 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Jasmin Knopf | -- | 3827 | 2022-04-21 12:24:09 | | | |
| 2 | Vivi Li | + 38 word(s) | 3865 | 2022-04-22 05:50:42 | | |
Video Upload Options
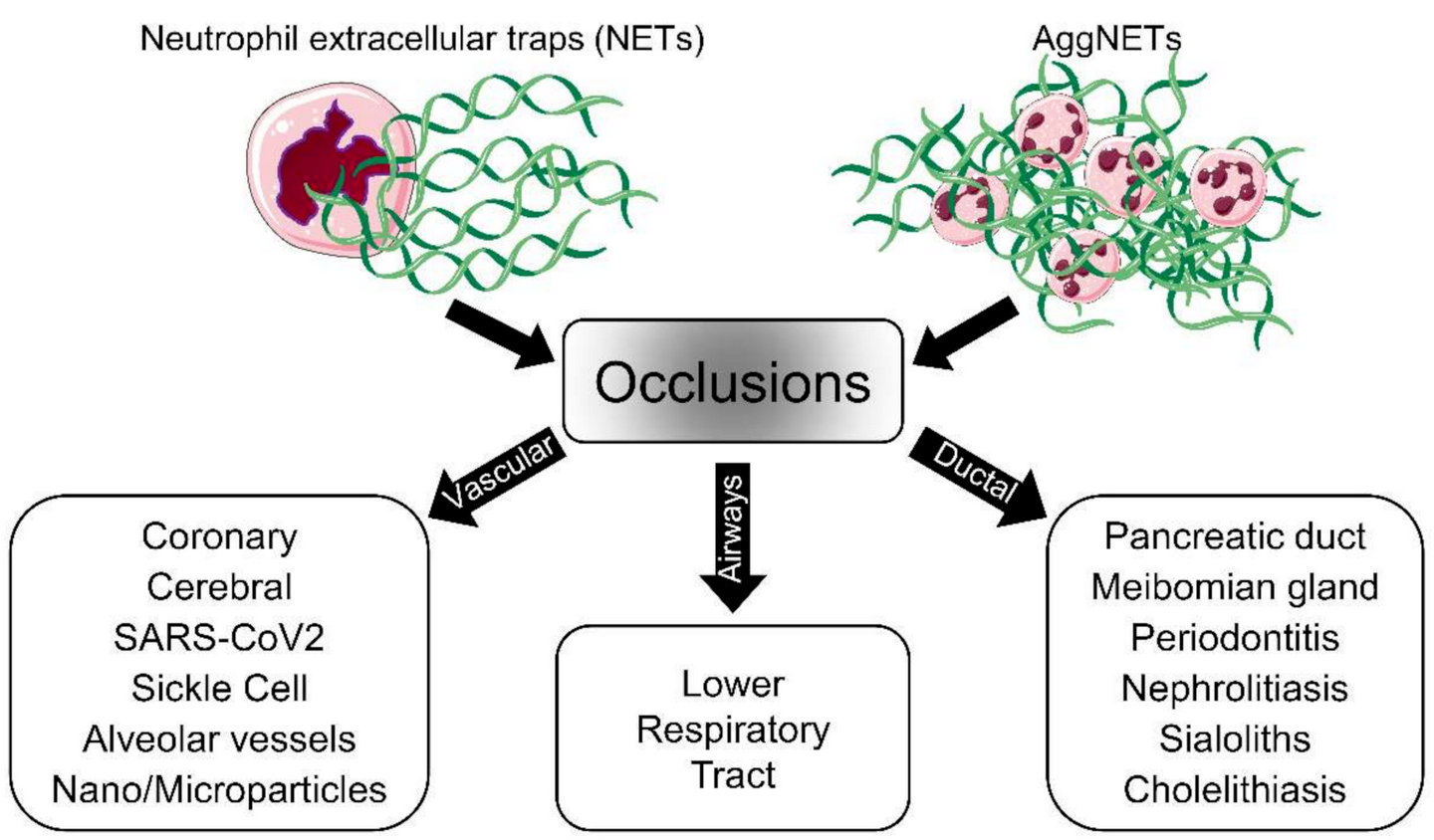
The discovery of neutrophil extracellular trap (NET) formation as a part of the defense mechanisms of the innate immune system has provided new insights into the pathologies of various diseases. Nowadays, NET formation is considered a double-edged sword, as NET remnants induce inflammation and aggregated NETs (aggNETs) reportedly occlude tubular structures like vessels or ducts. In this regard, elucidating the mechanism of NET-dependent occlusions is crucial for the development of new therapeutic approaches.
1. Introduction
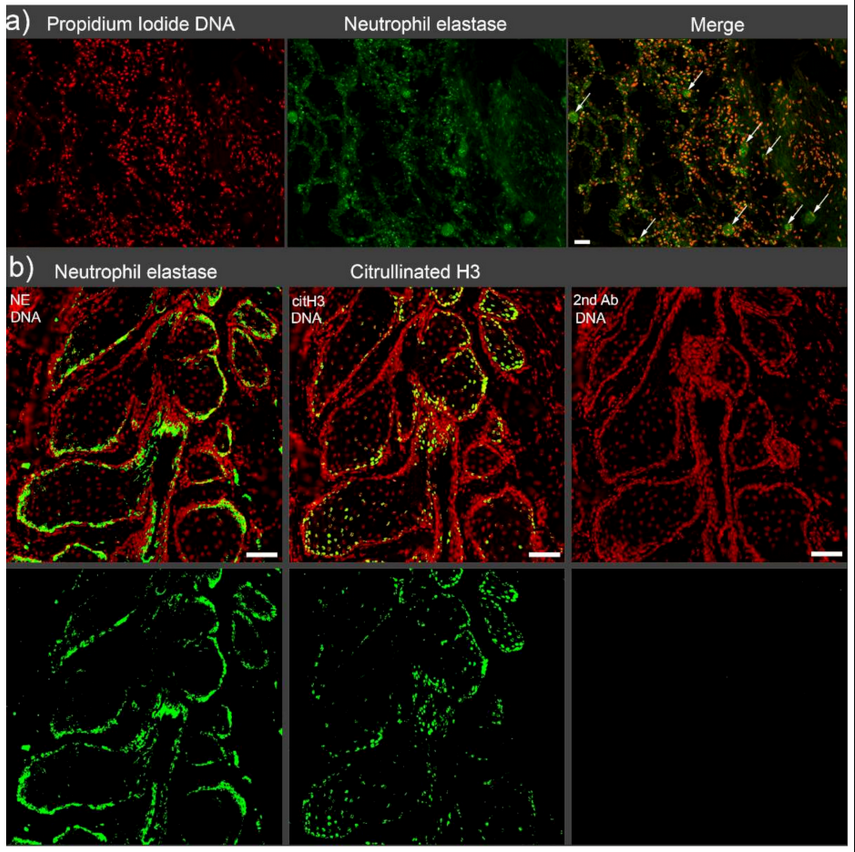
2. Vascular Occlusions
2.1. Vascular Occlusions in COVID-19
2.2. Coronary Occlusions
2.3. Cerebral Occlusions
2.4. Sickle Cell Disease
3. Airway Occlusions
4. Occlusions of Exocrine Glands and Ducts
4.1. Pancreatic Duct
The pancreatic duct, also known as the Wirsung duct, connects the pancreas and intestine through the common bile duct. Its primary function is to transport enzymes and bicarbonate, which aid digestion and neutralize the duodenal pH, respectively [64][65]. Occlusion of the pancreatic duct may cause pancreatitis. The occlusions of the ducts are directly proportional to the severity of the pancreatitis and depend on the duration of the disease [64][65].
