments of seeds are considered to positively affect the plant development, including seed germination, growth, and disease prevention. However, optimal conditions in terms of intensity and treatment duration of plasma depending on the variety and ecotype of the crop are crucial factors that would determine the success of this technique for future applications [22][23].
2. Plasma Treatment Method
Plasma is a partially or fully ionized gas that can be ignited at low atmospheric conditions and consists of charged species (electrons and negative and positive ions), neutral species (atomic and/or molecular radicals and non-radicals), electric fields, and photons. One of the earliest applications of plasma for treatment of seeds was studied in early 1960s, when the effects of glow discharge on cotton, wheat, alfalfa, red clover, sweet clover, beans, and several varieties of grass seeds were investigated. It was shown that the plasma treatment influences seed germination, moisture adsorption, and apparently reduces hard-seed content in legumes [24][25][26][27]. Since then, studies on plasma treatment of seeds have been expanded by using various kinds of plasma devices, which allow detailed studies on physical, chemical, and biological mechanisms of plasma that can be triggered by analysis of plasma components [28].
In the past decades, LTP at atmospheric pressure has opened up a new research field in biology and medicine [29][30]. Plasma treatment of seeds has been divided into two methods—direct and indirect—based on the contact of the plasma with the samples. For plasma treatment, plasma sources, such as dielectric barrier discharge (DBD) [31][32][33][34][35][36][37], radio frequency (RF) plasma [38][39][40][41][42][43][44], and atmospheric pressure plasma jet (APPJ) [15][45][46], have been used (Figure 1). The treatment is performed by controlling operating parameters, such as electrode structure, power source (voltage, frequency, and waveform), discharge gas (air, Ar, He, etc.), and other conditions (gas flow, gas pressure, gas temperature, etc.).
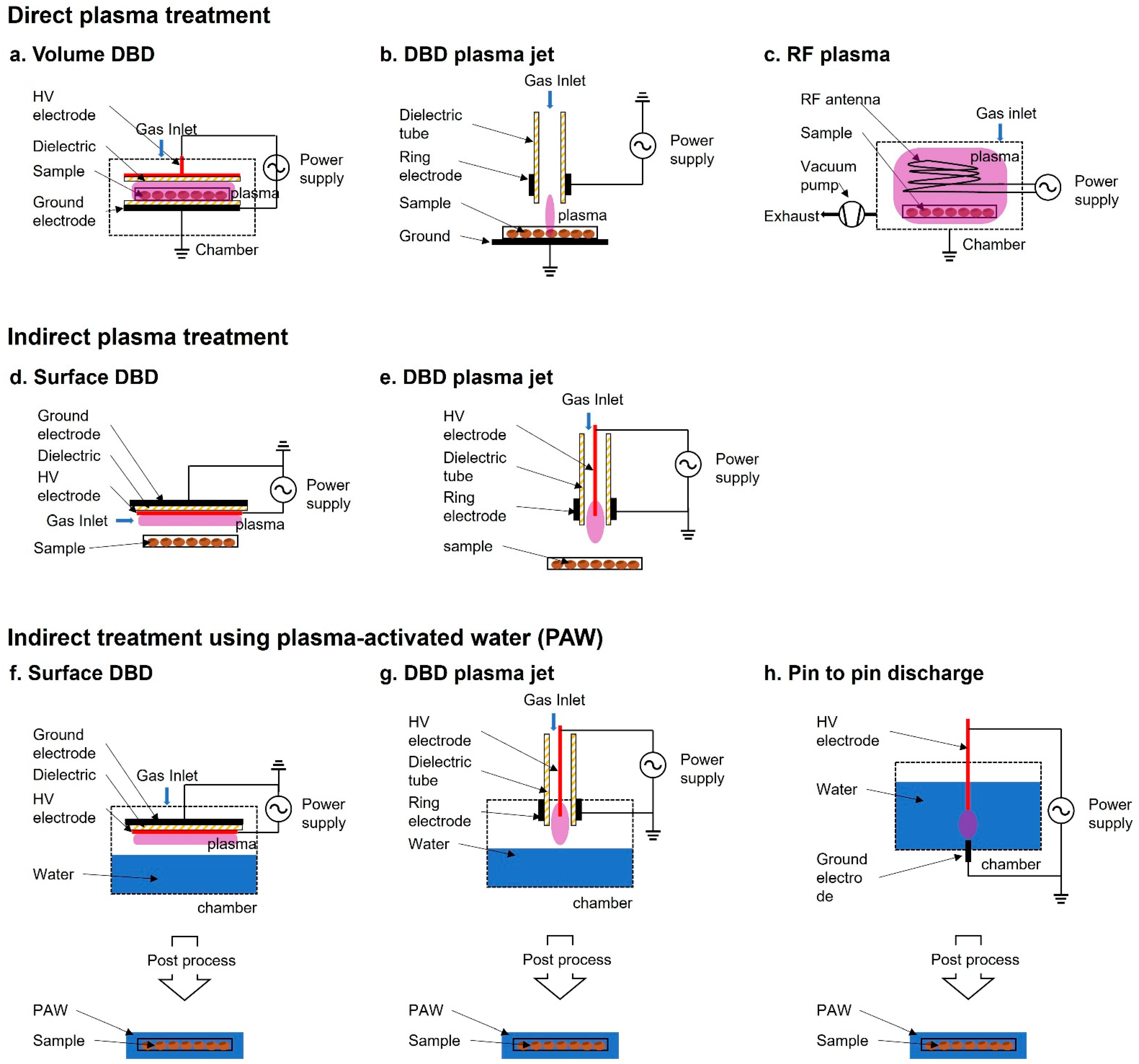
Figure 1. Schematic diagram of plasma devices used for seed treatment. (a–c) Direct plasma treatment: (a) volume dielectric barrier discharge (DBD), (b) DBD plasma jet, (c) Radiofrequency (RF) plasma; (d,e) Indirect plasma treatment where the sample is not directly in contact with the plasma discharge: (d) surface DBD, (e) DBD plasma jet; (f–h) Indirect treatment using plasma-activated water (PAW), (f) surface DBD, (g) DBD plasma jet, (h) pin-to-pin discharge.
In direct plasma treatment, the plasma devices usually consist of a place to plant seeds in the container module, where they are exposed to the plasma generated by the generator and electrode. Because the surface area of Surface DBD (SDBD) is relatively wider compared with that of other plasma devices, it has been more commonly used than other plasma devices for biological applications. For example, SDBD plasma device has been widely used in the germination and growth of Arabidopsis, barley, bell pepper, maize, pea, quinoa, and wheat [33][47][48][49][50][51]. The exposed seeds in the discharge area are directly affected by charged particles, reactive species (such as OH radicals, singlet oxygen, ozone, and hydrogen peroxide), electric fields, and photons (visible/ultraviolet (UV) radiation). A combination of these components is believed to be the main factor that promotes seed germination and growth. During the exposure, the seed surface interacts with the short lived and long-lived radicals, which appear due to secondary reactions.
In indirect treatment, the sample is not exposed to the plasma itself. The plasma does not directly affect the samples, but a gas-phase active species generated by the plasma and PAW affect the sample. Plasma-treated water causes changes in the physicochemical properties and PAW participates in the signaling pathway and eventually promotes seed germination, root and vegetative growth, and plant reproduction [52][53].
PAW, especially under atmospheric conditions, is known to change the electrical conductivity, pH, concentration of nitrite (NO2−), nitrate (NO3−), ozone (O3), and hydrogen peroxide (H2O2) [54]. These changes in physicochemical properties and components mainly contribute to the benefits of plasma treatment for seed germination and plant growth. Most research on seed treatment has been focused on the inhibition of microbial growth on the seed surface. The application of PAW for inhibition of microorganisms is usually linked to the increasing acidity with the duration of plasma treatment [55][56][57]. Moreover, the reactive oxygen and nitrogen species (RONS) content is increased, which also inhibits microbial growth that later affects seed germination and RONS functions in seed priming [10].
In addition to seed priming, the high nitrate and nitrite content in PAW is believed to be the major factor contributing to the improvement of plant growth under PAW irrigation because it can act as a substitute for nitrogen source. Hence, the concept of “plasma-fertilizer” was established. This concept is considered important because nitrogen is the backbone of all metabolic processes that directly affect plant growth. In addition, “plasma fertilizer” is one of the eco-friendly alternatives of nitrogen source that reduces the disadvantages associated with the use of chemical fertilizers. Therefore, it is not surprising that this is the major topic for future studies on plasma treatment for plant cultivation [58][59][60][61].
