Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Jean-Marc Sabatier | -- | 2226 | 2022-05-10 13:01:25 | | | |
| 2 | Dean Liu | Meta information modification | 2226 | 2022-05-11 03:30:11 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Sabatier, J.; Fajloun, Z.; Khazaal, S.; Harb, J.; Rima, M.; Annweiler, C.; Wu, Y.; Cao, Z.; Abi Khattar, Z.; Legros, C.; et al. Long COVID in the Nervous System. Encyclopedia. Available online: https://encyclopedia.pub/entry/22769 (accessed on 28 July 2026).
Sabatier J, Fajloun Z, Khazaal S, Harb J, Rima M, Annweiler C, et al. Long COVID in the Nervous System. Encyclopedia. Available at: https://encyclopedia.pub/entry/22769. Accessed July 28, 2026.
Sabatier, Jean-Marc, Ziad Fajloun, Shaymaa Khazaal, Julien Harb, Mohamad Rima, Cedric Annweiler, Yingliang Wu, Zhijian Cao, Ziad Abi Khattar, Christian Legros, et al. "Long COVID in the Nervous System" Encyclopedia, https://encyclopedia.pub/entry/22769 (accessed July 28, 2026).
Sabatier, J., Fajloun, Z., Khazaal, S., Harb, J., Rima, M., Annweiler, C., Wu, Y., Cao, Z., Abi Khattar, Z., Legros, C., & De Neurophysiopathologie, I. (2022, May 10). Long COVID in the Nervous System. In Encyclopedia. https://encyclopedia.pub/entry/22769
Sabatier, Jean-Marc, et al. "Long COVID in the Nervous System." Encyclopedia. Web. 10 May, 2022.
Copy Citation
Long COVID is a term coined by the World Health Organization (WHO) to describe a variety of persistent symptoms after acute SARS-CoV-2 infection. Long COVID has been demonstrated to affect various SARS-CoV-2-infected persons, independently of the acute disease severity.
SARS-CoV-2
COVID-19
Long COVID
Renin-Angiotensin System
1. The Central Nervous System
The persisting sequelae after SARS-CoV-2 infection involving the central nervous system (CNS) included predominant neurologic and psychiatric symptoms, such as memory and attention deficits, reduced capacity to do daily tasks, frequent headaches, changes in cutaneous sensation, autonomic dysfunction, chronic fatigue, and, in severe cases, delusions and paranoia [1][2][3]. Surprisingly, many patients showing these neurologic symptoms for more than a year after contracting COVID-19 are under 50 years old and were healthy and active before getting infected [4]. This supports the idea that the virus can have a serious impact on the CNS. In fact, imaging results of the UK Biobank cohort indicated specific areas of brain atrophy in COVID-19 patients, compared to a control group without COVID-19, highlighting a direct impact of the virus on the CNS [4]. Also, neuronal atrophy and degeneration of cranial nerves, including the olfactory nerve and the neighboring olfactory bulb, have previously been reported in patients with persistent hyposmia or anosmia after acute COVID-19 [5]. In agreement with these findings, patients with long COVID exhibited lower metabolic activity in their brain [4]. The etiology underlying these symptoms and alterations in the brain, however, remains unknown. Of note, affected zones, including the cerebral cortex and hippocampus, possess different members of the RAS system.
2. The Peripheral Nervous System
The peripheral nervous system (PNS) can be also affected by SARS-CoV-2 infection and show persistent symptoms for weeks or even months after the infection. Peripheral neuropathy symptoms differ depending on which nerves are damaged: motor, sensory, or autonomic. COVID-19 infection is thought to affect the autonomic nervous system (ANS). In fact, the well-known COVID-19 cytokine response storm is caused by sympathetic activity which causes pro-inflammatory cytokine release. Vagus nerve stimulation mediating anti-inflammatory responses highlight the relationship between SARS-CoV-2 infection and the PNS and suggest the ANS as a therapeutic target. Of note, vagus nerve dysfunction has also been reported in SARS-CoV-2 infection and proposed as a key pathophysiological hallmark of long COVID [6].
