 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Gang-Jee Ko | + 1633 word(s) | 1633 | 2022-01-11 03:54:08 | | | |
| 2 | Yvaine Wei | Meta information modification | 1633 | 2022-01-21 01:42:56 | | |
Video Upload Options
Metabolic syndrome is a cluster of metabolic indicators that increase the risk of diabetes and cardiovascular diseases. Visceral obesity and factors derived from altered adipose tissue, adipokines, play critical roles in the development of metabolic syndrome. Although the adipokines leptin and adiponectin improve insulin sensitivity, others contribute to the development of glucose intolerance, including visfatin, fetuin-A, resistin, and plasminogen activator inhibitor-1 (PAI-1). Leptin and adiponectin increase fatty acid oxidation, prevent foam cell formation, and improve lipid metabolism, while visfatin, fetuin-A, PAI-1, and resistin have pro-atherogenic properties.
1. Introduction
Metabolic syndrome is a combination of interrelated conditions that often occur together, including obesity, insulin resistance, glucose intolerance, hypertension, and dyslipidemia [1]. Metabolic syndrome is diagnosed as the presence of at least three of the following five characteristics: high waist–hip ratio, high blood pressure, elevated blood sugar level, increased triglycerides (TGs), and low high-density lipoprotein (HDL) cholesterol [2]. Metabolic syndrome is important because of its association with an increasing prevalence of diabetes and a higher risk of cardiovascular events such as heart disease and stroke, which have become major public health issues [3]. Dysregulation of certain adipokines can promote pathogenic conditions associated with obesity, lipid accumulation, and insulin resistance. These increase the risk of atherosclerosis [4].
2. Adipose Tissue as a Critical Endocrine Organ Causing Metabolic Syndrome
3. Dysregulation of Adipokines in Metabolic Syndrome
3.1. Leptin
3.2. Adiponectin
3.3. Visfatin
3.4. Fetuin-A
3.5. Plasminogen Activator Inhibitor-1 (PAI-1)
3.6. Omentin-1
3.7. Lipocalin-2
4. Molecular and Cellular Crosstalk in Central Obesity and Metabolic Syndrome
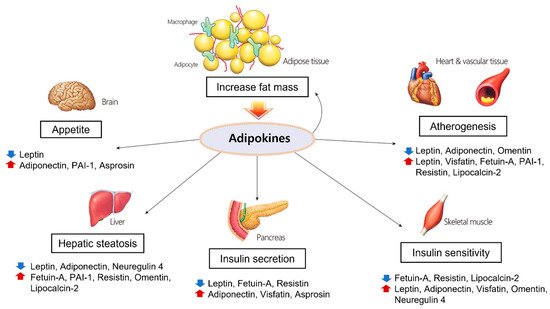
5. The Roles and Associated Mechanisms of Adipokines in Cardiovascular Diseases
5.1. Adiponectin
Adiponectin knockout mice exhibited a significantly increased expression of E-selectin, which is implicated in leukocyte rolling and leukocyte adhesion [36]. Aortic ring tissues derived from adiponectin knockout mice showed a decrease in endothelial NOS expression that might cause a defect in vasodilation. This was reversed by treatment with recombinant adiponectin [37]. In another study, adiponectin administration in obese rats increased endothelial NOS by activating the AMPK pathway and promoting NO production. This resulted in the relaxation of the aortic ring [38]. In cultured human umbilical vein endothelial cells, adiponectin showed a protective effect against angiotensin-II-induced vascular endothelial damage [39]. In addition, adiponectin attenuated angiotensin-II-induced NADPH oxidase activation in renal proximal tubular cells [40]. These studies suggest that adiponectin production is closely related to endothelial function in vasodilation.
5.2. Leptin
Leptin increased the vasodilatation of rat aortic rings in vitro via a nitric oxide (NO)-dependent mechanism [41], but another study showed that leptin had no effect on hemodynamics, even after blocking NO generation [42]. Moreover, leptin synthesis was found to increase when cultured with angiotensin II adipose cells and rats in vivo [43]. Leptin with adipose-tissue-derived angiotensin II can promote obesity-related hypertension [44]. In an in vitro study using endothelial cells of the human umbilical vein, leptin induced chronic oxidative stress in endothelial cells and promoted atherogenesis [45].
5.3. PAI-1
5.4. Omentin-1
5.5. Lipocalin-2
References
- Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z. The metabolic syndrome. Lancet 2005, 365, 1415–1428.
