Mitochondria control cellular fate by various mechanisms and are key drivers of cellular metabolism. Although the main function of mitochondria is energy production, they are also involved in cellular detoxification, cellular stabilization, as well as control of ketogenesis and glucogenesis. Conditions like neurodegenerative disease, insulin resistance, endocrine imbalances, liver and kidney disease are intimately linked to metabolic disorders or inflexibility and to mitochondrial dysfunction. Mitochondrial dysfunction due to a relative lack of micronutrients and substrates is implicated in the development of many chronic diseases. l-carnitine is a vital molecule that is found in all living cells. It is a quaternary amine (3-hydroxy-4-N-trimethylaminobutyrate) whose main function in mammalian cells is the transfer of long chain fatty acids across the inner mitochondrial membrane for β- oxidation and generation of ATP energy.
1. Introduction
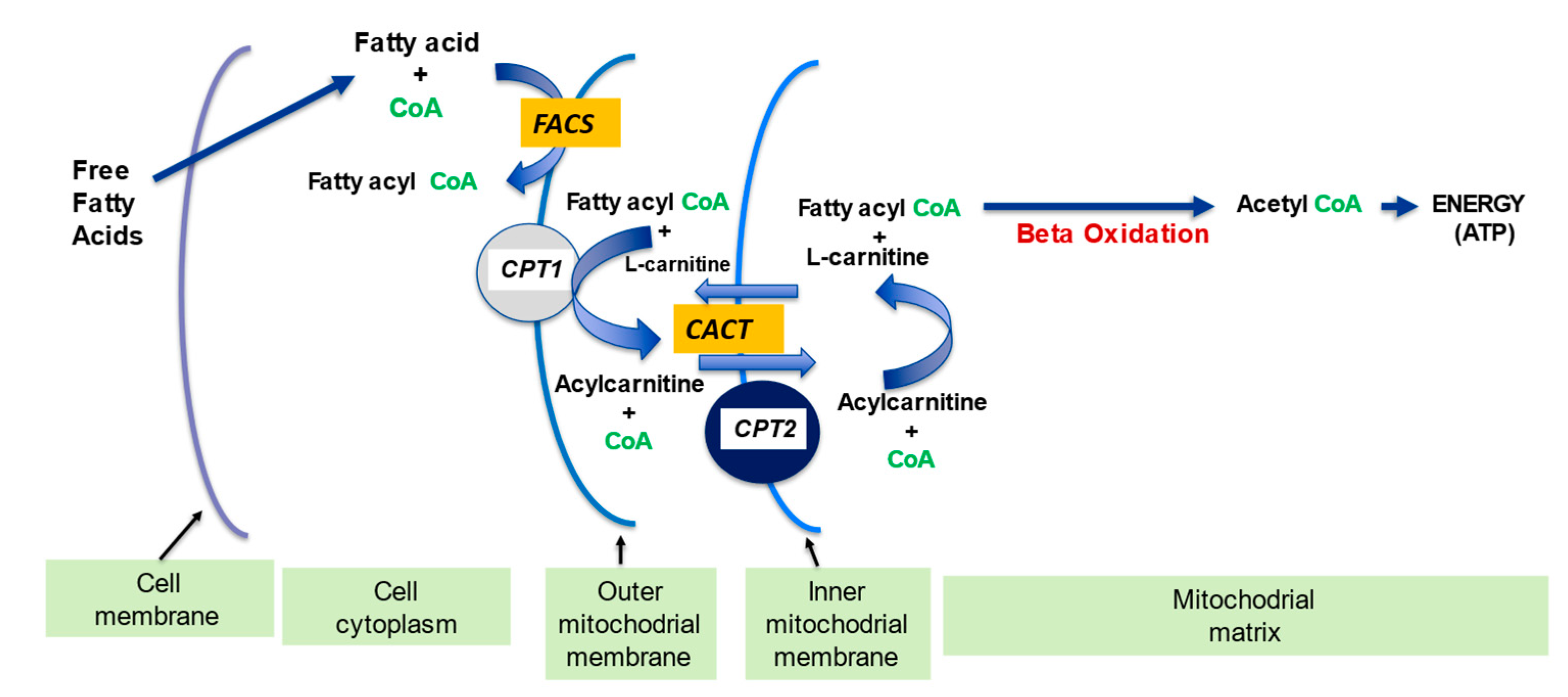
l-carnitine is a vital molecule that is found in all living cells. It is a quaternary amine (3-hydroxy-4-N-trimethylaminobutyrate) whose main function in mammalian cells is the transfer of long chain fatty acids across the inner mitochondrial membrane for β- oxidation and generation of ATP energy. This process requires specific enzymes and transporters and any defects in these can cause disorders of the carnitine cycle. Once fatty acids are inside mitochondria, they undergo the process of β-oxidation through a series of acyl-CoA dehydrogenase enzymes that mediate the shortening of long-chain fats used to produce ATP. Deficiency of any of these enzymes can cause symptoms similar to those encountered in disorders of the carnitine cycle.