3. Chemical and Physical Effects of Plasma Treatment
Direct plasma treatment of the seed surface is the most common treatment performed in seed germination studies. When the plasma contacts the seed surface, it changes the seed coat, resulting in reduced germination time and rapid growth and development [14][62]. The positive effects of plasma treatment of seeds have been observed in several species of horticultural crops [15]. Plasma studies in horticultural crops show that atmospheric plasma increases the germination rate and growth rate in radish, pepper, and tomato in correlation with the duration of treatment [20]. In addition to the germination of tomato seeds, use of helium plasma for seed priming increases its overall growth compared with that in control [15]. It is well known that plasma treatment of seeds is mainly affected by RONS. The production of RONS from plasma is also regarded as a major factor in enhancing seed germination, seedling growth, and plant defense. In addition, seed surface or seed coat changes are categorized as mechanical changes. Moreover, factors such as seed sterilization, heat, UV irradiation, ionization, and electromagnetic fields generated by plasma devices are also believed to play roles in seed germination induced by plasma treatment. Figure 2.
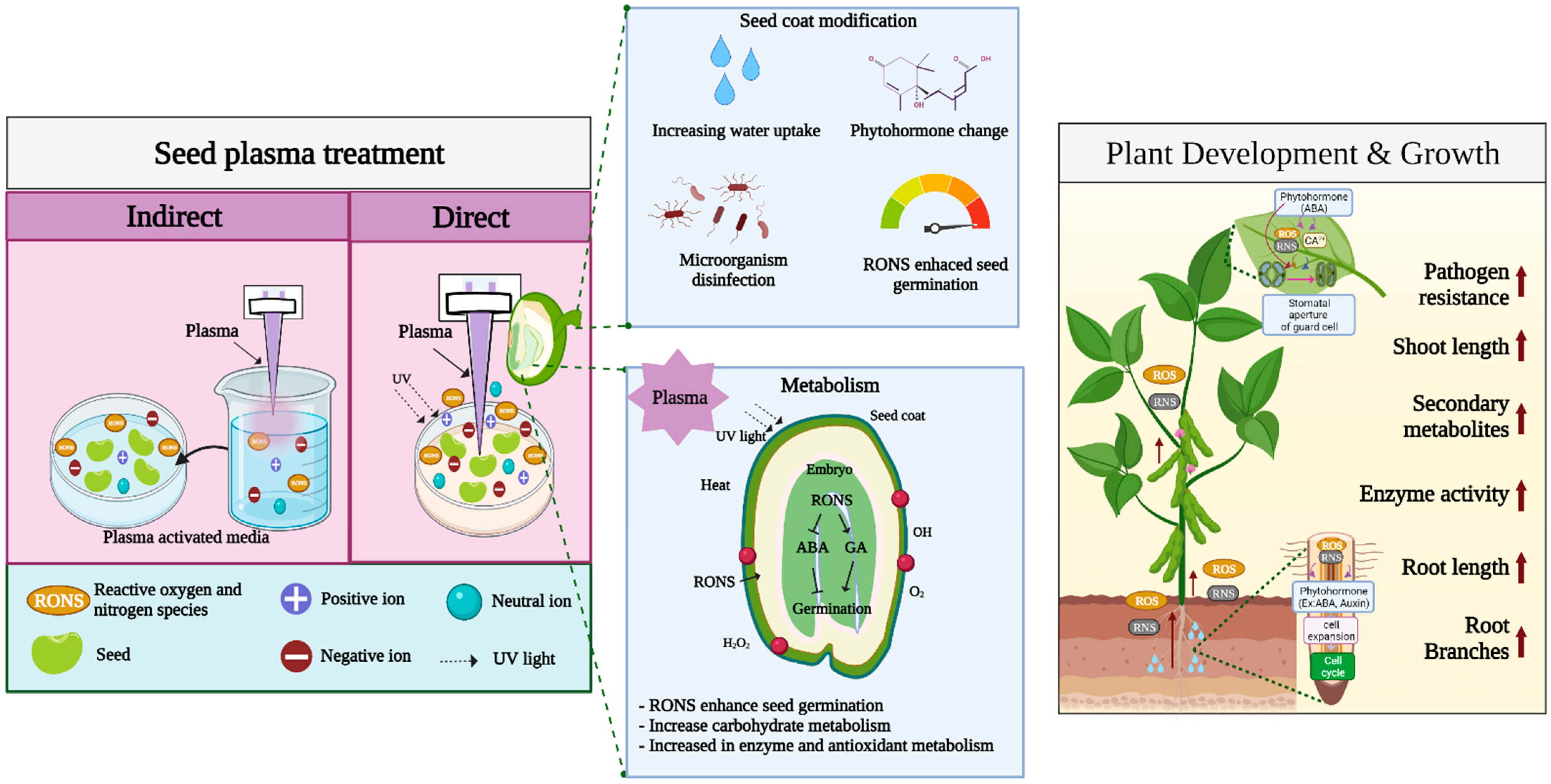
Figure 2. Overview of plasma treatment effects on seed germination and plant growth.
3.1. Chemical Effects of Plasma Treatment
Several terms have been used in literature for reactive species to describe the oxygen radicals and non-radicals. Reactive species are also grouped into reactive oxygen species (ROS: O2−, H2O2, O3, etc.) and reactive nitrogen species (RNS: •NO, •NO2, ONOO−, etc.). Some common reactive species include ozone (O3), hydrogen peroxide (H2O2), superoxide anion, peroxyl, nitric oxide, nitrogen dioxide, and peroxynitrite [63][64][65]. Reactive species generated from plasma treatment are believed to be the major factors affecting seed germination and plant growth. However, mechanisms underlying the effects of ROS and RNS on seed germination and development are not fully understood. There are few hypotheses about how the external ROS affect seed germination. One hypothesis is that external ROS are detected and perceived by the cells in seeds that induce signal transduction from the outer layer of the seeds. The other hypothesis is that during imbibition, water is the key factor in the absorption of ROS into the cell layers of the seeds. Thus, it increases the respiration of seeds and triggers a chain reaction of sugar oxidation to release metabolic energy in the form of ATP [47]. Therefore, the involvement of ROS in the respiration pathway is considered a primary and secondary trigger in seeds that causes transition from dormancy to metabolic activity.
A previous study showed the presence of external ROS in certain amounts in water during imbibition and also showed that wet seeds may trigger faster signaling in the intercellular process; however, the effect of ROS on dry seeds is hardly understood. There is a possibility that the effect of ROS on dry seeds is minimized or delayed. During plasma treatment, ROS penetrates the seeds, but no specific mechanism occurs until the start of imbibition [66]. However, how ROS are stored during the period before imbibition remains unknown. In addition, this theory does not fully explain how long-term storage after plasma treatment still a positive effect on the germination and growth of seeds compared with those of untreated seeds, especially when ROS are known to be mostly short-lived [67]. Biochemical changes in plasma-treated seeds apparently continue to occur even long after the seeds are treated [20][68]. These changes are related to gene expression, the oxidative process, protein concentration and hormones. It is believed that plasma treatment increases the pore size of the seed coat, which increases water imbibition and ROS absorption, leading to genetic regulation of seeds [69]. Other possible ways by which seeds and plants might absorb RONS generated from plasma treatment could be through the bypassing of protein channels, named aquaporins, which are mainly used for water transport [70]. It is important to note that, in addition to transport via aquaporins, ozone is absorbed through stomata in the seedlings and mature plants. Therefore, it has been observed that the accumulation of ROS in leaves through stomata and micropores is the main pathway by which ROS can travel further to other plant tissues. It was also shown that excessive absorption of ROS by plants could result in chloroplast damage [71].
A relatively small amount of H2O2 (0.12 ppm) in PAW, generated from a plasma device, increased the seed germination rate in tomato and pepper seeds [20]. In Arabidopsis, PAW containing 17–25.5 mg/L H2O2 had a positive effect on germination and seedling growth [14]. Among RNS, low nitrate concentration (100 ppm or less) enhances seed germination and seedling growth in plants, but the growth tends to be inhibited above 100 ppm [20][72][73][74]. However, plants apparently have their own nitrate and ROS sensitivity and show a dosage-dependent growth pattern.
One of the prominent members of reactive species, which is abundantly detected in plasma treatment, is hydrogen peroxide (H2O2). A previous study on the involvement of H2O2 in seed germination and seedling growth revealed its role in the absorption mechanism, signaling pathway, regulation of gene expression, protein modification, and other related factors [75]. The absorption of H2O2 into the cells occurs mainly through diffusion and is dependent on the anion channels; inside the cell, it breaks down into singlet oxygen and hydroxyl, thus, allowing easy transfer between cells [70][76]. During germination, H2O2 mediates the regulation of abscisic acid (ABA) catabolism and gibberellic acid (GA) biosynthesis [77]. In addition, upon exposure of seeds to external H2O2, the endogenous H2O2 levels also increase and induce several oxidative pathways, such as carbonylation and lipid peroxidation [78]. Moreover, the presence of H2O2 in cells is regarded as a priming factor that involves complex changes in the proteome, transcriptome, and hormone levels [78].