COVID-19-related autonomic dysfunction might be caused by the virus itself [7]. In fact, cohort studies have identified viral infections as a common precursor to autonomic diseases such as orthostatic hypotension and postural orthostatic tachycardia syndrome. These immune-mediated neurological disorders are caused by autoantibodies, such as those against α/β adrenoceptors and muscarinic receptors [8][9][10][11][12]. As a result, it could be possible that SARS-CoV-2 infection promotes autonomic dysfunctions through an autoimmune-mediated process. In agreement with this hypothesis, Guillain-Barré syndrome (GBS), a rare, autoimmune disorder that targets nerves, was associated with SARS-CoV-2 infection [13][14]. These effects are described in macrophage activation syndrome induced by RAS dysfunction, which is known to cause an alteration of the innate immunity responsible for autoimmune diseases.
Symptoms of fever, cough, interstitial pneumonia, hypo-ageusia, and hypo-anosmia were associated with GBS in conjunction with COVID-19 [15]. This hypothesis was also investigated by checking whether some symptoms of long COVID could be linked to a virus- or immune-mediated disturbance of the ANS, resulting in acute or long-term orthostatic intolerance syndromes. Orthostatic hypotension (OH), vasovagal syncope (VVS), and postural orthostatic tachycardia syndrome (POTS) are all orthostatic intolerance syndromes [16]. An aberrant autonomic reaction to orthostasis is central to the pathogenesis (standing up). In fact, blood accumulates in the pelvis and legs when a healthy individual stands, decreasing venous return. Baroreceptors in the heart and aorta perceive this and respond by raising sympathetic neuronal and adrenergic tone (mediated by norepinephrine and epinephrine, respectively). This causes tachycardia followed by splanchnic vascular bed vasoconstriction, which enhances venous return to the heart [16]. The release of adrenaline and norepinephrine in orthostatic intolerance induces the significant tachycardia, which manifests as palpitations, dyspnea, and chest pain, which are typical symptoms of long COVID [17]. The poor control of heart rate variability (HRV) in long COVID patients also supports the autonomic dysfunction in these patients. This dysregulation was linked to long COVID symptoms such as fatigue and hypoxia [18]. Signs of peripheral nerve and muscular system dysfunction were also reported after SARS-CoV-2 infection. The persistent symptoms of myalgia, weakness (or exercise intolerance), sensory dysfunctions (primarily positive symptoms, such as paraesthesia and neuropathic pain), and dysautonomia were all linked to long COVID [2].
3. The Outcomes of Neuroinflammation
An important role of neuroinflammation has also been linked to long COVID. In fact, abnormal humoral and cellular immune responses, systemic inflammatory markers such as IL-6, and autoantibodies targeting cellular receptors could all be involved in systemic and neurological long COVID sequelae [15]. The full scope of long COVID neurological problems has yet to be determined. Neuroinflammation and neuronal injury observed in acute COVID-19 raise the possibility that infection could speed up or induce the development of neurodegenerative illnesses like Alzheimer’s or Parkinson’s. There is currently no evidence on the neurodevelopmental trajectories of children who typically have modest COVID-19 and show low neurologic or neuropsychiatric symptoms during or after acute illness. Because of broad endothelial activation, which commonly involves the brain, those who suffer from the unusual multisystem inflammatory syndrome in children (MIS-C) may be at a higher risk for neurological complications [4].
