Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Pavel Horky | + 2892 word(s) | 2892 | 2021-11-08 10:37:18 | | | |
| 2 | Vivi Li | Meta information modification | 2892 | 2021-11-17 02:41:47 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Horky, P. Selenium Nanoparticles in the Plant Production. Encyclopedia. Available online: https://encyclopedia.pub/entry/16057 (accessed on 25 July 2026).
Horky P. Selenium Nanoparticles in the Plant Production. Encyclopedia. Available at: https://encyclopedia.pub/entry/16057. Accessed July 25, 2026.
Horky, Pavel. "Selenium Nanoparticles in the Plant Production" Encyclopedia, https://encyclopedia.pub/entry/16057 (accessed July 25, 2026).
Horky, P. (2021, November 16). Selenium Nanoparticles in the Plant Production. In Encyclopedia. https://encyclopedia.pub/entry/16057
Horky, Pavel. "Selenium Nanoparticles in the Plant Production." Encyclopedia. Web. 16 November, 2021.
Copy Citation
Selenium (Se) is a common trace metalloid found in the Earth’s crust. In 1817, chemist Jacob Berzelius isolated it for the first time, and since then, it has been known for its properties. Recent research has shown that treatment with Se at low concentrations has a beneficial influence on plant development and yield. Se may work as an essential factor by interfering with a several of physiological processes. It is a remarkable antioxidant and pro-oxidant agent of plants that helps to cope with a variety of abiotic stresses, such as salinity, drought, intense temperature fluctuations, toxic metals/metalloids, and other environmental pollutants and toxins.
selenium
nanoparticles
plant nutrition
agriculture
1. Selenium in Plants
Se is abundant, thus the amount of Se present in plants is proportional to the amount in the surrounding soil [1]. Many plants accumulate Se to the point where it becomes poisonous if consumed by livestock. Plants that accumulate massive volumes of Se are frequently found in areas with high Se concentration in the soil. The summarization of the main classes of Se utilizing plants is given in Figure 1. The plants which can accumulate larger quantities of Se are called indicator plants. Some Astragalus species, as well as prince’s plume and some woody asters, are considered as indicator plants (Figure 1A). The indicator plants can accumulate up to 3000 parts per million (ppm) of Se in their different parts including roots, shoots, and leaves. Some plants they can accumulate up to 50 ppm Se [2]. The secondary Se accumulators are some native ranges of plants and crop plants, including wheat, barley, alfalfa, and western wheatgrass, respectively (Figure 1B). The physiological conditions of the plant, as well as the species of the plant, influence how the Se is taken up and dispersed by its root [3]. Aerial silks are typically composed of approximately 80% selenite and 65% selenate by weight [1]. Forages contain Se in concentrations ranging from 0.2 to 0.6 ppm, putting livestock at threat of Se poisoning. Se in wheat plants’ seeds is normally stored as selenomethionine (SeMet), with varying levels of Se stored depending on the environment [4]. Compared to other minerals, plants mostly absorb selenite in higher concentrations. Considering that selenate and selenite have chemical characteristics, they both undergo the same metabolic pathway [1]. There is a strong correlation between Se deficiency in soil and the appearance of disease symptoms associated with low Se intake in humans and other mammals. For example, soil’s Se concentrations range from 8000 mg kg−1 in soils of Russia to 0.005 mg kg1 in soils of China and Finland. As a result, various efforts have been made to enhance agricultural food production with Se through fertilization, genetic breeding, or in biofortification strategy [5].

Figure 1. Images of plants containing Se: (A) indicator plants and (B) facultative Se-absorbing plants.
1.1. Effects of Selenium in Plants
Many studies have been conducted to determine the role of Se in plants. The issue of Se utilization of higher plants is still up in the air and this question has not been resolved. There are, however, some answers [6]. Some positive and negative effects of Se are discussed below and summarized in Figure 2.