Ozone, as one of the major ROS, was shown to be responsible for improved seed germination and induction of protein expression in seeds [79][80]. Ozone generated during the presence of UV radiation can eventually generate superoxide and hydroxyl radicals in the seed coat, which could be one of the main reasons for how the combination of external physical damage of the seed coat combined with chemical stimulus of the accumulated radicals works synergistically to increase germination. However, it is important to note that different device configurations produce different concentrations of ozone. Moreover, the purpose of treatment also determines the ozone concentration required. Postharvest treatment with 0.3 ppm ozone in combination with cold storage could inhibit the decay process and reduced severe infection in peach and table grapes [81][82]. In strawberry, plasma treatment using different sources of gas has been investigated; it was observed that plasma treatment for 5 min could produce 600–2800 ppm ozone that had a positive effect on microbial disinfection and strawberry freshness [83]. Similarly, various ozone concentrations were investigated in the plasma treatment of seeds. In Arabidopsis seeds, the effects of treatment with 200 ppm ozone, generated from a plasma device, for 10 min on seed coat modification were studied [84]. A low concentration of ozone (~1–5 ppm) is effective in plant growth by killing larvae in the soil and in fresh cut green leaf lettuce [85]. Interestingly, a similar concentration of ozone (~1–4 ppm) generated from surface discharge successfully reduced the number of nematodes and induced plant growth [86].
The biochemical mechanism underlying the effects of plasma treatment on seeds is very closely related to the metabolism of antioxidant enzymes. The seed coat may contain proteins or enzymes, such as NADPH oxidase, superoxide dismutase (SOD), and peroxidase (POD), which can convert the substrate into signaling molecules such as H2O2. Plasma treatment of wheat seedlings resulted in increased isoenzyme activities, such as POD and phenylalanine ammonia lyase (PAL), which are crucial for the production of polyphenols, which also participate in plant defense [87][88].
Another area of focus in plasma treatment is the potential application of RNS produced from plasma–liquid interaction as a liquid fertilizer for plant growth. Various approaches have been explored to obtain the most suitable device and treatment method for the production of high amounts of RNS in the solution. For example, a large volume of glow discharge has been tested as a liquid fertilizer in radish, tomatoes, and marigolds [89]; bubble discharge has been investigated in the cultivation of spinach, radish, Brassica rapa, and strawberry [90][91][92]; a plasma jet has been used for plasma-assisted nitrogen fixation for corn [93]. Collectively, these reports demonstrate the potential of plasma fertilizers as an alternative and a more eco-friendly approach for nitrogen source for plant cultivation. However, one of the challenges in plasma-assisted nitrogen fixation is the low pH or increased acidity of the solution treated with plasma, which damages the seed and plant exposed to such solutions. Plant growth is limited under acidic environment [94]. Therefore, studies on controlling the balance and on methods to overcome the acidity of plasma-activated solutions are being considered a priority in the plasma field. Lamichhane et al. recently demonstrated an innovative approach to control the acidity of plasma-treated water using a combination of chemical additives including Mg, Al, or Zn, which could neutralize via the reduction in pH [93]. Moreover, the presence of these additives increases the rate of reduction of nitrogen to ammonia, which results in the improvement of germination rate and seedling growth [93].
Inactivation of microbes by plasma treatment has been used as the fundamental technology in medicine and food processing [95][96][97]. Microbial inactivation, resulting from plasma treatment of seeds, also plays important roles in germination. The seed surface is usually exposed to the environment, which contains many types of particles, contaminants, and microbes that could have negative effects on seed germination. For example, in grain crops, such as rice, wheat, oat, and barley, the growth of Fusarium on the seed surface affects germination [98]. It is known that fungal infections on seeds often damage their viability and potentially reduce the yield. Moreover, fungal pathogen on seeds can also lead to a seed-borne infection, which can cause abundant yield loss. The plasma treatment of seeds has been shown to have positive effects on seed sterilization, including removal of fungal spore on the seeds. In rice, direct treatment using micro corona discharge of SDBD can inactivate the microorganisms on the husk, which leads to higher germination compared with that in untreated seeds [99]. This finding emphasizes the incorporation of Ar and air in plasma that would result in the production of ROS and RNS, which decontaminate and inactivate fungi on the seed surface. Another study using indirect treatment with arch discharge plasma showed successful inactivation of Fusarium fujikuroi (a fungal pathogen) in a submerged rice seed suspension; however, fungal spores are more effectively inactivated using ultrasonic waves as a source of ozone and shock-wave [100]. Collectively, these two examples show that both direct and indirect treatments are effective in inhibiting the fungus through the production of ROS and RNS. Moreover, positive results for microbial inactivation or sterilization of seeds were also confirmed in many different seeds, such as wheat, barley, oat, lentils, maize, chickpea, sunflower, and scots pine [12][101][102][103][104].
The mechanism underlying the effects of plasma treatment in the inactivation of microorganisms has been well-investigated using different device types, exposure times, power, gas sources, and other factors. Compared with conventional seed sterilization using active chemicals, the mechanism of plasma disinfection is often considered complex due to variations in devices and plasma components [105]. However, the general agreement for the use of plasma treatment for disinfection is due to the production of reactive species, exposure to which triggers a complex sequence of events in the microbe, resulting in the antimicrobial activity [106]. For example, exposure to ROS directly triggers molecular damage in cells, including DNA breakage, lipid peroxidation, and carbonylation of proteins [107][108]. Moreover, combined action of RNS and ROS is important for antibacterial activity. The effect of plasma with high content of ROS and RNS is more than that of plasma-treated water on the antimicrobial activity [109]. UV exposure also plays an important role, especially in the inactivation of bacteria using plasma treatment. It is known that mechanism of action of UV exposure may be related to several specific mechanisms, including direct destruction of genetic material, breaking of chemical bonds in organic compounds, and the phenomenon of plasma etching (UV-induced etching) [110][111][112].
Other applications of plasma treatment include the inactivation of viruses. In the field of plasma biomedicine, the inactivation of coronavirus using plasma devices has recently garnered a lot of attention and proved to be effective in treating viral infection and associated diseases [113][114][115]. In plants, potential applications of plasma treatment of virus-related diseases have been reported. Potato virus Y (PVY) homogenized in water was successfully inactivated by plasma treatment for 1 min [116]. A sample of pepper mild mottle virus (PMMoV) in water was also successfully inactivated using 99% argon and 1% oxygen plasma jet in 5 and 3 min [117]. As for the inactivation of bacteria, the mechanism of virus inactivation is primarily through the production of reactive species (ROS and RNS) with various physical effects, and the treatment could damage the virus particle and degrade viral DNA/RNA [118][119].
3.2. Physical Effects of Plasma Treatment
Plasma treatment of seeds, especially direct treatment, is considered to have a similar principle to plasma etching on the seed surface [120]. It is shown that seed coat modifications are presumably highly dependent on the type of plasma devices, power, and duration of the treatment. For example, treatment of Arabidopsis, wheat, radish, and oat seeds showed no distinct surface modifications based on seed coat morphology [121][122][123][124]. However, other studies revealed seed coat degradation by plasma treatment, which was observed using scanning electron microscope (SEM), in different plant seeds such as Arabidopsis, cotton, wheat, mimosa (Mimosa caesalpiniaefolia), erythrina (Erythrina velutina), pea, and onion [34][35][125][126][127][128][129]. Therefore, the effect of plasma treatment on seed germination may be due to the mechanical factors of seed coat modification, especially when treated with appropriate plasma device and configuration.
The seed coat protects the seed from the external environment and regulates the water absorption. Imbibition must occur in the correct ratio; otherwise, seeds may be damaged if imbibition is too slow or too fast [69]. Plasma treatment affects seed germination differently in different seeds of various species and different plant families, even for different variety and ecotype, and generally, a different setup is needed for optimum treatment condition. The seed coat consists of cuticle, epidermis, hypodermis, and parenchyma cells. The degradation of cuticle layers will allow water to be absorbed further into inner layers. Plasma treatment helps in the removal of lipid layers on the cuticle and epidermis, which accelerate the germination [130]. In Arabidopsis seeds, modifications in the composition of lipid compounds in the seed coat was detected after plasma treatment [37]. Interestingly, Arabidopsis seed coat mutants, gl2 and gpat5, and Col-0 were examined under plasma treatment; the germination rate was increased in plasma-treated seeds, even under osmotic and saline stress conditions, whereas the germination ratio in gpat5, with defective cuticle layers on the seeds, was not rescued by plasma treatment due to less permeability and more sensitivity to plasma processing. POD activity in testa and endosperm tissues was detected, where the major POD function occurred in ruptured seeds. This shows that in plasma-treated seeds, the structure and composition of lipid compounds are changed before germination and metabolism changes after germination [37].