l-carnitine is also involved in the excretion of unwanted products of intermediary metabolism and in balancing the Coenzyme A (CoA) pool within mitochondria. Its unique properties enable other functions in cell metabolism such as buffering excess acyl residues and removal of xenobiotics from cells. These actions give l-carnitine a role in protecting cellular membranes, preventing fatty acid accumulation, modulating ketogenesis and glucogenesis and elimination of toxic metabolites.
2. l-Carnitine, Mitochondria and Cellular Metabolism
Metabolism is a coordinated series of chemical reactions that occur in cells of living organisms, needed to create and sustain life. These biochemical processes are important for the storage, freeing, and use of energy, and for the proliferation and repair of cells. The rate of recovery and the overall health of the body are therefore intrinsically linked to the capabilities of these metabolic processes.
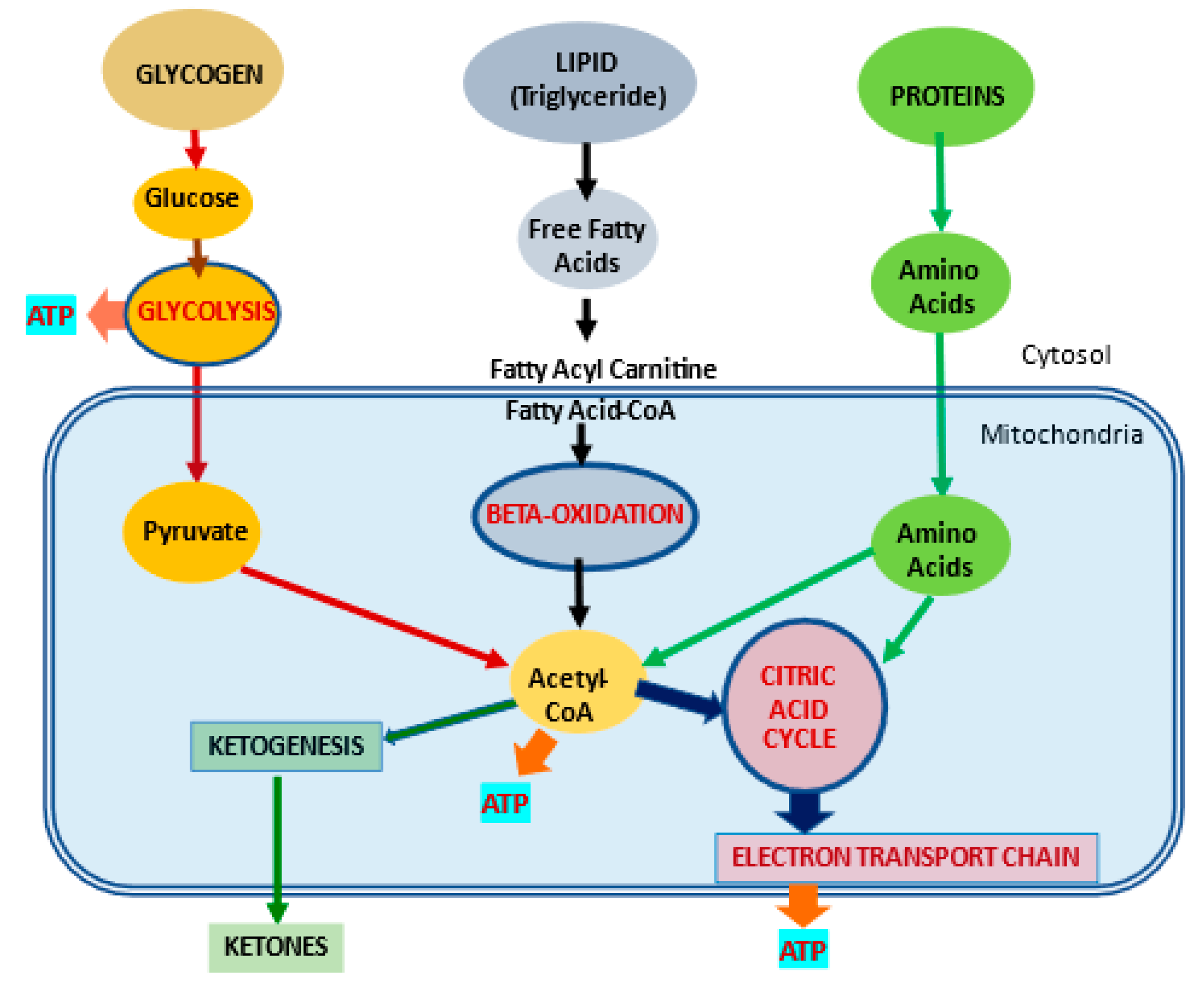
Most of the cellular energy production comes from mitochondria whose main function is the production of adenosine triphosphate (ATP), which is needed to drive cellular processes (Figure 1).