NETs have been reported to directly induce trypsin activation, inflammation, and tissue damage in severe acute pancreatitis induced by retrograde taurocholate infusion [66]. However, the direct cause of obstruction is not always found in human acute pancreatitis. Under physiological conditions, neutrophils are present in small amounts in the pancreas and enter the bicarbonate-rich pancreatic fluid and spontaneously form NETs [67]. In the case of severe inflammatory situations, the resulting neutrophilia can produce excessive amounts of NETs in the pancreas. This results in large and sticky aggregates prone to occlude pancreatic ducts. Leppkes et al. reported that sole neutrophilia in IL17 transgenic mice was the main driving force for the development of pancreatitis. In this case, excessive NETs formation was immediately induced by a high bicarbonate concentration in the pancreatic juice. Formed NETs tend to aggregate and occlude the ducts triggering focal acute pancreatic [68]. The reduced production and sizes of NETs and aggNETs in the pancreatic ducts of PAD4-KO mice prevented the development of focal pancreatitis [69][70].
4.2. Meibomian Gland
4.3. Periodontitis (Periodontal Crevicular Occlusions)
The gingival crevice, also called the gingival sulcus, is defined as a narrow V-shaped space between the inner aspect of the free gingival epithelium and the surrounding enamel of a tooth. Normally, its depth range is 1–3 mm. The gingival epithelium continuously produces gingival crevicular fluid (GCF) that is finally transferred into the oral cavity. It has been known for many years that the production of GCF and its composition change during inflammatory diseases like periodontitis. Therefore, GCF has been extensively studied as a diagnostic tool [76][77]. Under physiological conditions, the minimal amount of GCF present in the gingival crevice flows into the oral cavity, making the crevice a kind of duct. Upon inflammation, this fluid transforms into a purulent exudate containing large amounts of NETs and neutrophils expressing CD177 [78][79]. The exudate is extremely viscous due to the excessive amount of aggNETs. Vitkov and colleagues put forward the view of NETs-driven obstruction of the periodontal crevice. The authors speculated that the formation of a periodontal abscess might result from NETs-induced cervical obstructions [6] supported by the overproduction of NETs and the impaired clearance of NETs remnants [80]. This hypothesis has not yet been proven experimentally. However, the crucial roles of NETs in the initiation and progression of inflammatory periodontal diseases are surely worth investigating.
4.4. Gallstones
Obstruction of the biliary system, a common, serious, and painful condition, is one of the leading causes of hospitalization with significant morbidity and mortality. The formation of gallstones in the gallbladder or ducts, also referred to as cholelithiasis, are the most prevalent etiological event for biliary obstruction that results in biliary stasis. However, gallstone-caused biliary obstructions generate a high socioeconomic burden due to their high incidence, especially in developed countries [81][82]. Until recently, it has been proposed that gallstones simply form due to the supersaturation of cholesterol crystals. The contents of human gallstones were eventually investigated for NETs, as cholelithiasis is an occlusive condition [83]. After observing extracellular DNA and neutrophil elastase in gall sludge and gallstones, it was established that an intact NETs formation capacity is necessary to form gallstones in a murine model of cholelithiasis. Basically, NETs were crucial in the initiation and progression of gallstone formation by promoting the aggregation of biliary cholesterol and calcium crystals. Furthermore, a positive correlation between the neutrophil/lymphocyte ratio and the severity of gallstone-induced pancreatitis has been reported in clinical settings [84]. However, formal clinical trials employing NETs formation inhibition in patients with recurrent cholelithiasis are still needed [85].
4.5. Sialoliths
Sialoliths, also known as salivary stones, are the most prevalent obstructive disease of the salivary glands, especially for middle-aged patients. Stone formation is mostly seen in the submandibular gland, with an incidence of more than 80 percent. The parotid gland follows it with 13 percent and by sublingual and minor salivary glands at very low rates. The etiology of sialolith formation was initially considered a multifactorial interaction of calcium salts, organic and inorganic molecules, pH, and bacteria [86][87]. In two conflicting reports, inorganic materials [88] and organic components [89] have claimed to be the main components of sialoliths. However, since the role of inflammation in sialoliths formation [90] and the presence of bacterial residues, including bacterial DNA and biofilms in their structures, has been established [91], NETs have gained attention as a possible etiological factor. Furthermore, since bacterial biofilm structures at the core of sialoliths have been established, biofilms have been hypothesized to be an initial step in the formation of sialoliths [92]. Recently, neutrophil recruitment and NETs formation in the salivary system were suggested to initiate sialolithiasis. Indeed, the demonstration of NE activity associated with a high prevalence of extracellular DNA points to NETs as nidi for the formation of sialoliths [93]. The high content of bicarbonate in saliva strengthens this hypothesis as it facilitates the formation of NETs [67]. The interaction between neutrophils and the precipitated particulate matter is discussed to cause the salivary stone to grow. These results offer an alternative perspective for the “until now” proposed mechanisms of sialolithogenesis [94][95].