Seed surface modifications are usually investigated using light microscope or SEM. In addition, biochemical analysis is also performed to confirm the differences in seed coat content, such as lignin, cutin, polysaccharides, and other ROS-related proteins. Indirect treatment using plasma-treated water was also performed to determine the seed wettability, which showed possibilities of combined mechanism involving seed perforation and lower water tension that increases the surface area and ability of imbibition. Seed coat modifications are closely related to water permeability and water affinity on seed surface, also known seed wettability [131]. The increase in seed wettability improves the permeability of seeds to water, and thus, the imbibition process is accelerated. The wettability has been known to be majorly affected by the morphology and chemistry of seeds. Morphologically, plasma treatment causes surface etching or surface erosion that increases the roughness of the seed surface, resulting in the increase in seed volume ratio, and thus, wettability is increased. Chemically, the interaction of plasma and seed coat components affects the organic polymers in seeds, for example, it results in degradation of cutin and wax layers, thus reducing the hydrophobic activity of the seed coat and increasing water permeability [131][132][133].
The heat effect of plasma treatment was examined by comparing the plasma treatment with the heat plate treatment, which shows different effects on germination [121]. It was shown that heat is not the main factor contributing to the increase in germination of plasma-treated seeds. Although plasma treatment is regarded as “non thermal plasma or cold plasma”, the slight increase in temperature in plasma-treated seeds cannot be ignored as it may still affect the germination ratio when combined with other factors. Heat shock proteins (HSPs) are one of the important regulators of plant metabolism. They sense temperature changes, perceive signaling, and respond to protect other proteins from stress-induced damage [134][135][136]. Iranbakhsh et al. confirmed that heat resulted in the induction of HSP expression in plasma-treated wheat seeds [36]. However, it is difficult to distinguish whether heat increased the scarification of seeds, which led to the induction of germination and indirectly increased the expression of HSPs, or whether the heat generated by plasma treatment directly induced HSPs [18]. Moreover, in a study on plasma-treated sunflower seeds, no change in the expression of HSPs was evident in proteome profiling and an ambiguous result that the treatment may not induce significant accumulation of pathogenesis-related (PR) proteins or HSPs was obtained [42]. Remarkably, a recent study on the expression of HSP genes in maize grain treated with diffuse coplanar surface barrier discharge (DCSBD) showed induction of several HSP genes, including HSP101, HSP70, and HSF17, that affected grain vitality and seedling growth [137].
Ultraviolet light is regarded as one of the main factors in plasma sterilization, especially in the inactivation of surface microbes [96][138]. Single spectrum UV lights, such as UV-A, UV-B, and UV-C, have been shown to increase seed germination but act differently during seedling and plant growth. Exposure of Amberboa ramose seeds to UV-A resulted in increased germination and growth during plant development [139]. UV-B was shown to increase the germination rate of safflower (Carthamus tinctorious), radish, cabbage, kale, and agave seeds [140]. However, prolonged exposure to UV-B inhibits the growth of seedlings [140][141]. UV-C treatment in maize and sugar beet was also shown to increase germination rate and seedling growth [142]. During plasma treatment, the possibility that UV light directly affects seed germination is considered very low due to its short exposure duration and low intensity of irradiation from the plasma sources. In some studies, it was proposed that UV irradiation due to plasma treatment may have indirect effects combined with RONS interaction and not by improvement in seed wettability [125][143]. In addition, the nature of UV exposure could induce DNA damage, which could also affect germination and seedling growth. Short-term exposure of seeds and seedlings to UV could induce their growth by regulation of stress response, especially when the irradiation is perceived by the photoreceptor, leading to increased cell metabolism, including cell differentiation, division and elongation [144].
References
- Starič, P.; Vogel-Mikuš, K.; Mozetič, M.; Junkar, I. Effects of Nonthermal Plasma on Morphology, Genetics and Physiology of Seeds: A Review. Plants 2020, 9, 1736.
- Yan, D.; Lin, L.; Zvansky, M.; Kohanzadeh, L.; Taban, S.; Chriqui, S.; Keidar, M. Improving Seed Germination by Cold Atmospheric Plasma. Plasma 2022, 5, 98–110.
- Ikmal Misnal, M.F.; Redzuan, N.; Firdaus Zainal, M.N.; Raja Ibrahim, R.K.; Ahmad, N.; Agun, L. Emerging Cold Plasma Treatment on Rice Grains: A Mini Review. Chemosphere 2021, 274, 129972.
- Nonogaki, H. Seed Dormancy and Germination—Emerging Mechanisms and New Hypotheses. Front. Plant Sci. 2014, 5, 233.
- Tuan, P.A.; Sun, M.; Nguyen, T.-N.; Park, S.; Ayele, B.T. Molecular Mechanisms of Seed Germination. In Sprouted Grains; Feng, H., Nemzer, B., DeVries, J.W., Eds.; AACC International Press: St. Paul, MN, USA, 2019; pp. 1–24. ISBN 978-0-12-811525-1.
- Lutts, S.; Benincasa, P.; Wojtyla, L.; Kubala, S.; Pace, R.; Lechowska, K. Seed Priming: New Comprehensive Approaches for an Old Empirical Technique, New Challenges in Seed Biology-Basic and Translational Research Driving Seed Technology; InTechOpen: Rijeka, Croatia, 2016.
- Pawar, V.A.; Laware, S.L. Seed priming A critical review. Int. J. Sci. Res. Biol. Sci. 2018, 31, 94–101.
- Zulfiqar, F. Effect of Seed Priming on Horticultural Crops. Sci. Hortic. 2021, 286, 110197.
- Rasooli, Z.; Barzin, G.; Mahabadi, T.D.; Entezari, M. Stimulating Effects of Cold Plasma Seed Priming on Germination and Seedling Growth of Cumin Plant. South Afr. J. Bot. 2021, 142, 106–113.
- Nalwa, C.; Thakur, A.K.; Vikram, A.; Rane, R.; Vaid, A. Studies on plasma treatment and priming of seeds of bell pepper (Capsicum annuum L.). J. Appl. Nat. Sci. 2017, 9, 1505–1509.
- Darmanin, M.; Kozak, D.; de Oliveira Mallia, J.; Blundell, R.; Gatt, R.; Valdramidis, V.P. Generation of Plasma Functionalized Water: Antimicrobial Assessment and Impact on Seed Germination. Food Control 2020, 113, 107168.
- Selcuk, M.; Oksuz, L.; Basaran, P. Decontamination of Grains and Legumes Infected with Aspergillus Spp. and Penicillum Spp. by Cold Plasma Treatment. Bioresour. Technol. 2008, 99, 5104–5109.
- Kim, K.-H.; Kabir, E.; Jahan, S.A. Exposure to Pesticides and the Associated Human Health Effects. Sci. Total Environ. 2017, 575, 525–535.
- Bafoil, M.; Jemmat, A.; Martinez, Y.; Merbahi, N.; Eichwald, O.; Dunand, C.; Yousfi, M. Effects of Low Temperature Plasmas and Plasma Activated Waters on Arabidopsis Thaliana Germination and Growth. PLoS ONE 2018, 13, e0195512.
- Adhikari, B.; Adhikari, M.; Ghimire, B.; Adhikari, B.C.; Park, G.; Choi, E.H. Cold Plasma Seed Priming Modulates Growth, Redox Homeostasis and Stress Response by Inducing Reactive Species in Tomato (Solanum lycopersicum). Free Radic. Biol. Med. 2020, 156, 57–69.
- Adhikari, B.; Adhikari, M.; Park, G. The Effects of Plasma on Plant Growth, Development, and Sustainability. Appl. Sci. 2020, 10, 6045.
- Adhikari, B.; Pangomm, K.; Veerana, M.; Mitra, S.; Park, G. Plant Disease Control by Non-Thermal Atmospheric-Pressure Plasma. Front. Plant Sci. 2020, 11, 77.
- Koga, K.; Attri, P.; Kamataki, K.; Itagaki, N.; Shiratani, M.; Mildažiene, V. Impact of Radish Sprouts Seeds Coat Color on the Electron Paramagnetic Resonance Signals after Plasma Treatment. Jpn. J. Appl. Phys. 2020, 59, SHHF01.
- Attri, P.; Ishikawa, K.; Okumura, T.; Koga, K.; Shiratani, M.; Mildaziene, V.; Ishikawa, K.; Okumura, T.; Koga, K.; Shiratani, M.; et al. Impact of Seed Color and Storage Time on the Radish Seed Germination and Sprout Growth in Plasma Agriculture. Sci. Rep. 2021, 11, 2539.
- Sivachandiran, L.; Khacef, A. Enhanced Seed Germination and Plant Growth by Atmospheric Pressure Cold Air Plasma: Combined Effect of Seed and Water Treatment. RSC Adv. 2017, 7, 1822–1832.
- Lee, Y.; Lee, Y.Y.; Kim, Y.S.; Balaraju, K.; Mok, Y.S.; Yoo, S.J.; Jeon, Y. Enhancement of Seed Germination and Microbial Disinfection on Ginseng by Cold Plasma Treatment. J. Ginseng Res. 2021, 45, 519–526.