Figure 1. Metabolic pathways for major substrates in the body. Adapted from Virmani et al., 2015
[1]. The body utilizes carbohydrates as glucose, lipids as free fatty acids and proteins as amino acids as the major substrates for the production of ATP energy. Much of the energy from these substrates is produced in the mitochondria.
Mitochondrial dysfunction leads to a loss of energy production, fatty acid accumulation, excess reactive oxygen species(ROS) generation, membrane instability, and a shift to glucose metabolism
[2][1].
l-carnitine is central to mitochondrial processes, fatty acid metabolism, and the production of ATP. It also plays a major part in other important metabolic functions like cellular detoxification, control of ketogenesis and glucogenesis, and stabilization of cell membranes (
Table 1). New research suggests that it also participates in blood glucose homeostasis
[3].
l-carnitine continues to receive scientific attention as a therapy for kidney disease, cardiovascular disease, and diabetes, as well as symptoms associated with carnitine deficiency and mitochondrial disorders
[4][5][6].
Table 1. Major l-carnitine functions in cellular metabolism.
| l-Carnitine (LC) Function |
Description |
References |
| Fatty acid metabolism |
LC transports LCFA across inner mitochondrial membrane for subsequent beta oxidation and ATP production |
[4][5] |
| Regulation of mitochondrial acetyl-CoA/CoA ratio and acyl-CoA/CoA ratio |
LC forms an effective transport system for acetyl or acyl groups out of the mitochondria to maintain free CoA levels important for glycolysis and other processes |
[1][5] |
| Detoxification of potentially toxic metabolite |
LC binds acyl residues and helps in their elimination thereby maintaining metabolic flexibility |
[7] |
| Stabilization of cell membranes |
LC helps stabilize cell membranes via its effects on acetylation of membrane phospholipids and surface membrane effects |
[8][9] |
| Control of ketogenesis and gluconeogenesis |
LC affects ketogenesis by transporting free fatty acids into mitochondria for subsequent use in the production of ketones in the mitochondria. |
[10] |
2.2. Regulation of the Mitochondrial Acetyl-CoA/CoA Ratio and Acyl-CoA/CoA Ratio
l-carnitine has a key role in the regulation of acyl-CoA and acetyl-CoA, and in the preservation of free CoA levels within the mitochondria, which is crucial for maintaining metabolic flexibility.
l-carnitine plays a part in the carrier of activated acetyl and acyl groups. In partnership with the carnitine transferase enzyme (CPT) and carnitine translocase (CACT),
l-carnitine forms an effective transport system for acetyl groups (
Figure 3) or acyl groups (Figure 7) out of the mitochondria, thus preventing diminution of intramitochondrial free CoA. The buffering of the acetyl-CoA and acyl-CoA levels is important in allowing CoA-dependent reactions, especially glycolysis and the pyruvate oxidation to continue optimally
[1][13].