5. Therapeutic Approaches to Prevent NETs-Driven Occlusions
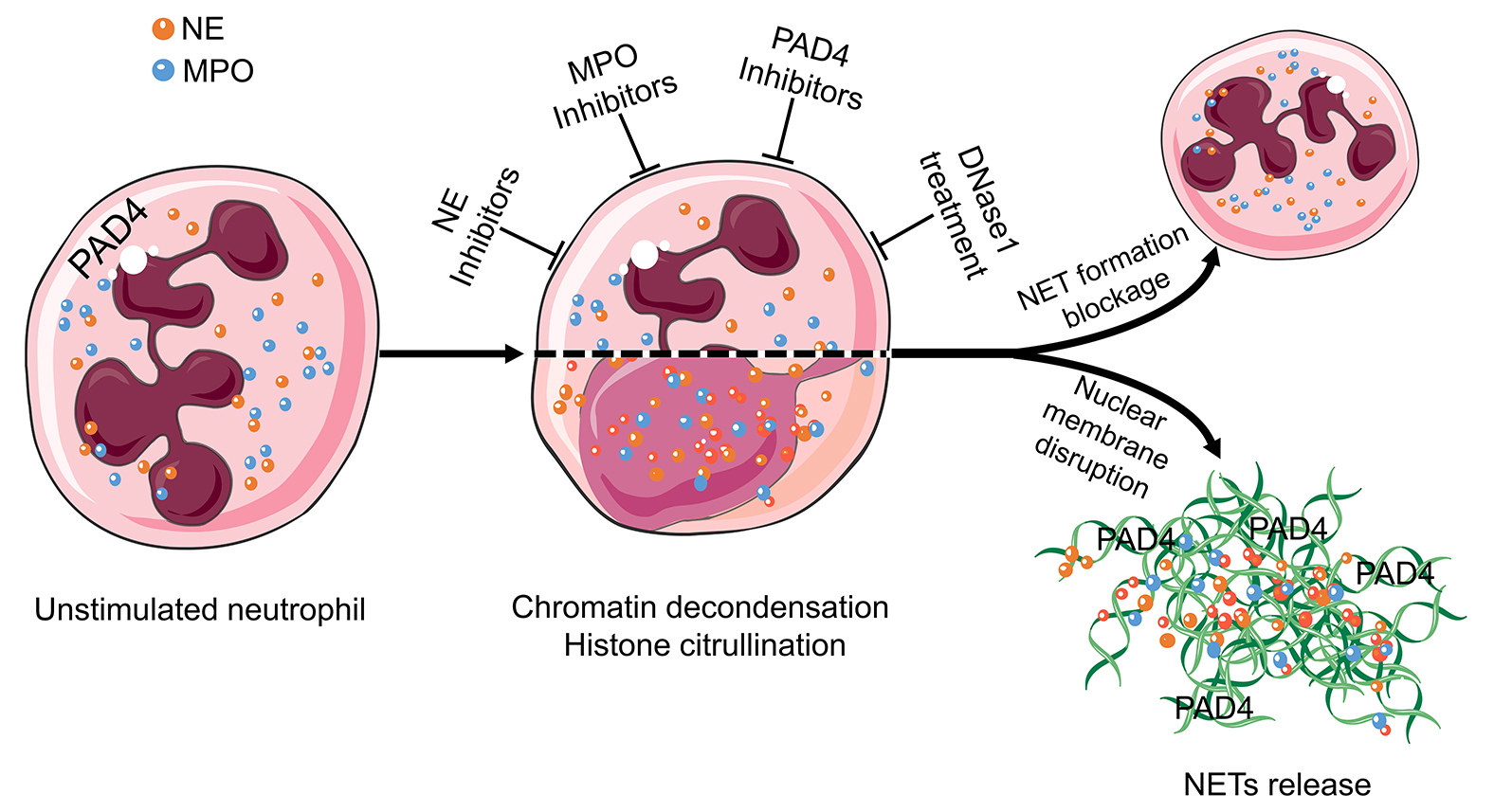
Figure 3 Targets in the treatment of aggNETs-driven occlusive pathologies. NE, MPO, PAD4, and extracellular DNA are promising targets for the suppression of excessive NETs formation. Enzymatic inhibitors of PAD4 and MPO have demonstrated its effectivity in preclinical settings of cholelithiasis, Meibomian gland dysfunction, and vasculitis. DNase treatment is effective at preventing death in severe sepsis and improving the symptoms in cystic fibrosis, bronchiolitis, and dry eye disease. The cartoons were modified from https://smart.servier.com accessed on 20 August 2021 in compliance with the terms of the Creative Commons Attribution 3.0 Unported License (CC BY 3.0). Abbreviations: MPO, myeloperoxidase; NE, neutrophil elastase; NET, neutrophil extracellular traps; and PAD4, peptidylarginine deiminase type IV.
5.1. Inhibition of PAD4 Reduces Formation and Size of NETs
Peptidylarginine deiminases (PADI) are a calcium-dependent enzyme family responsible for post-transcriptional deamination/citrullination. In this process, the positively charged arginine residues are converted into uncharged citrulline residues. The PADI family consists of five members. PAD4 is unique among them. It plays a role in the formation of NETs by ensuring the decondensation of chromatin [97]. This makes PAD4 a possible therapeutic target for the treatment of occlusive NETs-related diseases. The increased activity of PAD4 and the therapeutic potential of its inhibition have already been reported in preclinical settings for the formation of thrombosis [22], acute pancreatitis [70], cholelithiasis [83], meibomian gland disfunction [9], lung injury [98], and sickle cell disease [56].
5.2. Deoxyribonucleases Dismantle NETs
DNases have been studied for therapeutic purposes in obstructive conditions. DNase1 cleaves DNA by breaking its phosphodiester bonds. This disrupts the structural integrity of NETs and reduces the sizes and amounts of aggNETs [99]. It has been shown that disrupting NETs with DNase1 not only prevents vascular occlusion [25] but also recanalizes the already occluded vessel [100]. Similar results have been obtained in crystal clots-driven arterial occlusion [24]. The improvement of ventilation after the inhalation of DNase1 in RSV acute bronchiolitis has also been demonstrated [60]. DNase1 is already approved as an inhalant to reduce the viscosity of the mucus in the lungs of patients with cystic fibrosis [63].
5.3. Inhibitors of Myeloperoxidase Reduce the Early Phases of NET Formation
One of the most important elements of NETs, especially in the early phases, is MPO. When neutrophils encounter danger signals, MPO enters the nucleus and drives chromatin decondensation, a crucial step of NETs formation [101]. Inhibitors of MPO such as PF-1355 have been tried as NETs formation blockers in small vessel vasculitis. In this context, PF-1355 prevented excessive NETs formation and reduced leukocyte infiltration [102], making it a good candidate for further studies trying to interfere with NETs formation in occlusions. Furthermore, natural surfactants also have the ability to inhibit NETs formation in vitro [103]. Therefore, natural surfactants bear the potential to be used as therapeutical agents in occlusive conditions.
6. Conclusions
Neutrophils and NETs participate in the initiation, pathogenesis, and resolution phases of several inflammatory conditions. Considerable collateral damage is expected in most of the cases where neutrophils are involved. Therefore, robust regulatory mechanisms like apoptosis and NETs formation have evolved to endow neutrophils with the ability to drive the amelioration of the initial inflammation. The clearance of apoptotic neutrophils by professional phagocytes triggers potent anti-inflammatory and regenerative responses [104][105]. NETs formation and aggregation actively limit the spreading of pathogens and inflammatory mediators [7][106]. Unfortunately, those anatomical locations carrying fluids or air through the body are prone to be occluded by NETs generating heterogeneous pathologies. The study of the prevention or dissolution of such clogs will provide new therapeutic opportunities for old prevalent diseases.