- Song, J.-S.; Kim, S.B.; Ryu, S.; Oh, J.; Kim, D.-S. Emerging Plasma Technology That Alleviates Crop Stress During the Early Growth Stages of Plants: A Review. Front. Plant Sci. 2020, 11, 988.
- Waskow, A.; Avino, F.; Howling, A.; Furno, I. Entering the Plasma Agriculture Field: An Attempt to Standardize Protocols for Plasma Treatment of Seeds. Plasma Process. Polym. 2022, 19, 2100152.
- Nelson, S.O.; Stetson, L.E.; Stone, R.B.; Webb, J.C.; Pettibone, C.A.; Works, D.W.; Kehr, W.R.; VanRiper, G.E. Comparison of Infrared, Radiofrequency, and Gas-Plasma Treatments of Alfalfa Seed for Hard-Seed Reduction. Trans. ASAE 1964, 7, 276–280.
- Alan, C. Pettibone Some Effects of Gas-Plasma Radiation on Seeds. Trans. ASAE 1965, 8, 319–321.
- Webb, J.C.; Stone, B.; McDow, R.J.J. Response of Cottonseed to Audiofrequency Gas Plasma. Trans. ASAE 1966, 9, 872–874.
- Nelson, S.O.; Kehr, W.R.; Stetson, L.E.; Stone, R.B.; Webb, J.C. Alfalfa Seed Germination Response to Electrical Treatments1. Crop Sci. 1977, 17, 863–866.
- Ranieri, P.; Sponsel, N.; Kizer, J.; Rojas-Pierce, M.; Hernández, R.; Gatiboni, L.; Grunden, A.; Stapelmann, K. Plasma Agriculture: Review from the Perspective of the Plant and Its Ecosystem. Plasma Process. Polym. 2021, 18, 2000162.
- Laroussi, M.; Bekeschus, S.; Keidar, M.; Bogaerts, A.; Lu, X.; Hori, M.; Stapelmann, K.; Miller, V.; Reuter, S.; Laux, C.; et al. Low Temperature Plasma for Biology, Hygiene, and Medicine: Perspective and Roadmap. In IEEE Transactions on Radiation and Plasma Medical Sciences; IEEE: Piscataway, NJ, USA, 2021.
- Bruggeman, P.; Brandenburg, R. Atmospheric Pressure Discharge Filaments and Microplasmas: Physics, Chemistry and Diagnostics. J. Phys. Appl. Phys. 2013, 46, 464001.
- Gómez-Ramírez, A.; López-Santos, C.; Cantos, M.; García, J.L.; Molina, R.; Cotrino, J. Surface chemistry and germination improvement of Quinoa seeds subjected to plasma activation. Sci. Rep. 2017, 7, 1–12.
- Meng, Y.; Qu, G.; Wang, T.; Sun, Q.; Liang, D.; Hu, S. Enhancement of Germination and Seedling Growth of Wheat Seed Using Dielectric Barrier Discharge Plasma with Various Gas Sources. Plasma Chem. Plasma Proc. 2017, 37, 1105–1119.
- Rahman, M.M.; Sajib, S.A.; Rahi, M.S.; Tahura, S.; Roy, N.C.; Parvez, S.; Reza, M.A.; Talukder, M.R.; Kabir, A.H. Mechanisms and Signaling Associated with LPDBD Plasma Mediated Growth Improvement in Wheat. Sci. Rep. 2018, 8, 10498.
- Li, Y.; Wang, T.; Meng, Y.; Qu, G.; Sun, Q.; Liang, D. Air Atmospheric Dielectric Barrier Discharge Plasma Induced Germination and Growth Enhancement of Wheat Seed. Plasma Chem. Plasma Proc. 2017, 37, 1621–1634.
- Stolárik, T.; Henselová, M.; Martinka, M.; Novák, O.; Zahoranová, A.; Černák, M. Effect of Low-Temperature Plasma on the Structure of Seeds, Growth and Metabolism of Endogenous Phytohormones in Pea (Pisum sativum L.). Plasma Chem. Plasma Process. 2015, 35, 659–676.
- Iranbakhsh, A.; Ardebili, N.O.; Ardebili, Z.O.; Shafaati, M.; Ghoranneviss, M. Non-Thermal Plasma Induced Expression of Heat Shock Factor A4A and Improved Wheat (Triticum aestivum L.) Growth and Resistance against Salt Stress. Plasma Chem. Plasma Proc. 2018, 38, 29–44.
- Bafoil, M.; Le Ru, A.; Merbahi, N.; Eichwald, O.; Dunand, C.; Yousfi, M. New Insights of Low-Temperature Plasma Effects on Germination of Three Genotypes of Arabidopsis Thaliana Seeds under Osmotic and Saline Stresses. Sci. Rep. 2019, 9, 1–10.
- Hayashi, N.; Ono, R.; Nakano, R.; Shiratani, M.; Tashiro, K.; Kuhara, S.; Yasuda, K.; Hagiwara, H. DNA Microarray Analysis of Plant Seeds Irradiated by Active Oxygen Species in Oxygen Plasma. Plasma Med. 2016, 6.
- Nakano, R.; Tashiro, K.; Aijima, R.; Hayashi, N. Effect of Oxygen Plasma Irradiation on Gene Expression in Plant Seeds Induced by Active Oxygen Species. Plasma Med. 2016, 6, 303–313.
- Bormashenko, E.; Shapira, Y.; Grynyov, R.; Whyman, G.; Bormashenko, Y.; Drori, E. Interaction of Cold Radiofrequency Plasma with Seeds of Beans (Phaseolus vulgaris). J. Exp. Bot. 2015, 66, 4013–4021.
- Li, L.; Li, J.; Shen, M.; Hou, J.; Shao, H.; Dong, Y. Improving Seed Germination and Peanut Yields by Cold Plasma Treatment. Plasma Sci. Technol. 2016, 18, 1027–1033.
- Mildažiene, V.; Aleknavičiute, V.; Žukiene, R.; PauŽaite, G.; Naučiene, Z.; Filatova, I. Treatment of Common Sunflower (Helianthus annus L.) Seeds with Radio-Frequency Electromagnetic Field and Cold Plasma Induces Changes in Seed Phytohormone Balance, Seedling Development and Leaf Protein Expression. Sci. Rep. 2019, 9, 6437.
- Saberi, M.; Modarres-Sanavy, S.A.M.; Zare, R.; Ghomi, H. Amelioration of Photosynthesis and Quality of Wheat under Non-Thermal Radio Frequency Plasma Treatment. Sci. Rep. 2018, 8, 11655.
- Volkov, A.G.; Bookal, A.; Hairston, J.S.; Patel, D.; Volkov, A.G.; Bookal, A.; Hairston, J.S.; Patel, D. Radio Frequency Plasma Capacitor Can Increase Rates of Seeds Imbibition, Germination, and Radicle Growth. Funct. Plant Biol. 2020, 48, 312–320.
- Zhou, R.; Zhou, R.; Zhang, X.; Zhuang, J.; Yang, S.; Bazaka, K.; Ken Ostrikov, K. Effects of Atmospheric-Pressure N2, He, Air, and O2 Microplasmas on Mung Bean Seed Germination and Seedling Growth. Sci. Rep. 2016, 6, 32603.
- De Groot, G.J.J.B.; Hundt, A.; Murphy, A.B.; Bange, M.P.; Mai-Prochnow, A. Cold Plasma Treatment for Cotton Seed Germination Improvement. Sci. Rep. 2018, 8, 14372.
- Billah, M.; Sajib, S.A.; Roy, N.C.; Rashid, M.M.; Reza, M.A.; Hasan, M.M.; Talukder, M.R. Effects of DBD Air Plasma Treatment on the Enhancement of Black Gram (Vigna mungo L.) Seed Germination and Growth. Arch. Biochem. Biophys. 2020, 681, 108253.
- Islam, S.; Omar, F.B.; Sajib, S.A.; Roy, N.C.; Reza, A.; Hasan, M. Effects of LPDBD Plasma and Plasma Activated Water on Germination and Growth in Rapeseed (Brassica napus). Gesunde Pflanz. 2019, 71, 175–185.
- Tounekti, T.; Mujahid, Z.U.I.; Khemira, H. Non-Thermal Dielectric Barrier Discharge (DBD) Plasma Affects Germination of Coffee and Grape Seeds. In AIP Conference Proceedings; AIP Publishing LLC: Melville, NY, USA, 2018.
- Pawłat, J.; Starek, A.; Sujak, A.; Terebun, P.; Kwiatkowski, M.; Budzeń, M. Effects of Atmospheric Pressure Plasma Jet Operating with DBD on Lavatera Thuringiaca L. Seeds’ Germination. PLoS ONE 2018, 13, e0194349.
- Liu, B.; Honnorat, B.; Yang, H.; Arancibia, J.; Rajjou, L.; Rousseau, A. Non-Thermal DBD Plasma Array on Seed Germination of Different Plant Species. J. Phys. Appl. Phys. 2019, 52, 025401.