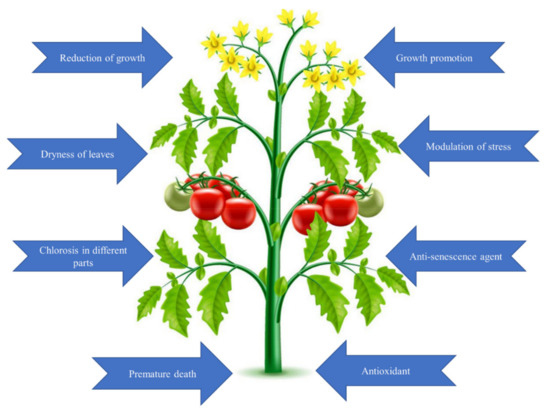
Figure 2. Demonstration of useful and harmful effects of Se on plant physiology.
1.1.1. Beneficial Effects of Selenium in Plants
Many studies have shown that Se, even at low concentrations, can stimulate plant growth. It was found that 5 µg of Se boosted a root growth and increased relative water content in hot pepper plants by 13% when compared to control plants [7]. Furthermore, both 3 and 5 µM concentration of Se triggered a 25% spike in leaf area, which resulted in an increase in the plant’s overall growth and biomass. Moreover, the growth of rice was reported to be stimulated by using a low dose of Se. In 2019, Hemmati et al. suggested that soil Se fertilization can be an accurate and efficient technique for improving the overall performance of plants [8]. Last year, Rady et al. (2020) confirmed that tomatoes treated with 40 M of Se established increased drought tolerance, which was accompanied by actions of several antioxidant enzymes, including ascorbate peroxidase (APX) by 44%, superoxide dismutase (SOD) by 56%, and catalase (CAT) by 57%, respectively [9]. The application of Se also increased accumulation of carbohydrates in the young leaf surfaces of potatoes.
Salinity harms agricultural production because it inhibits plants growth and yield. There have been many experiments conducted to investigate the crucial role of Se in protecting plants against salt-induced stress. Another study investigated the role of 1–25 mM of sodium selenite (Na2SeO3) in regulating salinity tolerance in maize by measuring its concentration in the plant [10]. Under salinity, their findings revealed that 1 mM Se increased plant growth and development, photosynthetic rate, and K+ content, while the ratio of Na+ was deceased. Moreover, the Se supplementation (20 mg L−1) improved the growth and chlorophyll content of maize by reducing oxidative damage because of high malondialdehyde (MDA) and hydrogen peroxide (H2O2) levels under high-salinity stress [11]. A significant finding was that supplementation of Se enhanced the salinity tolerance of maize at the reproductive phase more than at the vegetative stage. Several experiments were conducted to determine the significant function of Se for enhancing plant drought tolerance and drought resistance [12]. Abiotic stress is a significant constraint on the productivity of modern agriculture. Therefore, researchers are attempting to develop modern tactics that will be effective in dealing with plant stress. Exogenous stress protectants are becoming increasingly popular as a means of increasing stress tolerance [12]. Besides improving the photosynthetic activity, increasing the direct quenching of ROS, and upregulating both the enzymatic and nonenzymatic parts of the antioxidant defense system, Se can helps to decrease plant cell membrane damage. Noticeably, at significant amounts, Se tends to increase lipid peroxidation and α-tocopherol levels, but also, in particular, glutathione peroxidase (GPx) activity, which ultimately results in cell death. This contributes to the systemic defense against the tissue-damaging effects of ROS [13].