References
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535.
- Leppkes, M.; Schick, M.; Hohberger, B.; Mahajan, A.; Knopf, J.; Schett, G.; Muñoz, L.E.; Herrmann, M. Updates on NET formation in health and disease. Semin. Arthritis Rheum. 2019, 49, S43–S48.
- Ravindran, M.; Khan, M.A.; Palaniyar, N. Neutrophil Extracellular Trap Formation: Physiology, Pathology, and Pharmacology. Biomolecules 2019, 9, 365.
- Bruschi, M.; Moroni, G.; Sinico, R.A.; Franceschini, F.; Fredi, M.; Vaglio, A.; Cavagna, L.; Petretto, A.; Pratesi, F.; Migliorini, P.; et al. Neutrophil Extracellular Traps in the Autoimmunity Context. Front. Med. 2021, 8.
- Thiam, H.R.; Wong, S.L.; Wagner, D.D.; Waterman, C.M. Cellular Mechanisms of NETosis. Annu. Rev. Cell Dev. Biol. 2020, 36, 191–218.
- Vitkov, L.; Minnich, B.; Knopf, J.; Schauer, C.; Hannig, M.; Herrmann, M. NETs Are Double-Edged Swords with the Potential to Aggravate or Resolve Periodontal Inflammation. Cells 2020, 9, 2614.
- Schauer, C.; Janko, C.; Munoz, L.; Zhao, Y.; Kienhöfer, D.; Frey, B.; Lell, M.; Manger, B.; Rech, J.; Naschberger, E.; et al. Aggregated neutrophil extracellular traps limit inflammation by degrading cytokines and chemokines. Nat. Med. 2014, 20, 511–517.
- Leppkes, M.; Knopf, J.; Naschberger, E.; Lindemann, A.; Singh, J.; Herrmann, I.; Stürzl, M.; Staats, L.; Mahajan, A.; Schauer, C.; et al. Vascular occlusion by neutrophil extracellular traps in COVID-19. EBioMedicine 2020, 58, 102925.
- Mahajan, A.; Hasíková, L.; Hampel, U.; Grüneboom, A.; Shan, X.; Herrmann, I.; Garreis, F.; Bock, F.; Knopf, J.; Singh, J.; et al. Aggregated neutrophil extracellular traps occlude Meibomian glands during ocular surface inflammation. Ocul. Surf. 2021, 20, 1–12.
- Pfeiler, S.; Massberg, S.; Engelmann, B. Biological basis and pathological relevance of microvascular thrombosis. Thromb. Res. 2014, 133, S35–S37.
- Blanch-Ruiz, M.A.; Ortega-Luna, R.; Martinez-Cuesta, M.A.; Alvarez, A. The Neutrophil Secretome as a Crucial Link between Inflammation and Thrombosis. Int. J. Mol. Sci. 2021, 22, 4170.
- Kapoor, S.; Opneja, A.; Nayak, L. The role of neutrophils in thrombosis. Thromb. Res. 2018, 170, 87–96.
- Engelmann, B.; Massberg, S. Thrombosis as an intravascular effector of innate immunity. Nat. Rev. Immunol. 2012, 13, 34–45.
- Gould, T.J.; Lysov, Z.; Liaw, P.C. Extracellular DNA and histones: Double-edged swords in immunothrombosis. J. Thromb. Haemost. 2015, 13, S82–S91.
- Fuchs, T.A.; Brill, A.; Duerschmied, D.; Schatzberg, D.; Monestier, M.; Myers, D.D.; Wrobleski, S.K.; Wakefield, T.W.; Hartwig, J.H.; Wagner, D.D. Extracellular DNA traps promote thrombosis. Proc. Natl. Acad. Sci. USA 2010, 107, 15880–15885.
- Thalin, C.; Hisada, Y.; Lundstrom, S.; Mackman, N.; Wallen, H. Neutrophil Extracellular Traps: Villains and Targets in Arterial, Venous, and Cancer-Associated Thrombosis. Arter. Thromb. Vasc. Biol. 2019, 39, 1724–1738.
- Noubouossie, D.F.; Reeves, B.N.; Strahl, B.D.; Key, N.S. Neutrophils: Back in the thrombosis spotlight. Blood 2019, 133, 2186–2197.
- Arroyo, A.B.; Fernández-Pérez, M.P.; del Monte, A.; Águila, S.; Méndez, R.; Hernández-Antolín, R.; García-Barber, N.; Reyes-García, A.M.D.L.; González-Jiménez, P.; Arcas, M.I.; et al. miR-146a is a pivotal regulator of neutrophil extracellular trap formation promoting thrombosis. Haematologica 2020, 106, 1636–1646.
- Yu, X.; Diamond, S.L. Fibrin Modulates Shear-Induced NETosis in Sterile Occlusive Thrombi Formed under Haemodynamic Flow. Thromb. Haemost. 2019, 119, 586–593.
- Foley, J.H.; Walton, B.L.; Aleman, M.M.; O’Byrne, A.M.; Lei, V.; Harrasser, M.; Foley, K.A.; Wolberg, A.S.; Conway, E.M. Complement Activation in Arterial and Venous Thrombosis is Mediated by Plasmin. EBioMedicine 2016, 5, 175–182.
- Gaul, D.S.; Weber, J.; Van Tits, L.J.; Sluka, S.; Pasterk, L.; Reiner, M.F.; Calatayud, N.; Lohmann, C.; Klingenberg, R.; Pahla, J.; et al. Loss of Sirt3 accelerates arterial thrombosis by increasing formation of neutrophil extracellular traps and plasma tissue factor activity. Cardiovasc. Res. 2018, 114, 1178–1188.
- Sorvillo, N.; Mizurini, D.D.M.; Coxon, C.; Martinod, K.; Tilvawala, R.; Cherpokova, D.; Salinger, A.J.; Seward, R.J.; Staudinger, C.; Weerapana, E.; et al. Plasma Peptidylarginine Deiminase IV Promotes VWF-Platelet String Formation and Accelerates Thrombosis After Vessel Injury. Circ. Res. 2019, 125, 507–519.
- Stark, K.; Philippi, V.; Stockhausen, S.; Busse, J.; Antonelli, A.; Miller, M.; Schubert, I.; Hoseinpour, P.; Chandraratne, S.; Von Brühl, M.-L.; et al. Disulfide HMGB1 derived from platelets coordinates venous thrombosis in mice. Blood 2016, 128, 2435–2449.