- Bradu, C.; Kutasi, K.; Magureanu, M.; Puač, N.; Živković, S. Reactive Nitrogen Species in Plasma-Activated Water: Generation, Chemistry and Application in Agriculture. J. Phys. Appl. Phys. 2020, 53, 223001.
- Al-Sharify, Z.T.; Al-Sharify, T.A.; al-Obaidy Baker, W.; al-Azawi, A.M. Investigative Study on the Interaction and Applications of Plasma Activated Water(PAW). IOP Conf. Ser. Mater. Sci. Eng. 2020, 870, 012042.
- Thirumdas, R.; Kothakota, A.; Annapure, U.; Siliveru, K.; Blundell, R.; Gatt, R.; Valdramidis, V.P. Plasma Activated Water (PAW): Chemistry, Physico-Chemical Properties, Applications in Food and Agriculture. Trends Food Sci. Technol. 2018, 77, 21–31.
- Šimečková, J.; Krčma, F.; Klofáč, D.; Dostál, L.; Kozáková, Z. Influence of Plasma-Activated Water on Physical and Physical–Chemical Soil Properties. Water 2020, 12, 2357.
- Traylor, M.J.; Pavlovich, M.J.; Karim, S.; Hait, P.; Sakiyama, Y.; Clark, D.S.; Graves, D.B. Long-Term Antibacterial Efficacy of Air Plasma-Activated Water. J. Phys. Appl. Phys. 2011, 44, 472001.
- Munekata, P.E.S.; Domínguez, R.; Pateiro, M.; Lorenzo, J.M. Influence of Plasma Treatment on the Polyphenols of Food Products—A Review. Foods 2020, 9, 929.
- Graves, D.B.; Bakken, L.B.; Jensen, M.B.; Ingels, R. Plasma Activated Organic Fertilizer. Plasma Chem. Plasma Process. 2019, 39, 1–19.
- Zhang, S.; Rousseau, A.; Dufour, T. Promoting Lentil Germination and Stem Growth by Plasma Activated Tap Water, Demineralized Water and Liquid Fertilizer. RSC Adv. 2017, 7, 31244–31251.
- Misra, N.N.; Schlüter, O.; Cullen, P.J. Chapter 1—Plasma in Food and Agriculture. In Cold Plasma in Food and Agriculture; Misra, N.N., Schlüter, O., Cullen, P.J., Eds.; Academic Press: San Diego, CA, USA, 2016; pp. 1–16. ISBN 978-0-12-801365-6.
- Fan, L.; Liu, X.; Ma, Y.; Xiang, Q. Effects of Plasma-Activated Water Treatment on Seed Germination and Growth of Mung Bean Sprouts. J. Taibah Univ. Sci. 2020, 14, 823–830.
- Song, J.-S.; Lee, M.J.; Ra, J.E.; Lee, K.S.; Eom, S.; Ham, H.M.; Kim, H.Y.; Kim, S.B.; Lim, J. Growth and Bioactive Phytochemicals in Barley (Hordeum vulgare L.) Sprouts Affected by Atmospheric Pressure Plasma during Seed Germination. J. Phys. Appl. Phys. 2020, 53, 314002.
- Arjunan, K.P.; Sharma, V.K.; Ptasinska, S. Effects of Atmospheric Pressure Plasmas on Isolated and Cellular DNA—A Review. Int. J. Mol. Sci. 2015, 16, 2971–3016.
- Halliwell, B. Reactive Species and Antioxidants. Redox Biology Is a Fundamental Theme of Aerobic Life. Plant Physiol. 2006, 141, 312–322.
- Mhamdi, A.; Van Breusegem, F. Reactive Oxygen Species in Plant Development. Development 2018, 145, dev164376.
- El-Maarouf-Bouteau, H.; Bailly, C. Oxidative Signaling in Seed Germination and Dormancy. Plant Signal. Behav. 2008, 3, 175–182.
- Sarinont, T.; Amano, T.; Attri, P.; Koga, K.; Hayashi, N.; Shiratani, M.; Biophys, L.A.B. Effects of Plasma Irradiation Using Various Feeding Gases on Growth of Raphanus Sativus. Arch. Biochem. Biophys. 2016, 605, 129–140.
- Waskow, A.; Howling, A.; Furno, I. Mechanisms of Plasma-Seed Treatments as a Potential Seed Processing Technology. Front. Phys. 2021, 9, 174.
- Souza, F.H.; Marcos-Filho, J. The Seed Coat as a Modulator of Seed-Environment Relationships in Fabaceae. Braz. J. Bot. 2001, 24, 365–375.
- Yusupov, M.; Razzokov, J.; Cordeiro, R.M.; Bogaerts, A. Transport of Reactive Oxygen and Nitrogen Species across Aquaporin: A Molecular Level Picture. Oxidative Med. Cell. Longev. 2019, 2019, 1–11.
- Seol, Y.B.; Kim, J.; Park, S.H.; Chang, H.Y. Atmospheric Pressure Pulsed Plasma Induces Cell Death in Photosynthetic Organs via Intracellularly Generated ROS. Sci. Rep. 2017, 7, 1–11.
- Ka, D.H.; Priatama, R.A.; Park, J.Y.; Park, S.J.; Kim, S.B.; Lee, I.A.; Lee, Y.K. Plasma-Activated Water Modulates Root Hair Cell Density via Root Developmental Genes in Arabidopsis Thaliana L. Appl. Sci. 2021, 11, 2240.
- Lee, Y.K.; Lim, J.; Hong, E.J.; Kim, S.B. Plasma-Activated Water Regulates Root Hairs and Cotyledon Size Dependent on Cell Elongation in Nicotiana tabacum L. Plant Biotechnol. Rep. 2020, 14, 663–672.
- Park, D.P.; Davis, K.; Gilani, S.; Alonzo, C.-A.; Dobrynin, D.; Friedman, G.; Fridman, A.; Rabinovich, A.; Fridman, G. Reactive Nitrogen Species Produced in Water by Non-Equilibrium Plasma Increase Plant Growth Rate and Nutritional Yield. Curr. Appl. Phys. 2013, 13, S19–S29.
- Habibi, G. Hydrogen Peroxide (H2O2) Generation, Scavenging and Signaling in Plants. In Oxidative Damage to Plants; Ahmad, P., Ed.; Academic Press: San Diego, NJ, USA, 2014; pp. 557–584. ISBN 978-0-12-799963-0.
- Pei, Z.-M.; Murata, Y.; Benning, G.; Thomine, S.; Klüsener, B.; Allen, G.J.; Grill, E.; Schroeder, J.I. Calcium Channels Activated by Hydrogen Peroxide Mediate Abscisic Acid Signalling in Guard Cells. Nature 2000, 406, 731–734.
- Liu, Y.; Ye, N.; Liu, R.; Chen, M.; Zhang, J. H2O2 Mediates the Regulation of ABA Catabolism and GA Biosynthesis in Arabidopsis Seed Dormancy and Germination. J. Exp. Bot. 2010, 61, 2979–2990.
- Barba-Espín, G.; Diaz-Vivancos, P.; Job, D.; Belghazi, M.; Job, C.; Hernández, J.A. Understanding the Role of H2O2 during Pea Seed Germination: A Combined Proteomic and Hormone Profiling Approach. Plant Cell Environ. 2011, 34, 1907–1919.
- Kurek, K.; Plitta-Michalak, B.; Ratajczak, E. Reactive Oxygen Species as Potential Drivers of the Seed Aging Process. Plants 2019, 8, 174.
- Pandiselvam, R.; Mayookha, V.P.; Kothakota, A.; Sharmila, L.; Ramesh, S.V.; Bharathi, C.P. Impact of Ozone Treatment on Seed Germination-a Systematic Review. Ozone Sci. Eng. 2019, 42, 1–16.
- Palou, L.; Crisosto, C.H.; Smilanick, J.L.; Adaskaveg, J.E.; Zoffoli, J.P. Effects of Continuous 0.3 Ppm Ozone Exposure on Decay Development and Physiological Responses of Peaches and Table Grapes in Cold Storage. Postharvest Biol. Technol. 2002, 24, 39–48.
- Vlassi, E.; Vlachos, P.; Kornaros, M. Effect of Ozonation on Table Grapes Preservation in Cold Storage. J. Food Sci. Technol. 2018, 55, 2031–2038.
- Misra, N.N.; Moiseev, T.; Patil, S.; Pankaj, S.K.; Bourke, P.; Mosnier, J.P.; Keener, K.M.; Cullen, P.J. Cold Plasma in Modified Atmospheres for Post-Harvest Treatment of Strawberries. Food Bioprocess Technol. 2014, 7, 3045–3054.
- Waskow, A.; Howling, A.; Furno, I. Advantages and Limitations of Surface Analysis Techniques on Plasma-Treated Arabidopsis Thaliana Seeds. Front. Mater. 2021, 8, 123.