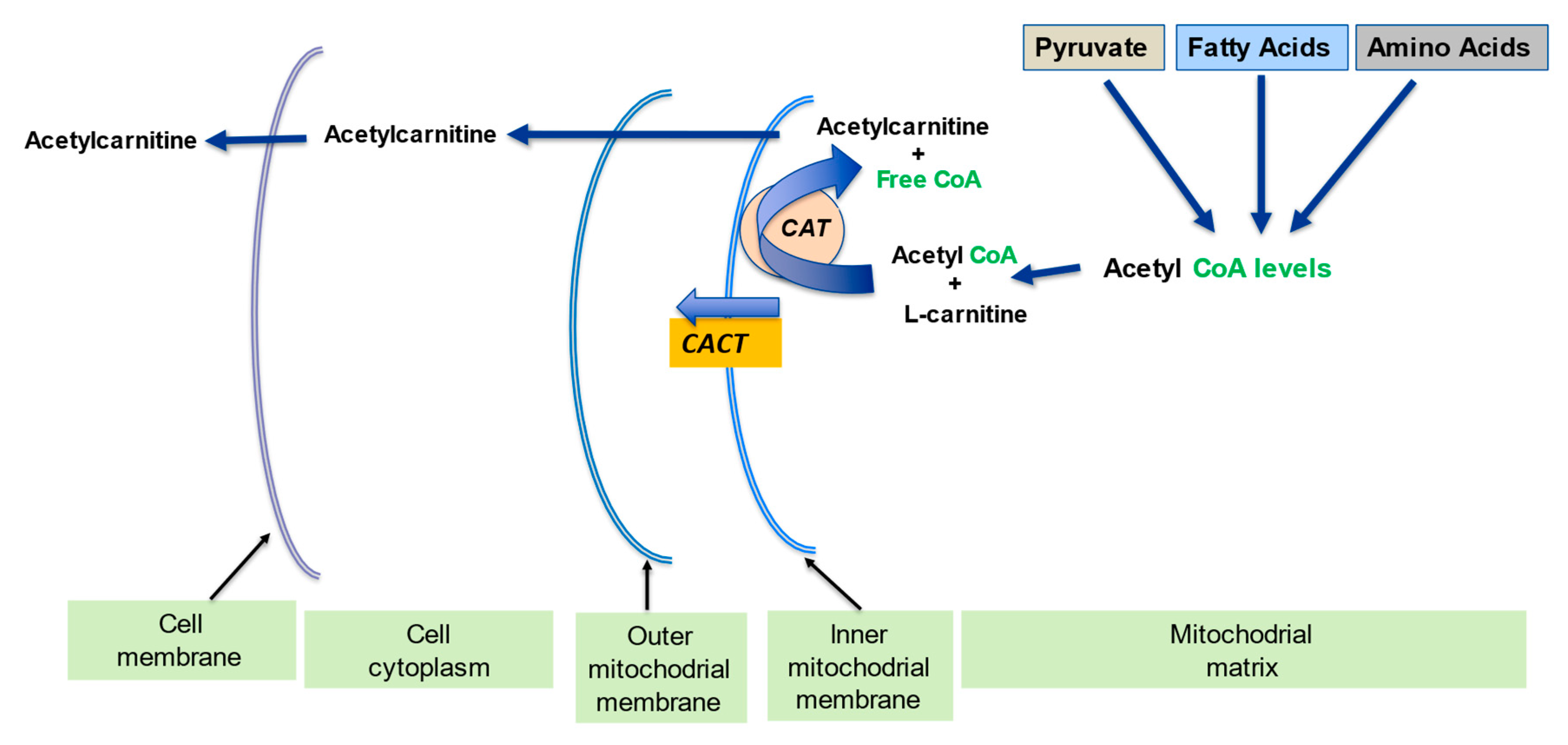
Figure 3. l-carnitine regulates the intramitochondrial acetyl-CoA/CoA ratio. Adapted from Virmani et al., 2015
[1]. The acetyl-CoA build up from pyruvate, fatty acids amino acids metabolism becomes a limiting step since free mitochondrial CoA levels decline.
l-carnitine converts the acetyl-CoA to acetyl-
l-carnitine, which can be transported out of the mitochondrial matrix, and in this way frees up the CoA.
The CACT translocase and CAT carnitine transferase, like all enzymes, can work in the reverse direction depending on the conditions. In this way, activated short, medium and long-chain acyl groups can be transported out of the mitochondrial matrix. In certain situations, cellular acyl-CoAs build-up as a result of an inborn error of fatty acid metabolism, and this also leads to an increase in acylcarnitines within the cell
[12][14][15].
2.3. Detoxification of Toxic Metabolites
The build-up of potentially toxic metabolites in the cell due to mitochondrial dysfunction and other mechanisms can also alter metabolic flexibility.
l-carnitine plays a part in removing potentially toxic metabolites associated with the β-oxidation of fatty acids (Figure 7).
l-carnitine binds acyl residues arising from the intermediary metabolism of amino acids and helps in their elimination
[16]. This mechanism is also essential in sequestering and subsequent removal of abnormal organic acids in several organic acidemia
[8].
l-carnitine coupling decreases the number of long chain acyl residues attached to CoA and increases the free CoA to acyl-CoA’s ratio
[9]. This process is essential in disorders of mitochondrial fatty acids oxidation where the fatty acyl-CoAs accrual in specific tissues such as in the heart and central nervous system can induce toxicity, trigger apoptosis and inflammation, and destabilize mitochondrial membrane
[17][18]. For this reason, the conjugation of specific chain length acyls with
l-carnitine creates very particular species of acylcarnitines that reflect any inborn errors of metabolism; thereby, they can also be utilized as markers for the screening of newborns
[8][10].