- Shi, C.; Kim, T.; Steiger, S.; Mulay, S.R.; Klinkhammer, B.M.; Baeuerle, T.; Melica, M.E.; Romagnani, P.; Möckel, D.; Baues, M.; et al. Crystal Clots as Therapeutic Target in Cholesterol Crystal Embolism. Circ. Res. 2020, 126, e37–e52.
- Jiménez-Alcázar, M.; Rangaswamy, C.; Panda, R.; Bitterling, J.; Simsek, Y.J.; Long, A.T.; Bilyy, R.; Krenn, V.; Renné, C.; Renné, T.; et al. Host DNases prevent vascular occlusion by neutrophil extracellular traps. Science 2017, 358, 1202–1206.
- de Buhr, N.; von Kockritz-Blickwede, M. The Balance of Neutrophil Extracellular Trap Formation and Nuclease Degradation: An Unknown Role of Bacterial Coinfections in COVID-19 Patients? mBio 2021, 12, e03304-20.
- Shi, H.; Zuo, Y.; Gandhi, A.A.; Sule, G.; Yalavarthi, S.; Gockman, K.; Madison, J.A.; Wang, J.; Zuo, M.; Shi, Y.; et al. Endothelial cell-activating antibodies in COVID-19. MedRxiv 2021.
- Ackermann, M.; Anders, H.-J.; Bilyy, R.; Bowlin, G.L.; Daniel, C.; De Lorenzo, R.; Egeblad, M.; Henneck, T.; Hidalgo, A.; Hoffmann, M.; et al. Patients with COVID-19: In the dark-NETs of neutrophils. Cell Death Differ. 2021, 1–15.
- Krauel, K.; Duerschmied, D. ICODE: The international COVID-19 thrombosis biomarkers colloquium—novel soluble biomarkers: Circulating cell-free nucleic acids and other molecules. J. Thromb. Thrombolysis 2021, 1–5.
- Barnes, B.J.; Adrover, J.M.; Baxter-Stoltzfus, A.; Borczuk, A.; Cools-Lartigue, J.; Crawford, J.M.; Daßler-Plenker, J.; Guerci, P.; Huynh, C.; Knight, J.S.; et al. Targeting potential drivers of COVID-19: Neutrophil extracellular traps. J. Exp. Med. 2020, 217, e20200652.
- Veras, F.P.; Pontelli, M.C.; Silva, C.M.; Toller-Kawahisa, J.E.; de Lima, M.; Nascimento, D.C.; Schneider, A.H.; Caetite, D.; Tavares, L.A.; Paiva, I.M.; et al. SARS-CoV-2-triggered neutrophil extracellular traps mediate COVID-19 pathology. J. Exp. Med. 2020, 217, e20201129.
- Radermecker, C.; Detrembleur, N.; Guiot, J.; Cavalier, E.; Henket, M.; D’Emal, C.; Vanwinge, C.; Cataldo, D.; Oury, C.; Delvenne, P.; et al. Neutrophil extracellular traps infiltrate the lung airway, interstitial, and vascular compartments in severe COVID-19. J. Exp. Med. 2020, 217, e20201012.
- Ackermann, M.; Verleden, S.E.; Kuehnel, M.; Haverich, A.; Welte, T.; Laenger, F.; Vanstapel, A.; Werlein, C.; Stark, H.; Tzankov, A.; et al. Pulmonary Vascular Endothelialitis, Thrombosis, and Angiogenesis in Covid-19. N. Engl. J. Med. 2020, 383, 120–128.
- Skendros, P.; Mitsios, A.; Chrysanthopoulou, A.; Mastellos, D.C.; Metallidis, S.; Rafailidis, P.; Ntinopoulou, M.; Sertaridou, E.; Tsironidou, V.; Tsigalou, C.; et al. Complement and tissue factor–enriched neutrophil extracellular traps are key drivers in COVID-19 immunothrombosis. J. Clin. Investig. 2020, 130, 6151–6157.
- Distelmaier, K.; Adlbrecht, C.; Jakowitsch, J.; Winkler, S.; Dunkler, D.; Gerner, C.; Wagner, O.; Lang, I.M.; Kubicek, M. Local complement activation triggers neutrophil recruitment to the site of thrombus formation in acute myocardial infarction. Thromb. Haemost. 2009, 102, 564–572.
- Franck, G.; Mawson, T.; Folco, E.; Molinaro, R.; Ruvkun, V.; Engelbertsen, D.; Liu, X.; Tesmenitsky, Y.; Shvartz, E.; Sukhova, G.; et al. Roles of PAD4 and netosis in experimental atherosclerosis and arterial injury: Implications for superficial erosion. Atherosclerosis 2018, 275, e11.
- Knight, J.S.; Luo, W.; O’Dell, A.A.; Yalavarthi, S.; Zhao, W.; Subramanian, V.; Guo, C.; Grenn, R.C.; Thompson, P.R.; Eitzman, D.T.; et al. Peptidylarginine Deiminase Inhibition Reduces Vascular Damage and Modulates Innate Immune Responses in Murine Models of Atherosclerosis. Circ. Res. 2014, 114, 947–956.
- Riegger, J.; Byrne, R.; Joner, M.; Chandraratne, S.; Gershlick, A.H.; Berg, J.M.T.; Adriaenssens, T.; Guagliumi, G.; Godschalk, T.C.; Neumann, F.-J.; et al. Histopathological evaluation of thrombus in patients presenting with stent thrombosis. A multicenter European study: A report of the prevention of late stent thrombosis by an interdisciplinary global European effort consortium. Eur. Heart J. 2015, 37, 1538–1549.
- de Boer, O.J.; Li, X.; Teeling, P.; Mackaay, C.; Ploegmakers, H.J.; van der Loos, C.M.; Daemen, M.J.; de Winter, R.J.; van der Wal, A.C. Neutrophils, neutrophil extracellular traps and interleukin-17 associate with the organisation of thrombi in acute myocardial infarction. Thromb. Haemost. 2013, 109, 290–297.
- Mangold, A.; Alias, S.; Scherz, T.; Hofbauer, T.M.; Jakowitsch, J.; Panzenböck, A.; Simon, D.; Laimer, D.; Bangert, C.; Kammerlander, A.; et al. Coronary Neutrophil Extracellular Trap Burden and Deoxyribonuclease Activity in ST-Elevation Acute Coronary Syndrome Are Predictors of ST-Segment Resolution and Infarct Size. Circ. Res. 2015, 116, 1182–1192.