1.1.2. Harmful Effects of Selenium in Plants
Although Se has a biofortifying effect on plants, when given to them in high concentrations, it could have also toxic effects on plant growth. It was found that a plant growth is negatively impacted by the alteration of S for Se in cysteine and methionine amino acids, which results in alteration of disulfide bonds in proteins, altering the structure and behavior of proteins, and having a devastating effect on overall growth [14][15]. Besides that, another damaging effect of Se during its incorporation into organic compounds is the depletion of glutathione, a nonenzymatic antioxidant that protects plants against ROS. Because of this asymmetry between detoxification and the creation of ROS, a significant oxidative burst occurs, with a resulting reduction in plant growth. It has been revealed by previous studies that plants’ mineral balance is disrupted by a surplus of Se, which causes adverse changes in the quantifiable composition of essential nutrients [5]. Se affects a variety of biochemical processes and physiological processes, including development, photosynthesis, respiration, gas exchange, water holding capacity, phloem unloading, and initiation of protease inhibitor genes, by altering the uptake, aggregation, and transit of mineral nutrients. For example, Na+ interferes with nutrients in plant tissues, which causes disturbance of various physiological events controlled by Se [10]. It is the quantitative proportions of Se and essential elements that determine the interaction between them, resulting in antagonistic and synergistic effects. Se ions can also alter the permeability coefficients of several ions in biomembranes, affecting the transport of those ions through the membrane [16]. Molnár et al. (2018) demonstrated that selenite-induced cell wall alterations and stomatal regulations in Arabidopsis were associated with lower stomatal density and that selenite sensitivity resulted in the stomatal opening, callose aggregation, serious oxidative stress, and moderate nitrosative modifications in plants. The Se-induced photosynthesis dysfunction is widely regarded as the most important cause of increased accumulation of ROS and oxidative stress, and it is widely recognized as one of the most important mechanisms of Se phytotoxicity [17].
The plant proteomes are targeted by Se toxicity, as was recently shown by Kolbert et al. (2019). An excess of Se in plants harms a variety of physiological and biochemical processes. One of the most significant negative consequence is a reduction in chlorophyll biosynthesis, which results in chlorosis. Maize handled with selenite (5–100 M) has shown enhanced levels of phosphorus (P) and calcium (Ca+) content, but decreased potassium (K) content. Tall fescue and white clover that had been treated with hydrogen selenate showed increased Ca+ bioconcentration and a decrease in P concentration in the opposite direction [18]. Se toxicity is unquestionably a result of the accumulation of excess Se within the plant cell and the competition among Se and S for the addition of structural components or involvement in biochemical reactions due to the chemical structural similarity between them. Because of Se toxicity, not only are seleno- and oxyproteins formed, but it has been discovered that nitroproteins have also been created. Using specific plants, Se-induced proteomic damage can be reduced by diverting protein synthesis into other pathways [16]. Furthermore, proteasomes can remove nitroproteins, selenoproteins, and oxyproteins that have been damaged or malformed. Another researcher showed that lettuce growth was stimulated when the hydrogen selenate content was low [19]. A significant reduction in lettuce yield was observed when the Se concentration in the shoots exceeded 20 mg kg−1 dry weight. Furthermore, Se exposure had a variety of effects on the morphology of the roots [16], which could be harmful for the plant. In a separate study, the effect of Se on lettuce was investigated, and the results revealed increased Se concentrations in the shoots, but decreased macronutrient accumulation in the leaves of lettuce, as well as growth reduction symptoms.
2. Agricultural Use of Nano SeNPs
2.1. Fertilizer for Crops
In terms of the application of SeNPs in the Se fertilization sector, fertilizers are one of the most important factors in increasing crop yields and agricultural productivity, as well as ensuring food security in developing countries [16]. With the progress of innovative, cutting-edge, and developing technologies such as nanotechnology, the implementation of SeNPs as an adjunct to standard Se fertilizers to improve crops has emerged as a viable option to conventional Se fertilizers.