2.4. Stabilization of Cell Membranes
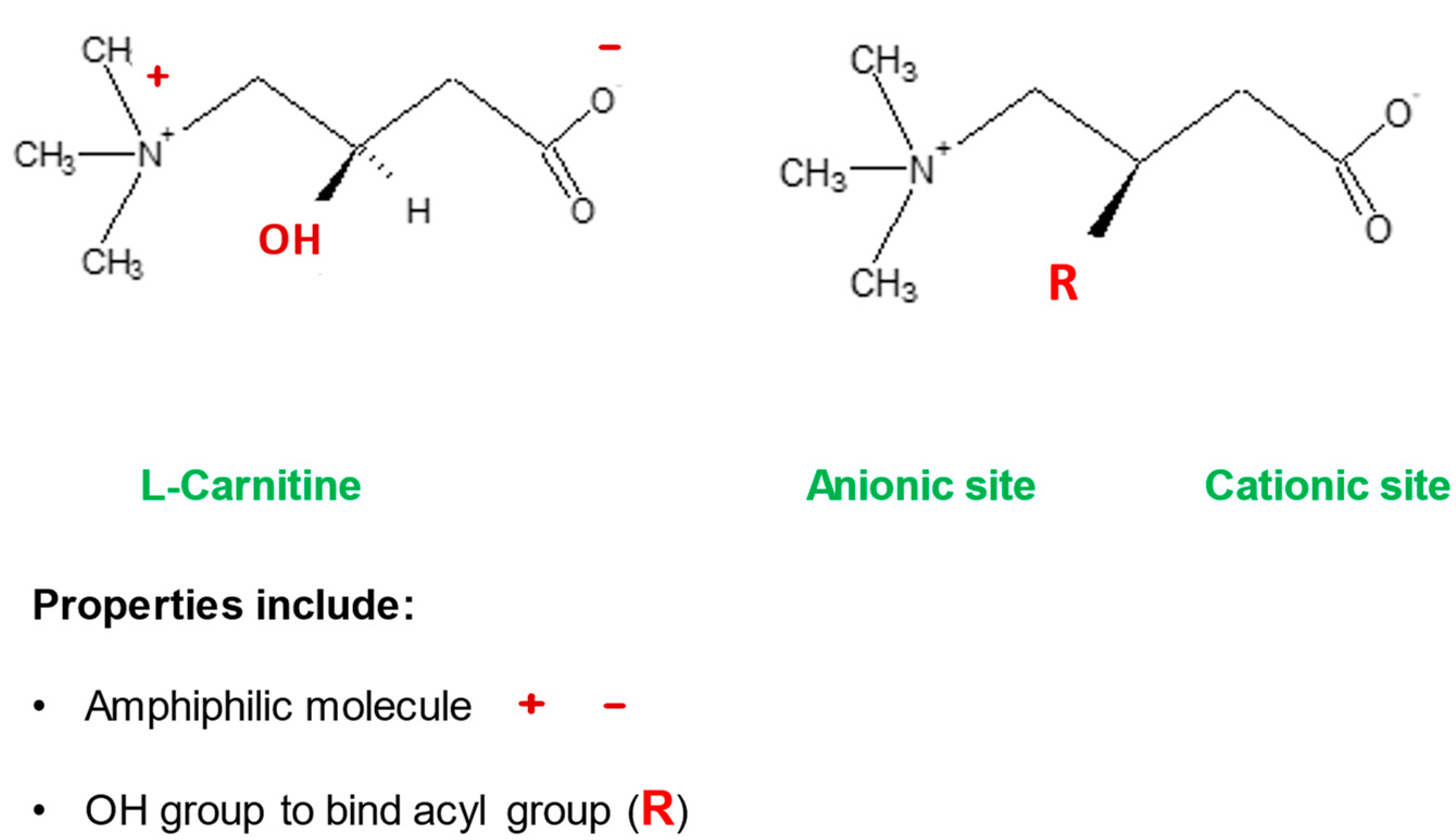
l-carnitine is also essential in maintaining membrane stability and function of plasma, mitochondria, and other organelles possibly via effects on acetylation of membrane phospholipids. Its amphiphilic nature would also allow for interaction with the surface charges on the cell membrane and may play a role in membrane stabilization
[19][20]. The charged tri-methylamino group and the carboxylic group on
l-carnitine would permit interaction with corresponding poles on the membrane phospholipids, glycolipids, and proteins (
Figure 4).