- Stakos, D.A.; Kambas, K.; Konstantinidis, T.; Mitroulis, I.; Apostolidou, E.; Arelaki, S.; Tsironidou, V.; Giatromanolaki, A.; Skendros, P.; Konstantinides, S.; et al. Expression of functional tissue factor by neutrophil extracellular traps in culprit artery of acute myocardial infarction. Eur. Heart J. 2015, 36, 1405–1414.
- Blasco, A.; Coronado, M.-J.; Hernández-Terciado, F.; Martín, P.; Royuela, A.; Ramil, E.; García, D.; Goicolea, J.; Del Trigo, M.; Ortega, J.; et al. Assessment of Neutrophil Extracellular Traps in Coronary Thrombus of a Case Series of Patients With COVID-19 and Myocardial Infarction. JAMA Cardiol. 2021, 6, 469.
- Liu, J.; Yang, D.; Wang, X.; Zhu, Z.; Wang, T.; Ma, A.; Liu, P. Neutrophil extracellular traps and dsDNA predict outcomes among patients with ST-elevation myocardial infarction. Sci. Rep. 2019, 9, 11599.
- Perez-De-Puig, I.; Miro-Mur, F.A.; Ferrer-Ferrer, M.; Gelpi, E.; Pedragosa, J.; Justicia, C.; Urra, X.; Chamorro, A.; Planas, A.M. Neutrophil recruitment to the brain in mouse and human ischemic stroke. Acta Neuropathol. 2014, 129, 239–257.
- Deng, J.; Zhao, F.; Zhang, Y.; Zhou, Y.; Xu, X.; Zhang, X.; Zhao, Y. Neutrophil extracellular traps increased by hyperglycemia exacerbate ischemic brain damage. Neurosci. Lett. 2020, 738, 135383.
- Zhou, P.; Li, T.; Jin, J.; Liu, Y.; Li, B.; Sun, Q.; Tian, J.; Zhao, H.; Liu, Z.; Ma, S.; et al. Interactions between neutrophil extracellular traps and activated platelets enhance procoagulant activity in acute stroke patients with ICA occlusion. EBioMedicine 2020, 53, 102671.
- Laridan, E.; Denorme, F.; Desender, L.; François, O.; Andersson, T.; Deckmyn, H.; Vanhoorelbeke, K.; De Meyer, S.F. Neutrophil extracellular traps in ischemic stroke thrombi. Ann. Neurol. 2017, 82, 223–232.
- Essig, F.; Kollikowski, A.M.; Pham, M.; Solymosi, L.; Stoll, G.; Haeusler, K.G.; Kraft, P.; Schuhmann, M.K. Immunohistological Analysis of Neutrophils and Neutrophil Extracellular Traps in Human Thrombemboli Causing Acute Ischemic Stroke. Int. J. Mol. Sci. 2020, 21, 7387.
- Wang, R.; Zhu, Y.; Liu, Z.; Chang, L.; Bai, X.; Kang, L.; Cao, Y.; Yang, X.; Yu, H.; Shi, M.J.; et al. Neutrophil extracellular traps promote tPA-induced brain hemorrhage via cGAS in mice with stroke. Blood 2021, 138, 91–103.
- Nader, E.; Romana, M.; Connes, P. The Red Blood Cell—Inflammation Vicious Circle in Sickle Cell Disease. Front. Immunol. 2020, 11, 454.
- Coates, T.D.; Chalacheva, P.; Zeltzer, L.; Khoo, M.C. Autonomic nervous system involvement in sickle cell disease. Clin. Hemorheol. Microcirc. 2018, 68, 251–262.
- Karsten, E.; Breen, E.; Herbert, B. Red blood cells are dynamic reservoirs of cytokines. Sci. Rep. 2018, 8, 3101.
- Schimmel, M.; Nur, E.; Biemond, B.J.; Van Mierlo, G.J.; Solati, S.; Brandjes, D.P.; Otten, H.-M.; Schnog, J.-J.; Zeerleder, S. Nucleosomes and neutrophil activation in sickle cell disease painful crisis. Haematologica 2013, 98, 1797–1803.
- Barbu, E.A.; Dominical, V.M.; Mendelsohn, L.; Thein, S.L. Neutrophils remain detrimentally active in hydroxyurea-treated patients with sickle cell disease. PLoS ONE 2019, 14, e0226583.
- A Barbu, E.; Mendelsohn, L.; Samsel, L.; Thein, S.L. Pro-inflammatory cytokines associate with NETosis during sickle cell vaso-occlusive crises. Cytokine 2020, 127, 154933.
- Hounkpe, B.W.; Chenou, F.; Domingos, I.D.F.; Msc, E.C.C.; Msc, M.J.D.V.C.S.; Araujo, A.S.; Lucena-Araújo, A.R.; Msc, P.V.D.S.N.; Malheiro, A.; Fraiji, N.A.; et al. Neutrophil extracellular trap regulators in sickle cell disease: Modulation of gene expression of PADI4, neutrophil elastase, and myeloperoxidase during vaso-occlusive crisis. Res. Pract. Thromb. Haemost. 2020, 5, 204–210.
- Cortjens, B.; de Boer, O.; De Jong, R.; Antonis, A.F.G.; Piñeros, Y.S.S.; Lutter, R.; Van Woensel, J.B.M.; Bem, R.A. Neutrophil extracellular traps cause airway obstruction during respiratory syncytial virus disease. J. Pathol. 2015, 238, 401–411.
- Silveira, J.S.; Antunes, G.L.; Gassen, R.B.; Breda, R.V.; Stein, R.T.; Pitrez, P.M.; Da Cunha, A.A. Respiratory syncytial virus increases eosinophil extracellular traps in a murine model of asthma. Asia Pac. Allergy 2019, 9, e32.
- Souza, P.S.S.; Barbosa, L.V.; Diniz, L.F.A.; da Silva, G.S.; Lopes, B.R.P.; Souza, P.M.R.; de Araujo, G.C.; Pessoa, D.; de Oliveira, J.; Souza, F.P.; et al. Neutrophil extracellular traps possess anti-human respiratory syncytial virus activity: Possible interaction with the viral F protein. Virus Res. 2018, 251, 68–77.
- Funchal, G.A.; Jaeger, N.; Czepielewski, R.S.; Machado, M.S.; Muraro, S.; Stein, R.; Bonorino, C.B.C.; Porto, B.N. Respiratory Syncytial Virus Fusion Protein Promotes TLR-4–Dependent Neutrophil Extracellular Trap Formation by Human Neutrophils. PLoS ONE 2015, 10, e0124082.
- Muraro, S.P.; De Souza, G.F.; Gallo, S.W.; Da Silva, B.K.; De Oliveira, S.D.; Vinolo, M.A.; Saraiva, E.M.; Porto, B.N. Respiratory Syncytial Virus induces the classical ROS-dependent NETosis through PAD-4 and necroptosis pathways activation. Sci. Rep. 2018, 8, 14166.