- Ölmez, H.; Akbas, M.Y. Optimization of Ozone Treatment of Fresh-Cut Green Leaf Lettuce. J. Food Eng. 2009, 90, 487–494.
- Mitsugi, F.; Abiru, T.; Ikegami, T.; Ebihara, K.; Aoqui, S.-I.; Nagahama, K. Influence of Ozone Generated by Surface Barrier Discharge on Nematode and Plant Growth. IEEE Trans. Plasma Sci. 2016, 44, 3071–3076.
- Los, A.; Ziuzina, D.; Boehm, D.; Cullen, P.J.; Bourke, P. Investigation of Mechanisms Involved in Germination Enhancement of Wheat (Triticum Aestivum) by Cold Plasma: Effects on Seed Surface Chemistry and Characteristics. Plasma Proc. Polym. 2019, 16, 1800148.
- Hasan, M.; Sohan, M.S.R.; Sajib, S.A.; Hossain, M.F.; Miah, M.; Maruf, M.M.H.; Khalid-Bin-Ferdaus, K.M.; Kabir, A.H.; Talukder, M.R.; Rashid, M.M.; et al. The Effect of Low-Pressure Dielectric Barrier Discharge (LPDBD) Plasma in Boosting Germination, Growth, and Nutritional Properties in Wheat. Plasma Chem. Plasma Process. 2022, 42, 339–362.
- Lindsay, A.; Byrns, B.; King, W.; Andhvarapou, A.; Fields, J.; Knappe, D.; Fonteno, W.; Shannon, S. Fertilization of Radishes, Tomatoes, and Marigolds Using a Large-Volume Atmospheric Glow Discharge. Plasma Chem. Plasma Process. 2014, 34, 1271–1290.
- Takahata, J.; Takaki, K.; Satta, N.; Takahashi, K.; Fujio, T.; Sasaki, Y. Improvement of Growth Rate of Plants by Bubble Discharge in Water. Jpn. J. Appl. Phys. 2014, 54, 01AG07.
- Takano, K.; Takahata, J.; Takaki, K.; Satta, N.; Takahashi, K.; Fujio, T. Improvement of Growth Rate of Brassica Para Var. Perviridis by Discharge inside Bubble under Water in Hydroponic Cultivation. Electron. Commun. Jpn. 2016, 99, 72–79.
- Kang, M.H.; Jeon, S.S.; Shin, S.M.; Veerana, M.; Ji, S.-H.; Uhm, H.-S.; Choi, E.-H.; Shin, J.H.; Park, G. Dynamics of Nitric Oxide Level in Liquids Treated with Microwave Plasma-Generated Gas and Their Effects on Spinach Development. Sci. Rep. 2019, 9, 1011.
- Lamichhane, P.; Veerana, M.; Lim, J.S.; Mumtaz, S.; Shrestha, B.; Kaushik, N.K.; Park, G.; Choi, E.H. Low-Temperature Plasma-Assisted Nitrogen Fixation for Corn Plant Growth and Development. Int. J. Mol. Sci. 2021, 22, 5360.
- Kidd, P.S.; Proctor, J. Why Plants Grow Poorly on Very Acid Soils: Are Ecologists Missing the Obvious? J. Exp. Bot. 2001, 52, 791–799.
- Sakudo, A.; Yagyu, Y.; Onodera, T. Disinfection and Sterilization Using Plasma Technology: Fundamentals and Future Perspectives for Biological Applications. Int. J. Mol. Sci. 2019, 20, 5216.
- Shintani, H.; Sakudo, A.; Burke, P.; McDonnell, G. Gas Plasma Sterilization of Microorganisms and Mechanisms of Action. Exp. Ther. Med. 2010, 1, 731–738.
- Laroussi, M. Cold Plasma in Medicine and Healthcare: The New Frontier in Low Temperature Plasma Applications. Front. Phys. 2020, 8, 74.
- Guo, J.; Wang, J.; Xie, H.; Jiang, J.; Li, C.; Li, W.; Li, L.; Liu, X.; Lin, F. Inactivation Effects of Plasma-Activated Water on Fusarium Graminearum. Food Control 2022, 134, 108683.
- Khamsen, N.; Onwimol, D.; Teerakawanich, N.; Dechanupaprittha, S.; Kanokbannakorn, W.; Hongesombut, K. Rice (Oryza sativa L.) Seed Sterilization and Germination Enhancement via Atmospheric Hybrid Nonthermal Discharge Plasma. ACS Appl. Mater. Interfaces 2016, 8, 19268–19275.
- Kang, M.H.; Pengkit, A.; Choi, K.; Jeon, S.S.; Choi, H.W.; Shin, D.B.; Choi, E.H.; Uhm, H.S.; Park, G. Differential Inactivation of Fungal Spores in Water and on Seeds by Ozone and Arc Discharge Plasma. PLoS ONE 2015, 10, e0139263.
- Świecimska, M.; Tulik, M.; Šerá, B.; Golińska, P.; Tomeková, J.; Medvecká, V.; Bujdáková, H.; Oszako, T.; Zahoranová, A.; Šerý, M. Non-Thermal Plasma Can Be Used in Disinfection of Scots Pine (Pinus sylvestris L.) Seeds Infected with Fusarium Oxysporum. Forests 2020, 11, 837.
- ŠERÁ, B.; ŠERÝ, M. Non-Thermal Plasma Treatment as a New Biotechnology in Relation to Seeds, Dry Fruits, and Grains. Plasma Sci. Technol. 2018, 20, 044012.
- Zahoranová, A.; Henselová, M.; Hudecová, D.; Kaliňáková, B.; Kováčik, D.; Medvecká, V.; Černák, M. Effect of Cold Atmospheric Pressure Plasma on the Wheat Seedlings Vigor and on the Inactivation of Microorganisms on the Seeds Surface. Plasma Chem. Plasma Process. 2016, 36, 397–414.
- Masiello, M.; Somma, S.; Lo Porto, C.; Palumbo, F.; Favia, P.; Fracassi, F.; Logrieco, A.F.; Moretti, A. Plasma Technology Increases the Efficacy of Prothioconazole against Fusarium graminearum and Fusarium proliferatum Contamination of Maize (Zea mays) Seedlings. Int. J. Mol. Sci. 2021, 22, 9301.
- Mravlje, J.; Regvar, M.; Vogel-Mikuš, K. Development of Cold Plasma Technologies for Surface Decontamination of Seed Fungal Pathogens: Present Status and Perspectives. J. Fungi 2021, 7, 650.
- Cold Plasma in Food and Agriculture: Fundamentals and Applications; Misra, N.N.; Schlüter, O.F.-K.; Cullen, P.J. (Eds.) Elsevier: Amsterdam, The Netherlands, 2016; ISBN 978-0-12-801365-6.
- Li, H.; Zhou, X.; Huang, Y.; Liao, B.; Cheng, L.; Ren, B. Reactive Oxygen Species in Pathogen Clearance: The Killing Mechanisms, the Adaption Response, and the Side Effects. Front. Microbiol. 2021, 11, 3610.
- Vaishampayan, A.; Grohmann, E. Antimicrobials Functioning through ROS-Mediated Mechanisms: Current Insights. Microorganisms 2021, 10, 61.
- Shaw, P.; Kumar, N.; Kwak, H.S.; Park, J.H.; Uhm, H.S.; Bogaerts, A.; Choi, E.H.; Attri, P. Bacterial Inactivation by Plasma Treated Water Enhanced by Reactive Nitrogen Species. Sci. Rep. 2018, 8, 11268.
- Moisan, M.; Barbeau, J.; Moreau, S.; Pelletier, J.; Tabrizian, M.; Yahia, L. Low-Temperature Sterilization Using Gas Plasmas: A Review of the Experiments and an Analysis of the Inactivation Mechanisms. Int. J. Pharm. 2001, 226, 1–21.
- Laroussi, M.; Leipold, F. Evaluation of the Roles of Reactive Species, Heat, and UV Radiation in the Inactivation of Bacterial Cells by Air Plasmas at Atmospheric Pressure. Int. J. Mass Spectrom. 2004, 233, 81–86.
- Lackmann, J.-W.; Schneider, S.; Edengeiser, E.; Jarzina, F.; Brinckmann, S.; Steinborn, E.; Havenith, M.; Benedikt, J.; Bandow, J.E. Photons and Particles Emitted from Cold Atmospheric-Pressure Plasma Inactivate Bacteria and Biomolecules Independently and Synergistically. J. R. Soc. Interface 2013, 10, 20130591.
- Chen, Z.; Garcia, G.; Arumugaswami, V.; Wirz, R.E. Cold Atmospheric Plasma for SARS-CoV-2 Inactivation. Phys. Fluids 2020, 32, 111702.