Figure 4. The amphiphilic nature of the
l-carnitine molecule. Adapted from Virmani and Binienda, 2004
[20]. The unique amphiphilic nature of
l-carnitine is attributed to the charges on its carboxyl and amine groups. The hydroxyl group gives it the ability to bind to fatty acids.
2.5. Control of Ketogenesis and Gluconeogenesis
Depending on the body state, e.g., fed or unfed, the liver regulates glucose and fatty acid synthesis, uptake, or release (
Figure 5). In the postprandial (fed) state, the liver produces glucose, stops fatty acid synthesis, starts fatty acid oxidation, and produces ketone bodies
[21].
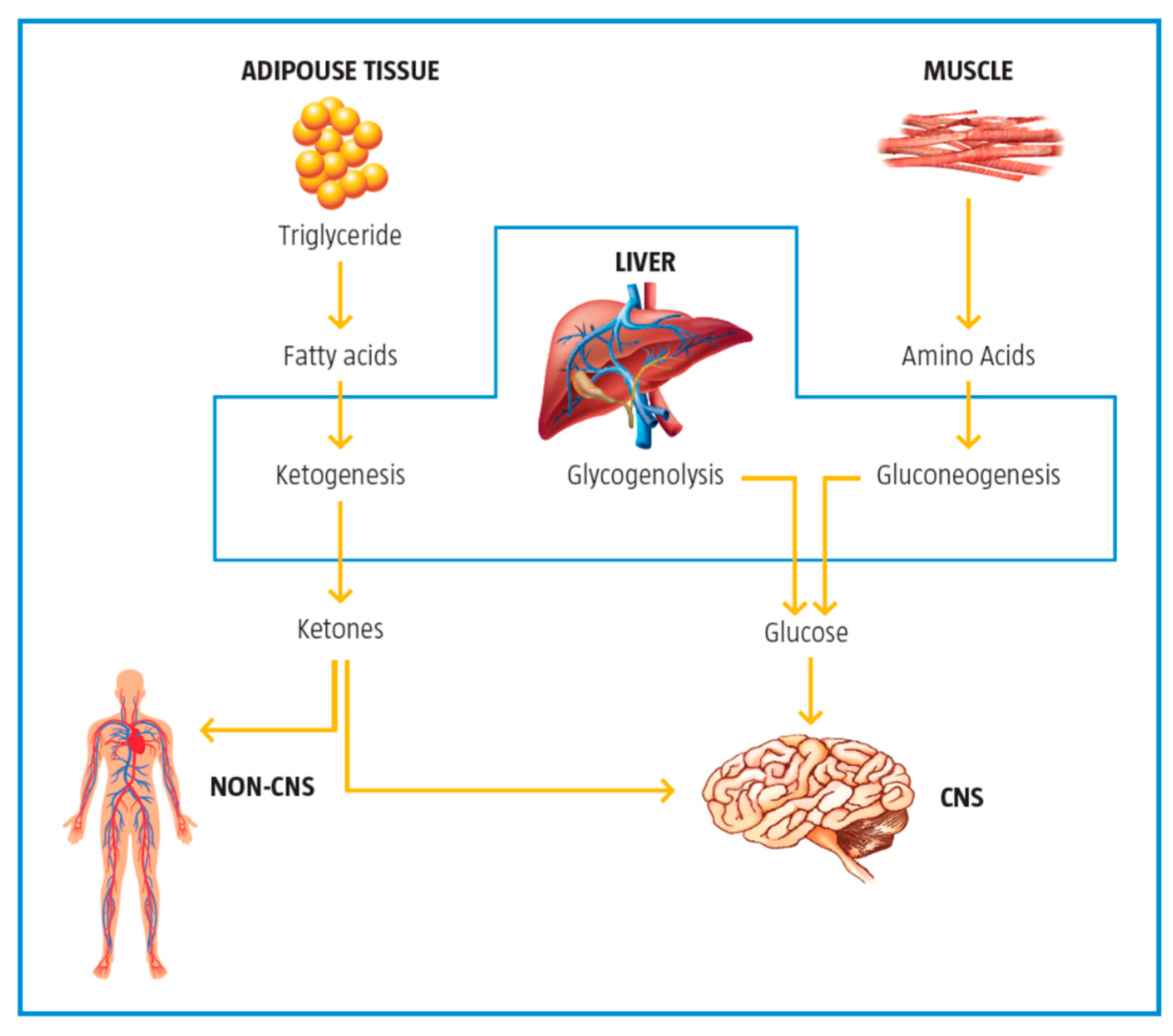
Figure 5. The role of the liver in glucose and fat metabolism. Adapted from Foster, 2004
[21].