- Besteman, S.B.; Callaghan, A.; Hennus, M.P.; Westerlaken, G.H.; Meyaard, L.; Bont, L.L. Signal inhibitory receptor on leukocytes (SIRL)-1 and leukocyte- associated immunoglobulin-like receptor (LAIR)-1 regulate neutrophil function in infants. Clin. Immunol. 2019, 211, 108324.
- Cortjens, B.; De Jong, R.; Bonsing, J.G.; Van Woensel, J.B.M.; Antonis, A.F.G.; A Bem, R. Local dornase alfa treatment reduces NETs-induced airway obstruction during severe RSV infection. Thorax 2017, 73, 578–580.
- Giglio, M.C.; Cassese, G.; Tomassini, F.; Rashidian, N.; Montalti, R.; Troisi, R.I. Post-operative morbidity following pancreatic duct occlusion without anastomosis after pancreaticoduodenectomy: A systematic review and meta-analysis. HPB 2020, 22, 1092–1101.
- Van Huijgevoort, N.C.; Veld, J.V.; Fockens, P.; Besselink, M.G.; Boermeester, M.A.; Arvanitakis, M.; Van Hooft, J.E. Success of extracorporeal shock wave lithotripsy and ERCP in symptomatic pancreatic duct stones: A systematic review and meta-analysis. Endosc. Int. Open 2020, 8, E1070–E1085.
- Merza, M.; Hartman, H.; Rahman, M.; Hwaiz, R.; Zhang, E.; Renström, E.; Luo, L.; Mörgelin, M.; Regner, S.; Thorlacius, H. Neutrophil Extracellular Traps Induce Trypsin Activation, Inflammation, and Tissue Damage in Mice With Severe Acute Pancreatitis. Gastroenterology 2015, 149, 1920–1931.e8.
- Maueröder, C.; Mahajan, A.S.; Paulus, S.; Gößwein, S.; Hahn, J.; Kienhöfer, D.; Biermann, M.; Tripal, P.; Friedrich, R.P.; Munoz, L.E.; et al. Ménage-à-Trois: The Ratio of Bicarbonate to CO2 and the pH Regulate the Capacity of Neutrophils to Form NETs. Front. Immunol. 2016, 7, 583.
- Leppkes, M.; Maueröder, C.; Hirth, S.; Nowecki, S.; Günther, C.; Billmeier, U.; Paulus, S.; Biermann, M.; Munoz, L.E.; Hoffmann, M.; et al. Externalized decondensed neutrophil chromatin occludes pancreatic ducts and drives pancreatitis. Nat. Commun. 2016, 7, 10973.
- Daniel, C.; Leppkes, M.; Muñoz, L.E.; Schley, G.; Schett, G.; Herrmann, M. Extracellular DNA traps in inflammation, injury and healing. Nat. Rev. Nephrol. 2019, 15, 559–575.
- Madhi, R.; Rahman, M.; Taha, D.; Mörgelin, M.; Thorlacius, H. Targeting peptidylarginine deiminase reduces neutrophil extracellular trap formation and tissue injury in severe acute pancreatitis. J. Cell. Physiol. 2018, 234, 11850–11860.
- Dietrich, J.; Garreis, F.; Paulsen, F. Pathophysiology of Meibomian Glands—An Overview. Ocul. Immunol. Inflamm. 2021, 1–8.
- Chhadva, P.; Goldhardt, R.; Galor, A. Meibomian Gland Disease: The Role of Gland Dysfunction in Dry Eye Disease. Ophthalmology 2017, 124, S20–S26.
- Estúa-Acosta, G.A.; Zamora-Ortiz, R.; Buentello-Volante, B.; García-Mejía, M.; Garfias, Y. Neutrophil Extracellular Traps: Current Perspectives in the Eye. Cells 2019, 8, 979.
- Mahajan, A.; Grüneboom, A.; Petru, L.; Podolska, M.J.; Kling, L.; Maueröder, C.; Dahms, F.; Christiansen, S.; Günter, L.; Krenn, V.; et al. Frontline Science: Aggregated neutrophil extracellular traps prevent inflammation on the neutrophil-rich ocular surface. J. Leukoc. Biol. 2019, 105, 1087–1098.
- Sabeti, S.; Kheirkhah, A.; Yin, J.; Dana, R. Management of meibomian gland dysfunction: A review. Surv. Ophthalmol. 2019, 65, 205–217.
- Baima, G.; Corana, M.; Iaderosa, G.; Romano, F.; Citterio, F.; Meoni, G.; Tenori, L.; Aimetti, M. Metabolomics of gingival crevicular fluid to identify biomarkers for periodontitis: A systematic review with meta-analysis. J. Periodontal Res. 2021, 56, 633–645.
- Bibi, T.; Khurshid, Z.; Rehman, A.; Imran, E.; Srivastava, K.; Shrivastava, D. Gingival Crevicular Fluid (GCF): A Diagnostic Tool for the Detection of Periodontal Health and Diseases. Molecules 2021, 26, 1208.
- Magán-Fernández, A.; Al-Bakri, S.M.R.; O’Valle, F.; Benavides-Reyes, C.; Abadía-Molina, F.; Mesa, F. Neutrophil Extracellular Traps in Periodontitis. Cells 2020, 9, 1494.
- Rudin, A.D.; Amirbeagi, F.; Davidsson, L.; Khamzeh, A.; Mros, S.T.; Thulin, P.; Welin, A.; Björkman, L.; Christenson, K.; Bylund, J. The neutrophil subset defined by CD177 expression is preferentially recruited to gingival crevicular fluid in periodontitis. J. Leukoc. Biol. 2020, 109, 349–362.
- Wang, J.; Zhou, Y.; Ren, B.; Zou, L.; He, B.; Li, M. The Role of Neutrophil Extracellular Traps in Periodontitis. Front. Cell. Infect. Microbiol. 2021, 11, 106.
- Coucke, E.M.; Akbar, H.; Kahloon, A.; Lopez, P.P. Biliary Obstruction; StatPearls: Treasure Island, FL, USA, 2021.
- Shaffer, E.A. Gallstone disease: Epidemiology of gallbladder stone disease. Best Pract. Res. Clin. Gastroenterol. 2006, 20, 981–996.
- Muñoz, L.E.; Boeltz, S.; Bilyy, R.; Schauer, C.; Mahajan, A.; Widulin, N.; Grüneboom, A.; Herrmann, I.; Boada, E.; Rauh, M.; et al. Neutrophil Extracellular Traps Initiate Gallstone Formation. Immunity 2019, 51, 443–450.e4.