4. SARS-CoV-2 Entry into the CNS and PNS
Hematogenous or transsynaptic pathways involving the ACE2 receptor, which is found on the surface of a variety of cells, including neurons, astrocytes, endothelial, and smooth muscle cells of cerebral blood vessels, and skeletal muscle cells, facilitate SARS-CoV-2 cell entry into the CNS and PNS [19]. In the CNS, ACE2 receptors are predominantly expressed in the olfactory bulb, amygdala, hippocampus, middle temporal gyrus, posterior cingulate cortex, and the brainstem; therefore, hyposmia, mood disorders, cognitive impairment, sleep disorders, and dysautonomia have been linked to the dysfunction of these ‘ACE2-rich’ brain areas [1][18][20][21]. SARS-CoV-2 could indeed infect brain cells such as neurons, astrocytes, and microglial cells (and possibly oligodendrocytes) by binding to their ACE2 receptors and, as a result, after entering the ear-nose-throat (ENT) sphere, attacking the olfactory bulb on the floor of the cranial box through the olfactory epithelium just below the nasal cavity level. The involvement of the brainstem and cerebellum is critical evidence suggesting that these brain regions may be involved in several neurological manifestations of long COVID which are similar to myalgic encephalomyelitis or chronic fatigue syndrome (ME/CFS) and POTS [5].
5. The Complexity of RAS in the CNS and Its Impairing in Long COVID
Besides the endocrine RAS regulating water and electrolyte balance, it was demonstrated that neurons have intracrine and local types of RAS [22][23]. RAS regulates brain homeostasis primarily by the action of four angiotensin receptor subtypes: AT1R, AT2R, MasR, and angiotensin II type-4 receptor (AT4R). AT1R induces vasoconstriction, proliferation, inflammation, and oxidative stress, whereas AT2R and MasR mitigate AT1R’s actions. AT1Rs are divided into two types: AT1A and AT1B. The AT1A receptor is mostly found in brain regions that contribute to blood pressure and electrolyte balance homeostasis, whereas the AT1B receptor is found in structures like the cerebral cortex and hippocampus that are involved in memory and higher brain functions. AT4R regulates the release of dopamine and acetylcholine, as well as memory and learning consolidation [24][25]. AT2R is abundantly expressed in the brain. It plays a role in brain damage healing (i.e., axonal regeneration, neurite development) as well as inflammation reduction and vasodilation. In other words, AT2R has a neuroprotective function by helping neuronal survival and protecting the brain from damages. In this context, the AT2R protects against the harmful effects of AT1R activation, which occurs during SARS-CoV-2 infection [26][27]. It has been shown that SARS-CoV-2 has a brain tropism, and the neurological dysfunctions reported could be due to the impairment of RAS in the CNS (Figure 1). It has been shown that SARS-CoV-2 can infect nerve cells (such as neurons and astrocytes that express the ACE2 receptor) in vitro and in vivo [23][24][28][29][30][31]. The fact that SARS-CoV-2 impairs RAS pathways suggests that many nervous sequelae of long COVID could be explained by ACE2 receptor loss and AngII/AT1R over-activation. The virus binds to ACE2, resulting in its downregulation and a shift in the dynamic balance between the two faces of the RAS: (1) ACE/Ang II/AT1R, which has proinflammatory features, and (2) ACE2/Ang-(1-7)/AT2R, which has anti-inflammatory properties. This imbalance due to the overactivation of the Ang II/AT1R axis in the brain leads to hypertension, neuroinflammation, increased oxidative stress, BBB disruption, and neurotoxicity [20][24][25].