During the catabolic state, the liver is a net producer of glucose by both glycogen breakdown and gluconeogenesis with more than half of the glucose produced being used by the brain to produce energy. Fatty acids from triglycerides are converted to ketones in the liver and are used as the second readily available fuel by the brain.
Ketogenesis occurs primarily in the mitochondria of liver cells, and CPT-1 is the rate limiting enzyme in this process. Fatty acids brought into the mitochondria via CPT-1 are broken down into acetyl CoA via β-oxidation. Two acetyl-CoA molecules are converted into acetoacetyl-CoA, which is then converted to HMG-CoA and subsequently to acetoacetate. Once acetoacetate reaches extrahepatic tissues, it is converted back to acetyl-CoA that can enter the citric acid cycle to produce ATP.
Ketogenesis is mainly modulated by insulin but also by other hormones like glucagon, cortisol, thyroid hormones, and catecholamines, which can increase free fatty acids availability for the ketogenic pathway
[21][22].
Mitochondria are also involved in insulin signaling. A number of key enzymes in the ketogenic pathway are activated when insulin levels are low. This low insulin state produces an increase in free fatty acids that are taken up by the mitochondria for the consequent increase in ketone body synthesis.
l-carnitine affects ketogenesis via its part in the uptake of free fatty acids and their subsequent use in the production of ketones. Ketone body production is affected by
l-carnitine status. In carnitine deficient states, long-chain fatty acyl-CoAs cannot be efficiently transported into mitochondria, thereby limiting the ketone body production
[23].
3. Metabolic Inflexibility
Metabolic flexibility is the capacity of the body to use different energy sources depending on the circumstances and availability of oxygen, substrates, and nutrient cofactors. The inability to adapt to the different cellular and environmental needs/stressors results in metabolic inflexibility, which ultimately leads to disease states like insulin resistance, lipid accumulation, and an inability to maintain homeostatic balance
[24][25][26] (
Figure 6).
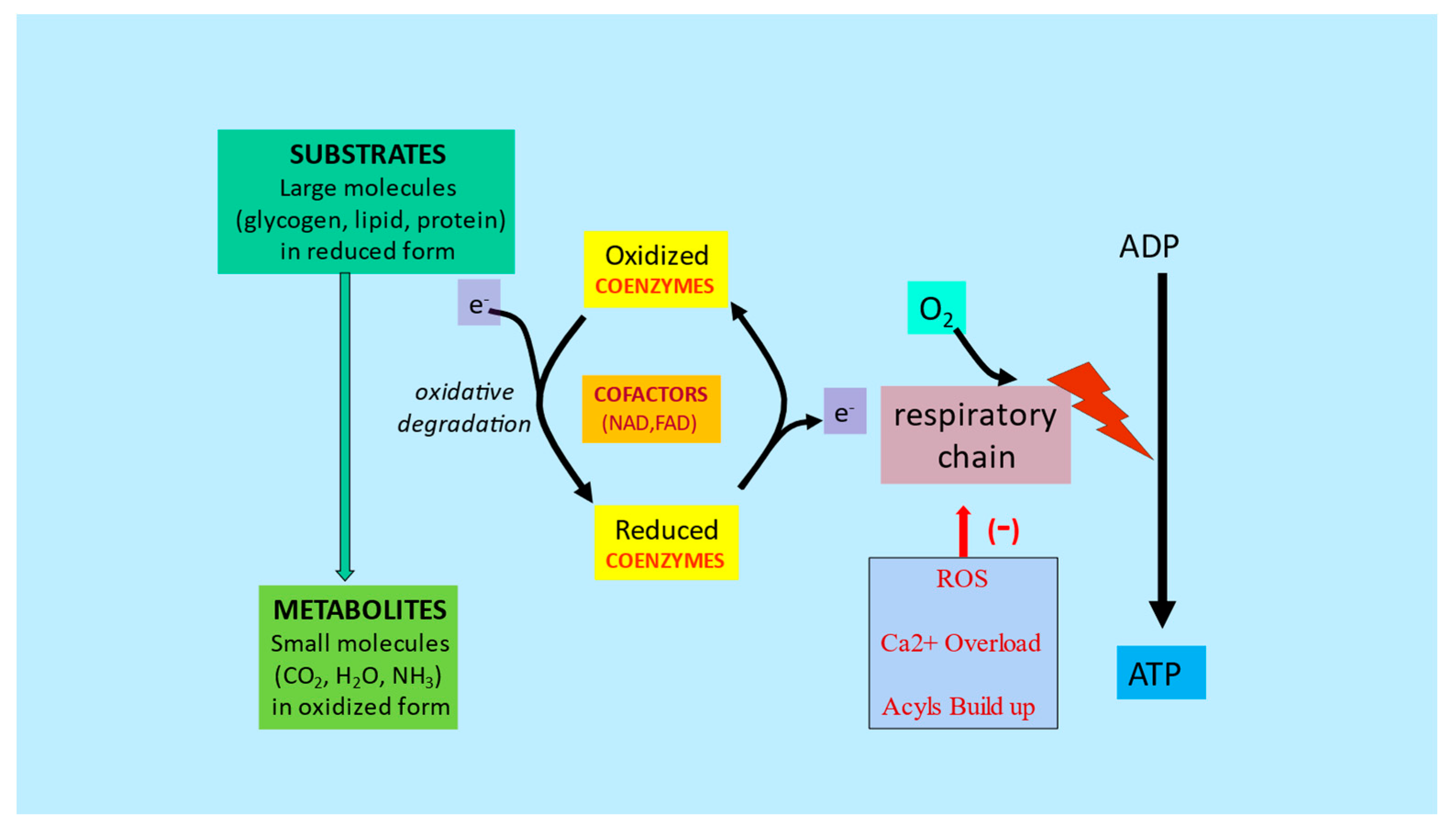
Figure 6. Substrates and nutrient cofactors in mitochondrial function and metabolic flexibility. Adapted from Virmani et al., 2013
[27]. Energy from starting substrates such as glycogen, lipids, and proteins are gradually extracted by the complex series of enzymes in the cellular cytoplasm and mitochondria, resulting in metabolites, mainly CO
2 and H
2O. Any dysfunction in this chain of events caused by lack of cofactors or mitochondrial dysfunction would lead to reduced ATP formation and increased ROS, Ca
2+ and acyls buildup.