The soil is a biodegradable material, and SeNPs are used to stimulate the soil organic matter. It has been shown that the humic substance, which is delivered synchronously with SeNPs and other NPs, handles the stimulation effect as well [20]. Microbial communities control organic matter degradation, which has a significant impact on soil fertility. Within specific environments, bacteria are not a faceless mixture of once-gained participants, but a structured, strictly-ordered polymicrobial community in which each participant has a specific functional role. As a bioreactor, the soil speeds up a wide range of biodegradation processes [21]. In terms of biological processes and yield in the soil and plants, the application of Se in standard fertilizer form is deemed less efficient than the application of SeNPs in standard fertilizer form [22]. Specifically, it has been found that Se can ease stress in plants because it causes the manufacture of secondary metabolites and increases the activities of antioxidant enzymes according to the literature [17]. To mitigate multiple kinds of abiotic stress, including extremely high temperatures, droughts, heavy metal accumulation, and salt, the usage of SeNPs is increasingly popular and is becoming increasingly important. The bean plant is used for soil fertility analysis, which enhances plant development. The plant has been evaluated and analyzed for chlorophyll and protein content. Plants showed effective growth at high concentrations of SeNPs [23]. A study by Hebat-Allah et al. (2019) showed that SeNPs impacted the growth of groundnut cultivars by altering photosynthetic pigments, lipid peroxidation, antioxidant enzymes (ascorbic acid peroxidase, catalase, peroxidase), total soluble sugars, phenol content, and total flavonoids in the plants. Tolerance to sandy soil conditions was improved when SeNPs were used as a stimulant and/or a stressor [24]. As an example, handling with SeNPs at a ratio of 100 mg/L in barley (Hordeum vulgare) crops cultivated under saline stress led to a direct accumulation of Se in leaves, an improvement in the amount of aggregate phenolic composites, and a decrease in the content of ROS-mediated cellular membrane harm markers, including such MDA, which may affect metabolism and be responsible for nutrient deficiencies [25]. In the study conducted on tobacco plants it was shown that SeNPs had no positive effect on the number of tobacco shoots, whereas 50 mg L−1 selenate completely inhibited the expansion of tobacco shoots. Increasing the concentrations of SeNPs significantly increased the rate of roots regeneration. With the use of 50–100 mg L−1 SeNPs, the roots were far more extensive and densely packed, and the fresh weight increased significantly as well. On the contrary, selenate completely inhibited the formation of roots at concentrations ranging from 50 to 100 mg L−1 [24]. In plant tissue culture, the biological activities of SeNPs were distinct from those of the selenate ion (SeO32-). It was showed that SeNPs concentrations ranging from 50 to 100 mg kg−1 significantly increased organogenesis and root system growth (>40%), whereas selenate had no such effects at any intensity. SeNPs concentration ranging from 50 to 100 mg kg−1 strongly prevented both callus development and root regeneration. This previous concentration was effective in stimulating not only roots initiation or roots elongation, but biomass production as well [24].
Another point worth mentioning is that SeNPs have been shown to reduce the concentrations of heavy metals throughout plant tissues, which are extremely toxic to the organism. It was discovered in most of the studies that Se is more of an adversary of noxious elements than lead (Pb) as well as cadmium (Cd); as a result, implementing SeNPs has been shown to significantly lower the concentrations of both elements in the environment, dropping the negative effects of these toxic substances on the plant’s environment. After the usage of SeNPs in female spinach plants, it was discovered that the Cd and Pb concentrations decreased by 66% and 19%, respectively. The protective effect of female spinach plants against Cd increases as the oxidation state of the plant decreases [26]. To date, the nanofertilizers are manufactured using metal ions such as silicone and Se. SeNPs are a potent low-dose stimulant with a noticeable effect.
2.2. Biofortification
The potential for gradual Se release from SeNPs for biofortification of plant foods has piqued the curiosity of researchers interested in using them in agroecosystems to minimize potential losses that can occur when commercial fertilizers are used. Therefore, SeNPs may be employed in biofortification, which aims to raise the Se content of comestible parts of plants to prevent Se insufficiency in humans and livestock because of this. SeNPs have also been proven to be less harmful to plants than ionic Se salts (SeO4−2 and SeO3−2), as shown in investigations of Nicotiana tabacum and Allium sativum [27]. It was described that the uptake of SeNPs was 1.7 times more decreased than the uptake ratio of SeO4−2 and SeO3−2 and that SeNPs were then absorbed into organic forms, including SeMet, which accumulated mostly in the root cell walls of the plants [28]. Furthermore, SeNPs generated by chemical means were more capably absorbed than SeNPs manufactured via biological approaches. For example, it has been published that wheat roots handled with 40 nm SeNPs, attained through chemical synthesis, absorbed 1.8 and 2.2 times more Se than wheat roots preserved to 140 and 240 nm SeNPs, respectively. This shows the importance of particulate type and amount of NP synthesis in this type of application [29]. Recently, there has been amplified attention in the practice of SeNPs for the biofortification of crops, owing to their ability to improve the quality, nutritional characteristics, and amount of Se available in the edible sections of plants. When compared to other regularly used Se sources, the consumption of plant foodstuffs biofortified with SeNPs may have an entirely different effect on the human body, depending on parameters such as the diameter of the NPs, the processing methods, and the surface makeup of the NPs [29]. As a result, a great number of in vivo experiments must be carried out to determine whether there are any harmful effects associated with the utilization of plant foods biofortified with SeNPs. Because of this, it is impossible to make a broad generalization about the recommended doses for the intake of foods biofortified using SeNPs [25].