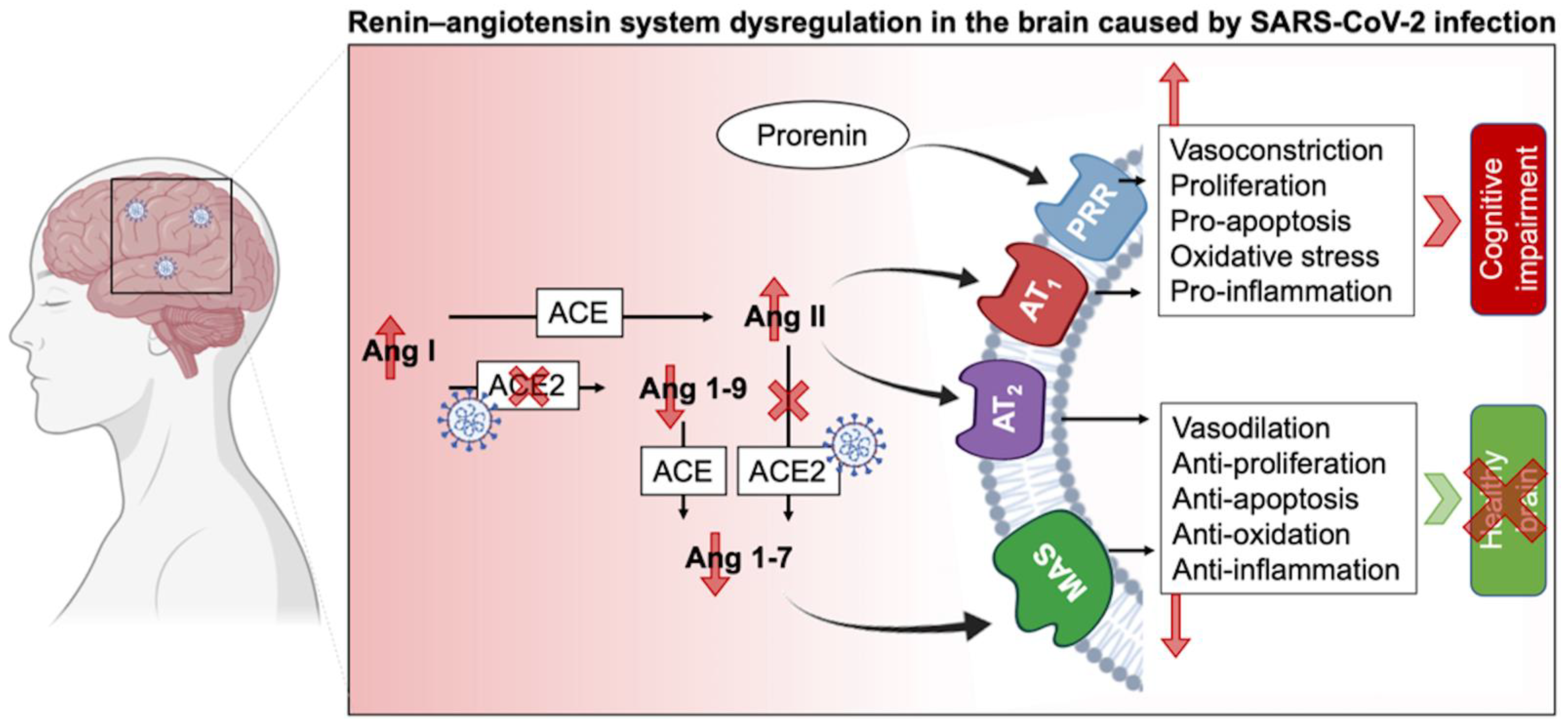
Figure 1. Schematic of RAS impairment in the brain duringSARS-CoV-2 infection. ACE: Angiotensin-converting enzyme; Ang: Angiotensin; AT1R: Angiotensin II receptor type 1; AT2R: Angiotensin II receptor type 2; MAS: Mas-related G protein-coupled receptors, PRR: Prorenin receptor.
The abundancy of RAS in the brain and its involvement in different physiological and cognitive processes explain the broad consequences of SARS-CoV-2 infection. The diversity of the RAS, notably via its two types, circulating and local RAS, emphasizes its important role in homeostasis. As such, circulating RAS affects nuclei in the hypothalamus and medulla via circumventricular organs, which are brain areas that lack a blood-brain barrier (BBB), and transmit to nuclei in the hypothalamus and medulla, while the independent local RAS of the brain, b-RAS, generates all components of the circulatory RAS [22]. On the other hand, the prorenin receptor (PRR) is widely expressed in neurons, and some microglial cells of many vascular brain regions as well as the brain cortex and basal ganglia [32]. Overstimulation of this system activates the Ang II/AT1R axis and thus may lead to cognitive impairment. Furthermore, PRR can establish its own signaling pathway and produce pro-oxidative effects. Although a link connecting PRR and neural development has been shown, the RAS-independent functions of PRR in the brain are yet unknown [1][33].