The onset of metabolic inflexibility has been linked to micronutrient deficiencies and nutrient overload, as well as the inevitable aging process. Recent research suggests that this process can be delayed or even reversed. To maintain metabolic flexibility, mitochondria need substrates, oxygen, and cofactors for energy production. Altered mitochondrial function leads to a reduction in energy production and an increase in reactive oxygen species, which negatively impacts all cellular functions such as calcium handling, anaplerosis, proteostasis, apoptosis, and autophagy
[27][28][29].
An imbalanced diet with a high intake of processed foods and fat leads to the phenomenon known as “empty calories”. Although foods with empty calories provide immediate energy, they cannot be used efficiently for maintaining body homeostasis and are ultimately stored as fat. Chronic overnutrition triggers mitochondrial metabolic confusion and reduced metabolic efficiency. This results in the accumulation of electrons which can trigger ROS generation, as well as an accumulation of acyl-CoA which further disrupts mitochondrial function and signaling. An inefficient mitochondrial buffering and repair system can ultimately lead to cellular dysfunction
[24][27].
Overeating and lack of exercise can lead to an excess of energy intake relative to body requirements. This can cause a malfunctioning of body control systems altering the delicate balance between requirements and expenditure. The systems controlling the body’s energy requirements are complex and they involve integration of metabolic and neuroendocrine signals. This occurs mainly in the brain and in particular in the hypothalamus; however, it also involves sensors at the cellular and organelle level in the liver, pancreas, muscles, adipose tissues, and gut
[30][31]. Mitochondrial dynamics also play an important role in energy homeostasis due to their ATP producing capacity
[32].
Mitochondrial dysfunction may underly cellular senescence when cells accumulate damage and lose their metabolic flexibility
[33][34]. When cellular metabolism is efficient, senescent cells are removed from the body by apoptosis. If these cells are not eliminated, it can result in disease states like cancer, kidney, liver, and heart disease.
Together with body control systems, mitochondria are key players in maintaining energy requirements and cell function. This will ensure a healthy body homeostasis and prevent the initiation of disease processes. To maintain metabolic energy equilibrium, it is necessary to eat the required type and amount of food, avoid eating empty calorie foods, and ensure correct micronutrient intake. Improving mitochondria function with a balanced diet and lifestyle but also correcting nutrient deficiencies can play a crucial role in maintaining metabolic flexibility and health
[35][36].
 +1 credit
+1 credit