2.3. Effect of SeNPs on Germination
Because of their ability to influence seed growth and sowing characteristics, SeNPs are an asset in agriculture. Plants grow more resilient to drought, disease, and pests when they are exposed to their effects [30]. It has been shown in studies that SeNPs have a substantial impact on seedling germination and development early in the ontogenesis process. There may be a dose–response relationship between NPs and plant development [31]. A higher concentration of NPs, for example, has been found to slow down seedling development when compared to controls. The impact of SeNPs on the germination characteristics of Hordeum vulgare L. seeds was investigated. Researchers found that SeNPs increased the length of shoots and roots, as well as the germination rate [32]. The sample treated with SeNPs preparation at a dosage of 4.65 g mL−1 had the greatest seed germination percentage [30]. The study’s findings suggest that SeNPs may serve as a seed-based supply of the microelement Se. There is less toxicity in the production of SeNPs in this research compared to Se in ionic form; thus, it may be utilized to repair biochemical processes and replenish Se in germinating seeds [32]. It is essential to investigate the effects of SeNPs preparation and to investigate the effects of SeNPs on the germination characteristics of other common crops such as maize, rice, and soybeans [30]. SeNPs also promote root development and organogenesis. In lettuce, ryegrass, Brassica oleracea, and potato plants, trace quantities of Se have been shown to increase growth [33].
References
- Wrobel, K.; Esperanza, M.G.; Barrientos, E.Y.; Escobosa, A.R.C. Different approaches in metabolomic analysis of plants exposed to selenium: A comprehensive review. Acta Physiol. Plant. 2020, 42, 125.
- Ralphs, M.H. Ecological relationships between poisonous plants and rangeland condition: A review. J. Range Manag. 2002, 55, 285–290.
- Rizwan, M.; Ali, S.; Rehman, M.Z.U.; Rinklebe, J.; Tsang, D.C.W.; Tack, F.M.G.; Abbasi, G.H.; Hussain, A.; Igalavithana, A.D.; Lee, B.C.; et al. Effects of selenium on the uptake of toxic trace elements by crop plants: A review. Crit. Rev. Environ. Sci. Technol. 2021, 51, 2531–2566.
- Kieliszek, M.; Bano, I.; Zare, H. A Comprehensive Review on Selenium and Its Effects on Human Health and Distribution in Middle Eastern Countries. Biol. Trace Elem. Res. 2021.
- Mora, M.L.; Duran, P.; Acuna, A.J.; Cartes, P.; Demanet, R.; Gianfreda, L. Improving selenium status in plant nutrition and quality. J. Soil Sci. Plant Nutr. 2015, 15, 486–503.
- Brodowska, M.S.; Kurzyna-Szklarek, M.; Haliniarz, M. Selenium in the Environment. J. Elem. 2016, 21, 1173–1185.
- Mozafariyan, M.; Shekari, L.; Hawrylak-Nowak, B.; Kamelmanesh, M.M. Protective Role of Selenium on Pepper Exposed to Cadmium Stress During Reproductive Stage. Biol. Trace Elem. Res. 2014, 160, 97–107.
- Hasanuzzaman, M.; Bhuyan, M.; Raza, A.; Hawrylak-Nowak, B.; Matraszek-Gawron, R.; Al Mahmud, J.; Nahar, K.; Fujita, M. Selenium in plants: Boon or bane? Environ. Exp. Bot. 2020, 178, 104170.