AT1R signaling can be also triggered by ACE overexpression [34] and results in (i) second messenger signaling including inositol trisphosphate, diacylglycerol, and arachidonic acid, as well as the (ii) activation of downstream effectors including phospholipases C, A, and D after G-protein coupling stimulation of AT1R by Ang II. The AT1R signaling cascade activates protein kinase C, Akt, intracellular protein kinases and serine/threonine kinases (such as mitogen-activated protein kinase (MAPK) family kinases). These facts highlight the diversity of the components that can be touched by RAS impairment and their consequent sequelae. For example, hypertrophy, vascular remodeling, and hyperplasia may arise from overactivation of the AT1R cascade [24][34][35]. The Ang II/AT2Rs counteract AT1R’s effects by promoting phospholipase A2 and activating multiple protein phosphatases as well as the nitric oxide (NO)/cyclic GMP pathway, triggering the release of arachidonic acid [3][27]. AT2R also suppresses cell growth and proliferation by blocking insulin and EGFR autophosphorylation. Furthermore, by inhibiting negative feedback, AT1R blockage promotes angiotensinogen and AT2R stimulation [3][27]. This balance between AT1R and AT2R is important for maintaining a normal physiological environment. Therefore, its perturbation, specifically via the AngII/AT1R connection, results in a slew of potentially harmful consequences on the endothelium, inflammation, and coagulation, in addition to the well-known vasoconstrictive effects in the brain that may potentiate the post-acute COVID symptoms. Since the interactions between AngII and AT2R and Ang II and MasR operate as a vital “protective arm” to balance these effects [36], ACE2 is considered as a key component of the anti-RAS system in the genesis and prevention of illness.
Positron emission tomography (PET) of long COVID patients, with persistent complaints at least three weeks after the onset of symptoms of acute infection, showed hypometabolism in their bilateral rectal/orbital gyrus (containing the olfactory gyrus), right temporal lobe (amygdala and hippocampus extending to the right thalamus), bilateral pons/medulla brainstem, and bilateral cerebellum. Interestingly, the metabolism of the frontal cluster (containing the olfactory gyrus) was worse in subjects receiving ACE inhibitors for high blood pressure, but better in subjects who received nasal decongestant spray [37]. These findings could point to the involvement of the ACE2 receptor in SARS-CoV-2 neurotropism, particularly in the olfactory bulb. This is likely due to the propagation pathway from the nose to the olfactory bulb, where the ACE2 receptor is strongly expressed, which has been hypothesized for various coronaviruses [38]. Cerebellar hypometabolism results in several symptoms including hyposmia/anosmia, memory/cognitive impairment, which is concordant with the involvement of this region in executive functions and working memory [39][40]. In addition, hypometabolism in the frontal cortex, brainstem, and cerebellum is associated with pain symptoms [41], particularly in fibromyalgia patients [42]. Other disorders have been linked to hypometabolism of the brainstem and cerebellum, such as insomnia and dysautonomia [43]. Taken together, the hypometabolism detected in brain regions of long COVID patients explain some of the persistent symptoms. The relationship between PET hypometabolism and the duration after the onset of the acute infection symptoms could suggest a link with the clinical severity. A more severe clinical profile is associated with a more severe PET hypometabolism and a longer duration of symptoms. Therefore, tracking functional brain activity can serve as a cerebral biomarker tool to classify long COVID patients regarding the severity of their symptoms and to distinguish affected patients from healthy subjects [37]. Also, these findings give insight to act quickly and restore brain metabolism in these patients before more severe diseases develop, such as Alzheimer’s disease (AD), which involves the severe reduction of the cerebral metabolic rate for glucose [44].
References
- Fajloun, Z.; Kovacic, H.; Annweiler, C.; Wu, Y.; Cao, Z.; Sabatier, J.-M. SARS-CoV-2-Induced Neurological Disorders in Symptomatic COVID-19 and Long COVID Patients: Key Role of Brain Renin-Angiotensin System. Infect. Disord.-Drug Targets 2022, 22.
- Dani, M.; Dirksen, A.; Taraborrelli, P.; Torocastro, M.; Panagopoulos, D.; Sutton, R.; Lim, P.B. Autonomic dysfunction in ‘long COVID’: Rationale, physiology and management strategies. Clin. Med. 2021, 21, e63–e67.