- Cho, S.K.; Jung, S.; Lee, K.J.; Kim, J.W. Neutrophil to lymphocyte ratio and platelet to lymphocyte ratio can predict the severity of gallstone pancreatitis. BMC Gastroenterol. 2018, 18, 18.
- O’Neil, L.J.; Kaplan, M.J. NETched in Stone. Immunity 2019, 51, 413–414.
- Ogle, O.E. Salivary Gland Diseases. Dent. Clin. N. Am. 2020, 64, 87–104.
- Bag, A.K.; Curé, J.K.; Chapman, P.R.; Singhal, A.; Mohamed, A.W.H. Imaging of Inflammatory Disorders of Salivary Glands. Neuroimaging Clin. North Am. 2018, 28, 255–272.
- Kraaij, S.; Brand, H.; Van Der Meij, E.; De Visscher, J. Biochemical composition of salivary stones in relation to stone- and patient-related factors. Med. Oral Patol. Oral Cir. Bucal 2018, 23, e540–e544.
- Tretiakow, D.; Skorek, A.; Wysocka, J.; Darowicki, K.; Ryl, J. Classification of submandibular salivary stones based on ultrastructural studies. Oral Dis. 2020.
- Ugga, L.; Ravanelli, M.; Pallottino, A.; Farina, D.; Leone, C. Diagnostic work-up in obstructive and inflammatory salivary gland disorders. Acta Otorhinolaryngol. Ital. 2017, 37, 83–93.
- Fusconi, M.; Petrozza, V.; Schippa, S.; de Vincentiis, M.; Familiari, G.; Pantanella, F.; Cirenza, M.; Iebba, V.; Battaglione, E.; Greco, A.; et al. Bacterial Biofilm in Salivary Gland Stones: Cause or Consequence? Otolaryngol. Head Neck Surg. 2016, 154, 449–453.
- Kao, W.K.; Chole, R.A.; Ogden, M.A. Evidence of a microbial etiology for sialoliths. Laryngoscope 2019, 130, 69–74.
- Schapher, M.; Koch, M.; Weidner, D.; Scholz, M.; Wirtz, S.; Mahajan, A.; Herrmann, I.; Singh, J.; Knopf, J.; Leppkes, M.; et al. Neutrophil Extracellular Traps Promote the Development and Growth of Human Salivary Stones. Cells 2020, 9, 2139.
- Harrison, J.D.; Triantafyllou, A.; Baldwin, D.; Schäfer, H. Histochemical and biochemical determination of calcium in salivary glands with particular reference to chronic submandibular sialadenitis. Virchows Arch. A Pathol. Anat. Histopathol. 1993, 423, 29–32.
- Marchal, F.; Kurt, A.-M.; Dulguerov, P.; Lehmann, W. Retrograde Theory in Sialolithiasis Formation. Arch. Otolaryngol. Head Neck Surg. 2001, 127, 66–68.
- Gierlikowska, B.; Stachura, A.; Gierlikowski, W.; Demkow, U. Phagocytosis, Degranulation and Extracellular Traps Release by Neutrophils-The Current Knowledge, Pharmacological Modulation and Future Prospects. Front. Pharmacol. 2021, 12, 666732.
- Wong, S.L.; Wagner, D.D. Peptidylarginine deiminase 4: A nuclear button triggering neutrophil extracellular traps in inflammatory diseases and aging. FASEB J. 2018, 32, 6258–6370.
- Du, M.; Yang, L.; Gu, J.; Wu, J.; Ma, Y.; Wang, T. Inhibition of Peptidyl Arginine Deiminase-4 Prevents Renal Ischemia-Reperfusion-Induced Remote Lung Injury. Mediat. Inflamm. 2020, 2020, 1724206.
- Podolska, M.J.; Mahajan, A.; Hahn, J.; Knopf, J.; Maueröder, C.; Petru, L.; Ullmann, M.; Schett, G.; Leppkes, M.; Herrmann, M.; et al. Treatment with DNases rescues hidden neutrophil elastase from aggregated NETs. J. Leukoc. Biol. 2019, 106, 1359–1366.
- Peña-Martínez, C.; Durán-Laforet, V.; Garcia-Culebras, A.; Ostos, F.; Hernández-Jiménez, M.; Bravo-Ferrer, I.; Pérez-Ruiz, A.; Ballenilla, F.; Díaz-Guzmán, J.; Pradillo, J.; et al. Pharmacological Modulation of Neutrophil Extracellular Traps Reverses Thrombotic Stroke tPA (Tissue-Type Plasminogen Activator) Resistance. Stroke 2019, 50, 3228–3237.
- Metzler, K.D.; Fuchs, T.A.; Nauseef, W.; Reumaux, D.; Roesler, J.; Schulze, I.; Wahn, V.; Papayannopoulos, V.; Zychlinsky, A. Myeloperoxidase is required for neutrophil extracellular trap formation: Implications for innate immunity. Blood 2011, 117, 953–959.
- Zheng, W.; Warner, R.; Ruggeri, R.; Su, C.; Cortes, C.; Skoura, A.; Ward, J.; Ahn, K.; Kalgutkar, A.; Sun, D.; et al. PF-1355, a mechanism-based myeloperoxidase inhibitor, prevents immune complex vasculitis and anti-glomerular basement membrane glomerulonephritis. J. Pharmacol. Exp. Ther. 2015, 353, 288–298.
- Schulz, A.; Raluy, L.P.; Kolman, J.P.; Konigs, I.; Trochimiuk, M.; Appl, B.; Reinshagen, K.; Boettcher, M.; Trah, J. The Inhibitory Effect of Curosurf((R)) and Alveofact((R)) on the Formation of Neutrophil Extracellular Traps. Front. Immunol. 2020, 11, 582895.
- Stern, M.; Savill, J.; Haslett, C. Human monocyte-derived macrophage phagocytosis of senescent eosinophils undergoing apoptosis. Mediation by alpha v beta 3/CD36/thrombospondin recognition mechanism and lack of phlogistic response. Am. J. Pathol. 1996, 149, 911–921.
- Voll, R.E.; Herrmann, M.; Roth, E.A.; Stach, C.; Kalden, J.R.; Girkontaite, I. Immunosuppressive effects of apoptotic cells. Nature 1997, 390, 350–351.
- Hahn, J.; Schauer, C.; Czegley, C.; Kling, L.; Petru, L.; Schmid, B.; Weidner, D.; Reinwald, C.; Biermann, M.H.C.; Blunder, S.; et al. Aggregated neutrophil extracellular traps resolve inflammation by proteolysis of cytokines and chemokines and protection from antiproteases. FASEB J. 2018, 33, 1401–1414.