- Rady, M.M.; Belal, H.E.E.; Gadallah, F.M.; Semida, W.M. Selenium application in two methods promotes drought tolerance in Solanum lycopersicum plant by inducing the antioxidant defense system. Sci. Hortic. 2020, 266, 109290.
- Subramanyam, K.; Du Laing, G.; Van Damme, E.J.M. Sodium Selenate Treatment Using a Combination of Seed Priming and Foliar Spray Alleviates Salinity Stress in Rice. Front. Plant Sci. 2019, 10, 116.
- Jozwiak, W.; Politycka, B. Effect of Selenium on Alleviating Oxidative Stress Caused by a Water Deficit in Cucumber Roots. Plants 2019, 8, 217.
- Agbolade, J.O.; David, O.; Ajiboye, A.; Kioko, J.; Jolayemi, O.; Olawuni, I.; Ojo, M.; Akomolafe, G.; Adekoya, M.; Komolafe, R. Morpho-physiological effect of selenium on salinity-stressed wheat (Triticum aestivum L.). J. Biol. Res. 2019, 92, 7650.
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive Oxygen Species and Antioxidant Defense in Plants under Abiotic Stress: Revisiting the Crucial Role of a Universal Defense Regulator. Antioxidants 2020, 9, 681.
- Ferreira, R.L.D.; Prado, R.D.; de Souza, J.P.; Gratao, P.L.; Tezotto, T.; Cruz, F.J.R. Oxidative Stress, Nutritional Disorders, and Gas Exchange in Lettuce Plants Subjected to Two Selenium Sources. J. Soil Sci. Plant Nutr. 2020, 20, 1215–1228.
- Pilon-Smits, E.A.H. On the Ecology of Selenium Accumulation in Plants. Plants 2019, 8, 197.
- Kaur, N.; Sharma, S.; Kaur, S.; Nayyar, H. Selenium in agriculture: A nutrient or contaminant for crops? Arch. Agron. Soil Sci. 2014, 60, 1593–1624.
- Molnar, A.; Kolbert, Z.; Keri, K.; Feigl, G.; Ordog, A.; Szollosi, R.; Erdei, L. Selenite-induced nitro-oxidative stress processes in Arabidopsis thaliana and Brassica juncea. Ecotoxicol. Environ. Saf. 2018, 148, 664–674.
- Naseem, M.; Anwar-ul-Haq, M.; Wang, X.K.; Farooq, N.; Awais, M.; Sattar, H.; Malik, H.A.; Mustafa, A.; Ahmad, J.; El-Esawi, M.A. Influence of Selenium on Growth, Physiology, and Antioxidant Responses in Maize Varies in a Dose-Dependent Manner. J. Food Qual. 2021, 2021, 6642018.
- Husen, A.; Siddiqi, K.S. Plants and microbes assisted selenium nanoparticles: Characterization and application. J. Nanobiotechnol. 2014, 12, 28.
- Gudkov, S.V.; Shafeev, G.A.; Glinushkin, A.P.; Shkirin, A.V.; Barmina, E.V.; Rakov, I.I.; Simakin, A.V.; Kislov, A.V.; Astashev, M.E.; Vodeneev, V.A.; et al. Production and Use of Selenium Nanoparticles as Fertilizers. ACS Omega 2020, 5, 17767–17774.
- Rajput, V.D.; Minkina, T.; Feizi, M.; Kumari, A.; Khan, M.; Mandzhieva, S.; Sushkova, S.; El-Ramady, H.; Verma, K.K.; Singh, A.; et al. Effects of Silicon and Silicon-Based Nanoparticles on Rhizosphere Microbiome, Plant Stress and Growth. Biology 2021, 10, 791.
- Jain, R.; Seder-Colomina, M.; Jordan, N.; Dessi, P.; Cosmidis, J.; van Hullebusch, E.D.; Weiss, S.; Farges, F.; Lens, P.N.L. Entrapped elemental selenium nanoparticles affect physicochemical properties of selenium fed activated sludge. J. Hazard. Mater. 2015, 295, 193–200.