- Alberti, K.G.; Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z.; Cleeman, J.I.; Donato, K.A.; Fruchart, J.C.; James, W.P.; Loria, C.M.; Smith, S.C., Jr.; et al. Harmonizing the metabolic syndrome: A joint interim statement of the International Diabetes Federation Task Force on Epidemiology and Prevention; National Heart, Lung, and Blood Institute; American Heart Association; World Heart Federation; International Atherosclerosis Society; and International Association for the Study of Obesity. Circulation 2009, 120, 1640–1645.
- Li, X.; Zhai, Y.; Zhao, J.; He, H.; Li, Y.; Liu, Y.; Feng, A.; Li, L.; Huang, T.; Xu, A.; et al. Impact of Metabolic Syndrome and It’s Components on Prognosis in Patients With Cardiovascular Diseases: A Meta-Analysis. Front. Cardiovasc. Med. 2021, 8, 704145.
- Kahn, C.R.; Wang, G.; Lee, K.Y. Altered adipose tissue and adipocyte function in the pathogenesis of metabolic syndrome. J. Clin. Investig. 2019, 129, 3990–4000.
- Hu, J.; Yang, S.; Zhang, A.; Yang, P.; Cao, X.; Li, X.; Goswami, R.; Wang, Y.; Luo, T.; Liao, K.; et al. Abdominal Obesity Is More Closely Associated With Diabetic Kidney Disease Than General Obesity. Diabetes Care 2016, 39, e179–e180.
- Fan, H.; Li, X.; Zheng, L.; Chen, X.; Lan, Q.; Wu, H.; Ding, X.; Qian, D.; Shen, Y.; Yu, Z.; et al. Abdominal obesity is strongly associated with Cardiovascular Disease and its Risk Factors in Elderly and very Elderly Community-dwelling Chinese. Sci. Rep. 2016, 6, 21521.
- Smith, U. Abdominal obesity: A marker of ectopic fat accumulation. J. Clin. Investig. 2015, 125, 1790–1792.
- Neeland, I.J.; Ross, R.; Despres, J.P.; Matsuzawa, Y.; Yamashita, S.; Shai, I.; Seidell, J.; Magni, P.; Santos, R.D.; Arsenault, B.; et al. Visceral and ectopic fat, atherosclerosis, and cardiometabolic disease: A position statement. Lancet Diabetes Endocrinol. 2019, 7, 715–725.
- Abate, N.; Garg, A.; Peshock, R.M.; Stray-Gundersen, J.; Adams-Huet, B.; Grundy, S.M. Relationship of generalized and regional adiposity to insulin sensitivity in men with NIDDM. Diabetes 1996, 45, 1684–1693.
- Scheja, L.; Heeren, J. The endocrine function of adipose tissues in health and cardiometabolic disease. Nat. Rev. Endocrinol. 2019, 15, 507–524.
- Thomas, D.; Apovian, C. Macrophage functions in lean and obese adipose tissue. Metabolism 2017, 72, 120–143.
- Appari, M.; Channon, K.M.; McNeill, E. Metabolic Regulation of Adipose Tissue Macrophage Function in Obesity and Diabetes. Antioxid. Redox Signal. 2018, 29, 297–312.
- De Ferranti, S.; Mozaffarian, D. The perfect storm: Obesity, adipocyte dysfunction, and metabolic consequences. Clin. Chem. 2008, 54, 945–955.
- Munzberg, H.; Morrison, C.D. Structure, production and signaling of leptin. Metabolism 2015, 64, 13–23.
- Maeda, K.; Okubo, K.; Shimomura, I.; Funahashi, T.; Matsuzawa, Y.; Matsubara, K. cDNA cloning and expression of a novel adipose specific collagen-like factor, apM1 (AdiPose Most abundant Gene transcript 1). Biochem. Biophys. Res. Commun. 1996, 221, 286–289.
- Hada, Y.; Yamauchi, T.; Waki, H.; Tsuchida, A.; Hara, K.; Yago, H.; Miyazaki, O.; Ebinuma, H.; Kadowaki, T. Selective purification and characterization of adiponectin multimer species from human plasma. Biochem. Biophys. Res. Commun. 2007, 356, 487–493.
- Dahl, T.B.; Holm, S.; Aukrust, P.; Halvorsen, B. Visfatin/NAMPT: A multifaceted molecule with diverse roles in physiology and pathophysiology. Annu. Rev. Nutr. 2012, 32, 229–243.
- Romacho, T.; Sanchez-Ferrer, C.F.; Peiro, C. Visfatin/Nampt: An adipokine with cardiovascular impact. Mediat. Inflamm. 2013, 2013, 946427.
- Curat, C.A.; Wegner, V.; Sengenes, C.; Miranville, A.; Tonus, C.; Busse, R.; Bouloumie, A. Macrophages in human visceral adipose tissue: Increased accumulation in obesity and a source of resistin and visfatin. Diabetologia 2006, 49, 744–747.