- Annweiler, C.; Bourgeais, A.; Faucon, E.; Cao, Z.; Wu, Y.; Sabatier, J.-M. Neurological, Cognitive, and Behavioral Disorders during COVID-19: The Nitric Oxide Track. J. Am. Geriatr. Soc. 2020, 68, 1922–1923.
- Spudich, S.; Nath, A. Nervous system consequences of COVID-19. Science 2022, 375, 267–269.
- Kandemirli, S.G.; Altundag, A.; Yildirim, D.; Tekcan Sanli, D.E.; Saatci, O. Olfactory Bulb MRI and Paranasal Sinus CT Findings in Persistent COVID-19 Anosmia. Acad. Radiol. 2021, 28, 28–35.
- Proal, A.D.; VanElzakker, M.B. Long COVID or Post-acute Sequelae of COVID-19 (PASC): An Overview of Biological Factors That May Contribute to Persistent Symptoms. Front. Microbiol. 2021, 12, 698169.
- Goldstein, D.S. The extended autonomic system, dyshomeostasis, and COVID-19. Clin. Auton. Res. 2020, 30, 299–315.
- Li, H.; Kem, D.C.; Reim, S.; Khan, M.; Vanderlinde-Wood, M.; Zillner, C.; Collier, D.; Liles, C.; Hill, M.A.; Cunningham, M.W.; et al. Agonistic Autoantibodies as Vasodilators in Orthostatic Hypotension. Hypertension 2012, 59, 402–408.
- Yu, X.; Stavrakis, S.; Hill, M.A.; Huang, S.; Reim, S.; Li, H.; Khan, M.; Hamlett, S.; Cunningham, M.W.; Kem, D.C. Autoantibody activation of beta-adrenergic and muscarinic receptors contributes to an “autoimmune” orthostatic hypotension. J. Am. Soc. Hypertens. 2012, 6, 40–47.
- Ruzieh, M.; Batizy, L.; Dasa, O.; Oostra, C.; Grubb, B. The role of autoantibodies in the syndromes of orthostatic intolerance: A systematic review. Scand. Cardiovasc. J. 2017, 51, 243–247.
- Li, H.; Yu, X.; Liles, C.; Khan, M.; Vanderlinde-Wood, M.; Galloway, A.; Zillner, C.; Benbrook, A.; Reim, S.; Collier, D.; et al. Autoimmune Basis for Postural Tachycardia Syndrome. J. Am. Heart Assoc. 2014, 3, e000755.
- Gunning, W.T.; Kvale, H.; Kramer, P.M.; Karabin, B.L.; Grubb, B.P. Postural Orthostatic Tachycardia Syndrome Is Associated With Elevated G-Protein Coupled Receptor Autoantibodies. J. Am. Heart Assoc. 2019, 8, e013602.
- Uncini, A.; Vallat, J.-M.; Jacobs, B.C. Guillain-Barré syndrome in SARS-CoV-2 infection: An instant systematic review of the first six months of pandemic. J. Neurol. Neurosurg. Psychiatry 2020, 91, 1105–1110.
- Khalifa, M.; Zakaria, F.; Ragab, Y.; Saad, A.; Bamaga, A.; Emad, Y.; Rasker, J.J. Guillain-Barré Syndrome Associated With Severe Acute Respiratory Syndrome Coronavirus 2 Detection and Coronavirus Disease 2019 in a Child. J. Pediatric Infect. Dis. Soc. 2020, 9, 510–513.
- Wallukat, G.; Hohberger, B.; Wenzel, K.; Fürst, J.; Schulze-Rothe, S.; Wallukat, A.; Hönicke, A.-S.; Müller, J. Functional autoantibodies against G-protein coupled receptors in patients with persistent Long-COVID-19 symptoms. J. Transl. Autoimmun. 2021, 4, 100100.
- Fedorowski, A. Postural orthostatic tachycardia syndrome: Clinical presentation, aetiology and management. J. Intern. Med. 2019, 285, 352–366.