- Cortázar, O.D.; Megía-Macías, A.; Moreno, S.; Brun, A.; Gómez-Casado, E. Vulnerability of SARS-CoV-2 and PR8 H1N1 Virus to Cold Atmospheric Plasma Activated Media. Sci. Rep. 2022, 12, 263.
- Mohamed, H.; Nayak, G.; Rendine, N.; Wigdahl, B.; Krebs, F.C.; Bruggeman, P.J.; Miller, V. Non-Thermal Plasma as a Novel Strategy for Treating or Preventing Viral Infection and Associated Disease. Front. Phys. 2021, 9, 286.
- Filipić, A.; Primc, G.; Zaplotnik, R.; Mehle, N.; Gutierrez-Aguirre, I.; Ravnikar, M.; Mozetič, M.; Žel, J.; Dobnik, D. Cold Atmospheric Plasma as a Novel Method for Inactivation of Potato Virus Y in Water Samples. Food Environ. Virol. 2019, 11, 220–228.
- Filipić, A.; Dobnik, D.; Tušek Žnidarič, M.; Žegura, B.; Štern, A.; Primc, G.; Mozetič, M.; Ravnikar, M.; Žel, J.; Gutierrez Aguirre, I. Inactivation of Pepper Mild Mottle Virus in Water by Cold Atmospheric Plasma. Front. Microbiol. 2021, 12, 14.
- Filipić, A.; Gutierrez-Aguirre, I.; Primc, G.; Mozetič, M.; Dobnik, D. Cold Plasma, a New Hope in the Field of Virus Inactivation. Trends Biotechnol. 2020, 38, 1278–1291.
- Kaushik, N.; Mitra, S.; Baek, E.J.; Nguyen, L.N.; Bhartiya, P.; Kim, J.H.; Choi, E.H.; Kaushik, N.K. The Inactivation and Destruction of Viruses by Reactive Oxygen Species Generated through Physical and Cold Atmospheric Plasma Techniques: Current Status and Perspectives. J. Adv. Res. 2022.
- Dauwe, R.; Roulard, R.; Ramos, M.; Thiombiano, B.; Mesnard, F.; Gontier, E.; Jamali, A. Etching of the Seed Cuticle by Cold Plasma Shortens Imbibitional Leakage in Linum usitatissimum L. Ind. Crops Prod. 2021, 167, 113536.
- Kitazaki, S.; Sarinont, T.; Koga, K.; Hayashi, N.; Shiratani, M. Plasma Induced Long-Term Growth Enhancement of Raphanus sativus L. Using Combinatorial Atmospheric Air Dielectric Barrier Discharge Plasmas. Curr. Appl. Phys. 2014, 14, S149–S153.
- Mildažiene, V.; Pauzaite, G.; Malakauskiene, A.; Zukiene, R.; Nauciene, Z.; Filatova, I. Response of Perennial Woody Plants to Seed Treatment by Electromagnetic Field and Low-Temperature Plasma. Bioelectromagnetics 2016, 37, 536–548.
- Sera, B.; Spatenka, P.; Sery, M.; Vrchotova, N.; Hruskova, I. Influence of Plasma Treatment on Wheat and Oat Germination and Early Growth. IEEE Trans. Plasma Sci. 2010, 38, 2963–2968.
- Bormashenko, E.; Grynyov, R.; Bormashenko, Y.; Drori, E. Cold Radiofrequency Plasma Treatment Modifies Wettability and Germination Speed of Plant Seeds. Sci. Rep. 2012, 2, 741.
- Gao, X.; Zhang, A.; Héroux, P.; Sand, W.; Sun, Z.; Zhan, J. Effect of dielectric barrier discharge cold plasma on pea seed growth. J. Agric. Food Chem. 2019, 67, 10813–10822.
- Wang, X.Q.; Zhou, R.W.; De Groot, G.; Bazaka, K.; Murphy, A.B.; Ostrikov, K.K. Spectral Characteristics of Cotton Seeds Treated by a Dielectric Barrier Discharge Plasma. Sci. Rep. 2017, 7, 1–9.
- Da Silva, A.R.M.; Farias, M.L.; da Silva, D.L.S.; Vitoriano, J.O.; de Sousa, R.C.; Alves-Junior, C. Using Atmospheric Plasma to Increase Wettability, Imbibition and Germination of Physically Dormant Seeds of Mimosa Caesalpiniafolia. Coll. Surf. Biointerfaces 2017, 157, 280–285.
- Junior, C.A.; De Oliveira Vitoriano, J.; Da Silva, D.L.S.; De Lima Farias, M.; De Lima Dantas, N.B. Water Uptake Mechanism and Germination of Erythrina Velutina Seeds Treated with Atmospheric Plasma. Sci. Rep. 2016, 6, 1–7.
- Terebun, P.; Kwiatkowski, M.; Starek, A.; Reuter, S.; Mok, Y.S.; Pawłat, J. Impact of Short Time Atmospheric Plasma Treatment on Onion Seeds. Plasma Chem. Plasma Process. 2021, 41, 559–571.
- Sehrawat, R.; Thakur, A.K.; Vikram, A.; Vaid, A.; Rane, R. Effect of Cold Plasma Treatment on Physiological Quality of Okra Seed. J. Hill Agric. 2017, 8, 66–71.
- Shapira, Y.; Multanen, V.; Whyman, G.; Bormashenko, Y.; Chaniel, G.; Barkay, Z.; Bormashenko, E. Plasma Treatment Switches the Regime of Wetting and Floating of Pepper Seeds. Coll. Surf. Biointerfaces 2017, 157, 417–423.
- Molina, R.; Lalueza, A.; López-Santos, C.; Ghobeira, R.; Cools, P.; Morent, R.; de Geyter, N.; González-Elipe, A.R. Physicochemical Surface Analysis and Germination at Different Irrigation Conditions of DBD Plasma-Treated Wheat Seeds. Plasma Process. Polym. 2021, 18, 2000086.
- Zahoranová, A.; Hoppanová, L.; Šimončicová, J.; Tučeková, Z.; Medvecká, V.; Hudecová, D.; Kaliňáková, B.; Kováčik, D.; Černák, M. Effect of Cold Atmospheric Pressure Plasma on Maize Seeds: Enhancement of Seedlings Growth and Surface Microorganisms Inactivation. Plasma Chem. Plasma Process. 2018, 38, 969–988.
- Al-Whaibi, M.H. Plant heat-shock proteins: A mini review. J. King Saud. Univ. Sci. 2011, 23, 139–150.
- Waters, E.R.; Vierling, E. Plant Small Heat Shock Proteins—Evolutionary and Functional Diversity. New Phytol. 2020, 227, 24–37.
- Bourgine, B.; Guihur, A. Heat Shock Signaling in Land Plants: From Plasma Membrane Sensing to the Transcription of Small Heat Shock Proteins. Front. Plant Sci. 2021, 12, 1583.
- Holubová, Ľ.; Švubová, R.; Slováková, Ľ.; Bokor, B.; Chobotová Kročková, V.; Renčko, J.; Uhrin, F.; Medvecká, V.; Zahoranová, A.; Gálová, E. Cold Atmospheric Pressure Plasma Treatment of Maize Grains—Induction of Growth, Enzyme Activities and Heat Shock Proteins. Int. J. Mol. Sci. 2021, 22, 8509.
- Moisan, M.; Barbeau, J.; Crevier, M.-C.; Pelletier, J.; Philip, N.; Saoudi, B. Plasma Sterilization. Methods and Mechanisms. Pure Appl. Chem. 2002, 74, 349–358.
- Ibrahim, M.; Imram, M.; Hussain, A.; Aslam, M.; Rehmani, F.S.; Ali, B. Phytochemical studies on Amberboa ramose. Pak. J. Chem. 2012, 2, 24–28.
- Farokh, P.; Mahmoodzadeh, H.; Satari, T.N. Response of seed germination of safflower to UV-B radiation. Res. J. Env. Sci. 2010, 4, 70–74.
- Noble, R.E. Effects of UV-irradiation on seed germination. Sci. Total Env. 2002, 299, 173–176.
- Sadeghianfar, P.; Nazari, M.; Backes, G. Exposure to Ultraviolet (UV-C) radiation increases germination rate of maize (Zea maize L.) and sugar beet (Beta vulgaris) seeds. Plants 2019, 8, 49.
- Lotfy, K.; Al-Harbi, N.A.; Abd El-Raheem, H. Cold Atmospheric Pressure Nitrogen Plasma Jet for Enhancement Germination of Wheat Seeds. Plasma Chem. Plasma Process. 2019, 39, 897–912.
- Caldwell, M.M.; Björn, L.O.; Bornman, J.F.; Flint, S.D.; Kulandaivelu, G.; Teramura, A.H. Effects of Increased Solar Ultraviolet Radiation on Terrestrial Ecosystems. J. Photochem. Photobiol. Biol. 1998, 46, 40–52.
More
Information
Subjects:
Biology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
7.3K
Revisions:
3 times
(View History)
Update Date:
27 May 2022
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No