- Chatterjee, P.; Seal, S.; Mukherjee, S.; Kundu, R.; Mukherjee, S.; Ray, S.; Mukhopadhyay, S.; Majumdar, S.S.; Bhattacharya, S. Adipocyte fetuin-A contributes to macrophage migration into adipose tissue and polarization of macrophages. J. Biol. Chem. 2013, 288, 28324–28330.
- Dasgupta, S.; Bhattacharya, S.; Biswas, A.; Majumdar, S.S.; Mukhopadhyay, S.; Ray, S.; Bhattacharya, S. NF-kappaB mediates lipid-induced fetuin-A expression in hepatocytes that impairs adipocyte function effecting insulin resistance. Biochem. J. 2010, 429, 451–462.
- Kim, J.A.; Choi, K.M. Newly Discovered Adipokines: Pathophysiological Link between Obesity and Cardiometabolic Disorders. Front. Physiol. 2020, 11, 568800.
- Antonopoulos, A.S.; Tousoulis, D. The molecular mechanisms of obesity paradox. Cardiovasc. Res. 2017, 113, 1074–1086.
- Dhawan, D.; Sharma, S. Abdominal Obesity, Adipokines and Non-communicable Diseases. J. Steroid Biochem. Mol. Biol. 2020, 203, 105737.
- Kwok, K.H.; Lam, K.S.; Xu, A. Heterogeneity of white adipose tissue: Molecular basis and clinical implications. Exp. Mol. Med. 2016, 48, e215.
- Unamuno, X.; Gomez-Ambrosi, J.; Rodriguez, A.; Becerril, S.; Fruhbeck, G.; Catalan, V. Adipokine dysregulation and adipose tissue inflammation in human obesity. Eur. J. Clin. Investig. 2018, 48, e12997.
- Antonopoulos, A.S.; Margaritis, M.; Coutinho, P.; Digby, J.; Patel, R.; Psarros, C.; Ntusi, N.; Karamitsos, T.D.; Lee, R.; De Silva, R.; et al. Reciprocal effects of systemic inflammation and brain natriuretic peptide on adiponectin biosynthesis in adipose tissue of patients with ischemic heart disease. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 2151–2159.
- Hocking, S.L.; Stewart, R.L.; Brandon, A.E.; Suryana, E.; Stuart, E.; Baldwin, E.M.; Kolumam, G.A.; Modrusan, Z.; Junutula, J.R.; Gunton, J.E.; et al. Subcutaneous fat transplantation alleviates diet-induced glucose intolerance and inflammation in mice. Diabetologia 2015, 58, 1587–1600.
- Skurk, T.; Alberti-Huber, C.; Herder, C.; Hauner, H. Relationship between adipocyte size and adipokine expression and secretion. J. Clin. Endocrinol. Metab. 2007, 92, 1023–1033.
- Wang, B.; Wood, I.S.; Trayhurn, P. Dysregulation of the expression and secretion of inflammation-related adipokines by hypoxia in human adipocytes. Pflug. Arch. 2007, 455, 479–492.
- Pasarica, M.; Sereda, O.R.; Redman, L.M.; Albarado, D.C.; Hymel, D.T.; Roan, L.E.; Rood, J.C.; Burk, D.H.; Smith, S.R. Reduced adipose tissue oxygenation in human obesity: Evidence for rarefaction, macrophage chemotaxis, and inflammation without an angiogenic response. Diabetes 2009, 58, 718–725.
- Wadey, R.M.; Connolly, K.D.; Mathew, D.; Walters, G.; Rees, D.A.; James, P.E. Inflammatory adipocyte-derived extracellular vesicles promote leukocyte attachment to vascular endothelial cells. Atherosclerosis 2019, 283, 19–27.
- Lakka, H.M.; Laaksonen, D.E.; Lakka, T.A.; Niskanen, L.K.; Kumpusalo, E.; Tuomilehto, J.; Salonen, J.T. The metabolic syndrome and total and cardiovascular disease mortality in middle-aged men. JAMA 2002, 288, 2709–2716.
- Hall, J.E.; do Carmo, J.M.; da Silva, A.A.; Wang, Z.; Hall, M.E. Obesity-induced hypertension: Interaction of neurohumoral and renal mechanisms. Circ. Res. 2015, 116, 991–1006.
- Chandra, A.; Neeland, I.J.; Berry, J.D.; Ayers, C.R.; Rohatgi, A.; Das, S.R.; Khera, A.; McGuire, D.K.; de Lemos, J.A.; Turer, A.T. The relationship of body mass and fat distribution with incident hypertension: Observations from the Dallas Heart Study. J. Am. Coll. Cardiol. 2014, 64, 997–1002.