- Port, J.D.; Gilbert, E.M.; Larrabee, P.; Mealey, P.; Volkman, K.; Ginsburg, R.; Hershberger, R.E.; Murray, J.; Bristow, M.R. Neurotransmitter depletion compromises the ability of indirect-acting amines to provide inotropic support in the failing human heart. Circulation 1990, 81, 929–938.
- Barizien, N.; Le Guen, M.; Russel, S.; Touche, P.; Huang, F.; Vallée, A. Clinical characterization of dysautonomia in long COVID-19 patients. Sci. Rep. 2021, 11, 14042.
- Wang, Q.; Zhang, Y.; Wu, L.; Niu, S.; Song, C.; Zhang, Z.; Lu, G.; Qiao, C.; Hu, Y.; Yuen, K.-Y.; et al. Structural and Functional Basis of SARS-CoV-2 Entry by Using Human ACE2. Cell 2020, 181, 894–904.e9.
- Eshak, N.; Abdelnabi, M.; Ball, S.; Elgwairi, E.; Creed, K.; Test, V.; Nugent, K. Dysautonomia: An Overlooked Neurological Manifestation in a Critically ill COVID-19 Patient. Am. J. Med. Sci. 2020, 360, 427–429.
- Balcom, E.F.; Nath, A.; Power, C. Acute and chronic neurological disorders in COVID-19: Potential mechanisms of disease. Brain 2021, 144, 3576–3588.
- Saavedra, J.M. Brain Angiotensin II: New Developments, Unanswered Questions and Therapeutic Opportunities. Cell. Mol. Neurobiol. 2005, 25, 485–512.
- El-Arif, G.; Khazaal, S.; Farhat, A.; Harb, J.; Annweiler, C.; Wu, Y.; Cao, Z.; Kovacic, H.; Abi Khattar, Z.; Fajloun, Z.; et al. Angiotensin II Type I Receptor (AT1R): The Gate towards COVID-19-Associated Diseases. Molecules 2022, 27, 2048.
- El-Arif, G.; Farhat, A.; Khazaal, S.; Annweiler, C.; Kovacic, H.; Wu, Y.; Cao, Z.; Fajloun, Z.; Khattar, Z.A.; Sabatier, J.M. The Renin-Angiotensin System: A Key Role in SARS-CoV-2-Induced COVID-19. Molecules 2021, 26, 6945.
- Grobe, J.L.; Xu, D.; Sigmund, C.D. An Intracellular Renin-Angiotensin System in Neurons: Fact, Hypothesis, or Fantasy. Physiology 2008, 23, 187–193.
- Annweiler, C.; Papon, N.; Sabatier, J.-M.; Barré, J. DAMPening Severe COVID-19 with Dexamethasone. Infect. Disord.-Drug Targets 2022, 22, 11–12.
- Abadir, P.M. The Frail Renin-Angiotensin System. Clin. Geriatr. Med. 2011, 27, 53–65.
- Kobori, H.; Nangaku, M.; Navar, L.G.; Nishiyama, A. The Intrarenal Renin-Angiotensin System: From Physiology to the Pathobiology of Hypertension and Kidney Disease. Pharmacol. Rev. 2007, 59, 251–287.
- Li, W.; Moore, M.J.; Vasilieva, N.; Sui, J.; Wong, S.K.; Berne, M.A.; Somasundaran, M.; Sullivan, J.L.; Luzuriaga, K.; Greenough, T.C.; et al. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 2003, 426, 450–454.
- Lyoo, K.-S.; Kim, H.M.; Lee, B.; Che, Y.H.; Kim, S.-J.; Song, D.; Hwang, W.; Lee, S.; Park, J.-H.; Na, W.; et al. Direct neuronal infection of SARS-CoV-2 reveals cellular and molecular pathology of chemosensory impairment of COVID-19 patients. Emerg. Microbes Infect. 2022, 11, 406–411.