- Mikula, K.; Izydorczyk, G.; Skrzypczak, D.; Mironiuk, M.; Moustakas, K.; Witek-Krowiak, A.; Chojnacka, K. Controlled release micronutrient fertilizers for precision agriculture—A review. Sci. Total Environ. 2020, 712, 136365.
- El-Ramady, H.; Abdalla, N.; Taha, H.S.; Alshaal, T.; El-Henawy, A.; Faizy, S.; Shams, M.S.; Youssef, S.M.; Shalaby, T.; Bayoumi, Y.; et al. Selenium and nano-selenium in plant nutrition. Environ. Chem. Lett. 2016, 14, 123–147.
- Garza-Garcia, J.J.O.; Hernandez-Diaz, J.A.; Zamudio-Ojeda, A.; Leon-Morales, J.M.; Guerrero-Guzman, A.; Sanchez-Chipres, D.R.; Lopez-Velazquez, J.C.; Garcia-Morales, S. The Role of Selenium Nanoparticles in Agriculture and Food Technology. Biol. Trace Elem. Res. 2021.
- Golubkina, N.A.; Folmanis, G.E.; Tananaev, I.G.; Krivenkov, L.V.; Kosheleva, O.V.; Soldatenko, A.V. Comparative Evaluation of Spinach Biofortification with Selenium Nanoparticles and Ionic Forms of the Element. Nanotechnol. Russ. 2017, 12, 569–576.
- Li, Y.X.; Zhu, N.L.; Liang, X.J.; Zheng, L.R.; Zhang, C.X.; Li, Y.F.; Zhang, Z.Y.; Gao, Y.X.; Zhao, J.T. A comparative study on the accumulation, translocation and transformation of selenite, selenate, and SeNPs in a hydroponic-plant system. Ecotoxicol. Environ. Saf. 2020, 189, 109955.
- Golubkina, N.A.; Folmanis, G.E.; Tananaev, I.G. Comparative evaluation of selenium accumulation by allium species after foliar application of selenium nanoparticles, sodium selenite and sodium selenate. Dokl. Biol. Sci. 2012, 444, 176–179.
- Wang, K.; Wang, Y.Q.; Li, K.; Wan, Y.N.; Wang, Q.; Zhuang, Z.; Guo, Y.B.; Li, H.F. Uptake, translocation and biotransformation of selenium nanoparticles in rice seedlings (Oryza sativa L.). J. Nanobiotechnology 2020, 18, 103.
- Siddiqui, S.A.; Blinov, A.V.; Serov, A.V.; Gvozdenko, A.A.; Kravtsov, A.A.; Nagdalian, A.A.; Raffa, V.V.; Maglakelidze, D.G.; Blinova, A.A.; Kobina, A.V.; et al. Effect of Selenium Nanoparticles on Germination of Hordeum Vulgare Barley Seeds. Coatings 2021, 11, 862.
- Malagoli, M.; Schiavon, M.; dall’Acqua, S.; Pilon-Smits, E.A.H. Effects of selenium biofortification on crop nutritional quality. Front. Plant Sci. 2015, 6, 280.
- Ikram, M.; Raja, N.I.; Javed, B.; Mashwani, Z.U.R.; Hussain, M.; Ehsan, M.; Rafique, N.; Malik, K.; Sultana, T.; Akram, A. Foliar applications of bio-fabricated selenium nanoparticles to improve the growth of wheat plants under drought stress. Green Process. Synth. 2020, 9, 706–714.
- Bideshki, A.; Arvin, M.J.; Aien, A.; Hasandokht, M.R.; Khalighi, A. Interactive effects of Foliar 24-Epibrassinolide and selenium applications on yield, reduce nitrate accumulation and selenium enrichment in potato tuber in field. Cogent Food Agric. 2019, 5, 1690315.
More
Information
Subjects:
Agriculture, Dairy & Animal Science
Contributor
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.4K
Revisions:
2 times
(View History)
Update Date:
17 Nov 2021
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No