- Ouedraogo, R.; Gong, Y.; Berzins, B.; Wu, X.; Mahadev, K.; Hough, K.; Chan, L.; Goldstein, B.J.; Scalia, R. Adiponectin deficiency increases leukocyte-endothelium interactions via upregulation of endothelial cell adhesion molecules in vivo. J. Clin. Investig. 2007, 117, 1718–1726.
- Cao, Y.; Tao, L.; Yuan, Y.; Jiao, X.; Lau, W.B.; Wang, Y.; Christopher, T.; Lopez, B.; Chan, L.; Goldstein, B.; et al. Endothelial dysfunction in adiponectin deficiency and its mechanisms involved. J. Mol. Cell. Cardiol. 2009, 46, 413–419.
- Deng, G.; Long, Y.; Yu, Y.R.; Li, M.R. Adiponectin directly improves endothelial dysfunction in obese rats through the AMPK-eNOS Pathway. Int. J. Obes. 2010, 34, 165–171.
- Zhi, Z.; Pengfei, Z.; Xiaoyi, T.; Genshan, M. Adiponectin ameliorates angiotensin II-induced vascular endothelial damage. Cell Stress Chaperones 2014, 19, 705–713.
- Fang, F.; Liu, G.C.; Kim, C.; Yassa, R.; Zhou, J.; Scholey, J.W. Adiponectin attenuates angiotensin II-induced oxidative stress in renal tubular cells through AMPK and cAMP-Epac signal transduction pathways. Am. J. Physiol. Renal Physiol. 2013, 304, F1366–F1374.
- Lembo, G.; Vecchione, C.; Fratta, L.; Marino, G.; Trimarco, V.; d’Amati, G.; Trimarco, B. Leptin induces direct vasodilation through distinct endothelial mechanisms. Diabetes 2000, 49, 293–297.
- Mitchell, J.L.; Morgan, D.A.; Correia, M.L.; Mark, A.L.; Sivitz, W.I.; Haynes, W.G. Does leptin stimulate nitric oxide to oppose the effects of sympathetic activation? Hypertension 2001, 38, 1081–1086.
- Skurk, T.; van Harmelen, V.; Blum, W.F.; Hauner, H. Angiotensin II promotes leptin production in cultured human fat cells by an ERK1/2-dependent pathway. Obes. Res. 2005, 13, 969–973.
- Adamczak, M.; Kokot, F.; Wiecek, A.W. Relationship between plasma renin profile and leptinaemia in patients with essential hypertension. J. Hum. Hypertens. 2000, 14, 503–509.
- Bouloumie, A.; Marumo, T.; Lafontan, M.; Busse, R. Leptin induces oxidative stress in human endothelial cells. FASEB J. 1999, 13, 1231–1238.
- Erickson, L.A.; Fici, G.J.; Lund, J.E.; Boyle, T.P.; Polites, H.G.; Marotti, K.R. Development of venous occlusions in mice transgenic for the plasminogen activator inhibitor-1 gene. Nature 1990, 346, 74–76.
- Eren, M.; Painter, C.A.; Atkinson, J.B.; Declerck, P.J.; Vaughan, D.E. Age-dependent spontaneous coronary arterial thrombosis in transgenic mice that express a stable form of human plasminogen activator inhibitor-1. Circulation 2002, 106, 491–496.
- Watanabe, K.; Watanabe, R.; Konii, H.; Shirai, R.; Sato, K.; Matsuyama, T.A.; Ishibashi-Ueda, H.; Koba, S.; Kobayashi, Y.; Hirano, T.; et al. Counteractive effects of omentin-1 against atherogenesisdagger. Cardiovasc. Res. 2016, 110, 118–128.
- Niersmann, C.; Carstensen-Kirberg, M.; Maalmi, H.; Holleczek, B.; Roden, M.; Brenner, H.; Herder, C.; Schottker, B. Higher circulating omentin is associated with increased risk of primary cardiovascular events in individuals with diabetes. Diabetologia 2020, 63, 410–418.
- Jang, Y.; Lee, J.H.; Wang, Y.; Sweeney, G. Emerging clinical and experimental evidence for the role of lipocalin-2 in metabolic syndrome. Clin. Exp. Pharmacol. Physiol. 2012, 39, 194–199.
- Hemdahl, A.L.; Gabrielsen, A.; Zhu, C.; Eriksson, P.; Hedin, U.; Kastrup, J.; Thoren, P.; Hansson, G.K. Expression of neutrophil gelatinase-associated lipocalin in atherosclerosis and myocardial infarction. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 136–142.