- Rothan, H.A.; Kumari, P.; Stone, S.; Natekar, J.P.; Arora, K.; Auroni, T.T.; Kumar, M. SARS-CoV-2 Infects Primary Neurons from Human ACE2 Expressing Mice and Upregulates Genes Involved in the Inflammatory and Necroptotic Pathways. Pathogens 2022, 11, 257.
- Nguyen, G.; Delarue, F.; Burcklé, C.; Bouzhir, L.; Giller, T.; Sraer, J.-D. Pivotal role of the renin/prorenin receptor in angiotensin II production and cellular responses to renin. J. Clin. Investig. 2002, 109, 1417–1427.
- Nakagawa, P.; Gomez, J.; Grobe, J.L.; Sigmund, C.D. The Renin-Angiotensin System in the Central Nervous System and Its Role in Blood Pressure Regulation. Curr. Hypertens. Rep. 2020, 22, 7.
- Jackson, L.; Eldahshan, W.; Fagan, S.; Ergul, A. Within the Brain: The Renin Angiotensin System. Int. J. Mol. Sci. 2018, 19, 876.
- Cao, Z.; Wu, Y.; Faucon, E.; Sabatier, J.-M. SARS-CoV-2 & COVID-19: Key-Roles of the ‘Renin-Angiotensin’ System / Vitamin D Impacting Drug and Vaccine Developments. Infect. Disord.-Drug Targets 2020, 20, 348–349.
- Paz Ocaranza, M.; Riquelme, J.A.; García, L.; Jalil, J.E.; Chiong, M.; Santos, R.A.S.; Lavandero, S. Counter-regulatory renin–angiotensin system in cardiovascular disease. Nat. Rev. Cardiol. 2020, 17, 116–129.
- Guedj, E.; Campion, J.Y.; Dudouet, P.; Kaphan, E.; Bregeon, F.; Tissot-Dupont, H.; Guis, S.; Barthelemy, F.; Habert, P.; Ceccaldi, M.; et al. 18F-FDG brain PET hypometabolism in patients with long COVID. Eur. J. Nucl. Med. Mol. Imaging 2021, 48, 2823–2833.
- Baig, A.M.; Khaleeq, A.; Ali, U.; Syeda, H. Evidence of the COVID-19 Virus Targeting the CNS: Tissue Distribution, Host–Virus Interaction, and Proposed Neurotropic Mechanisms. ACS Chem. Neurosci. 2020, 11, 995–998.
- Marvel, C.L.; Morgan, O.P.; Kronemer, S.I. How the motor system integrates with working memory. Neurosci. Biobehav. Rev. 2019, 102, 184–194.
- Bodranghien, F.; Bastian, A.; Casali, C.; Hallett, M.; Louis, E.D.; Manto, M.; Mariën, P.; Nowak, D.A.; Schmahmann, J.D.; Serrao, M.; et al. Consensus Paper: Revisiting the Symptoms and Signs of Cerebellar Syndrome. The Cerebellum 2016, 15, 369–391.
- Borsook, D.; Sava, S.; Becerra, L. The Pain Imaging Revolution: Advancing Pain Into the 21st Century. Neuroscientist 2010, 16, 171–185.
- Guedj, E.; Cammilleri, S.; Niboyet, J.; Dupont, P.; Vidal, E.; Dropinski, J.-P.; Mundler, O. Clinical Correlate of Brain SPECT Perfusion Abnormalities in Fibromyalgia. J. Nucl. Med. 2008, 49, 1798–1803.
- Canto, C.B.; Onuki, Y.; Bruinsma, B.; van der Werf, Y.D.; De Zeeuw, C.I. The Sleeping Cerebellum. Trends Neurosci. 2017, 40, 309–323.
- Mosconi, L.; Pupi, A.; De Leon, M.J. Brain Glucose Hypometabolism and Oxidative Stress in Preclinical Alzheimer’s Disease. Ann. N. Y. Acad. Sci. 2008, 1147, 180–195.
More
Information
Subjects:
Infectious Diseases
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.4K
Revisions:
2 times
(View History)
Update Date:
11 May 2022
Table of Contents



Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No