Aptamers are single-stranded nucleic acids with defined tertiary structures for selective binding to target molecules by utilizing complementary shape interactions. The secondary structures of aptamers can be predicted from their sequences
[1]. Many programs have been developed to identify and study aptamer binding motifs, including AptaTRACE
[2] for identifying sequence-structure binding motifs in high-throughput systematic evolution of ligands by exponential enrichment (HT-SELEX) data. Aptamotif
[3] and APTANI
[4] also identified sequence-structure motifs in SELEX-derived aptamers using an ensemble-based approach. As only a small fraction of the sequence is involved in direct contact with the target, aptamer truncation can be used to find the minimum sequence while maintaining selective binding activity. Ultimately, aptamer truncation will help to reduce the material cost of aptamer synthesis, enable material quality assurance and prevent unexpected toxicity
[5][6][7][8]. Many studies have demonstrated effective aptamer truncations in different ways. Truncations on the stem-loop regions showed a significant increase in the binding affinity for the VEGF-165
[9]. Aptamer acquisition from a random region has shown successful identification of 15 nt
[10] or 20 nt
[11] aptamers. Moreover, a 14 nucleotides aptamer was reported as the smallest functional unit of the transferrin receptor aptamer with enhanced binding affinity to the transferrin receptor
[12].
Vimentin is an extracellular matrix protein that is part of the intermediate filament protein family
[13]. Overexpression of vimentin may be associated with increased metastatic capacity through the epithelial to mesenchymal transformation (EMT) of ovarian tumor cells
[14]. Studies have shown that increased expression of vimentin correlates to decreased survival rate in a variety of cancers such as colorectal, cervical, breast, gastric, and non-small cell lung cancers, to name a few
[15][16][17]. There are limited studies, however, exploring the relationship between vimentin expression and ovarian tumor prognosis. A recent study by Szubert et al.
[18] showed a prolonged overall survival rate with an increased rate of vimentin expression in the ovarian tumor cells. While this relationship is contrary to that seen in other tumor pathologies, it remains evident that a relationship exists between vimentin and overall tumor prognosis, and a marker for vimentin would serve as an invaluable tool in assessing tumor prognosis, as well as potentially serving as a marker for targeted molecular imaging. Currently, there are aptamers against vimentin for isolation of circulating tumor cells undergoing epithelial mesenchymal transition
[19]. Zamay et al. used aptamer NAS-24 which binds to vimentin for intracellular targeting of mouse ascites adenocarcinoma cells in vitro and in vivo
[20]. We have identified two phosphorothioated aptamers, thioaptamers, V3 and V5 that have high affinity and specificity binding to vimentin through our innovative morphology-based tissue aptamer selection (Morph-X-Select) method
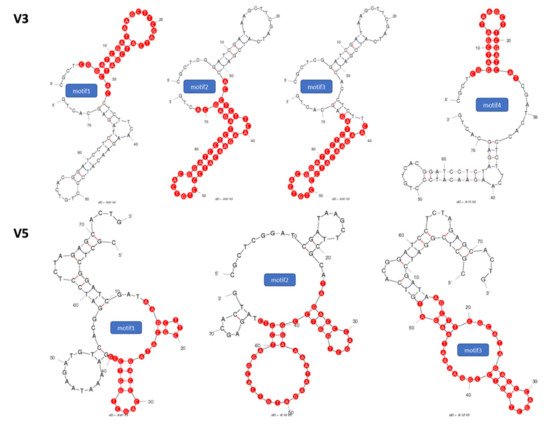
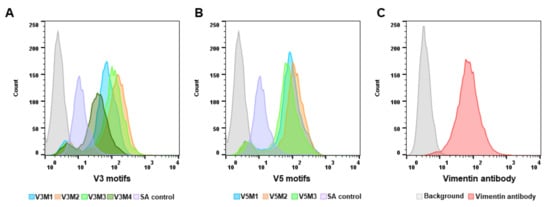
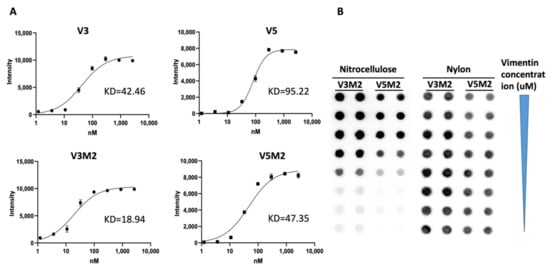
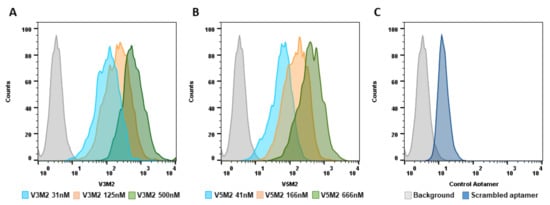
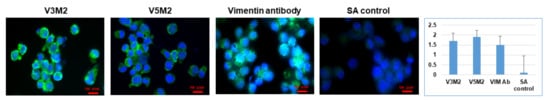
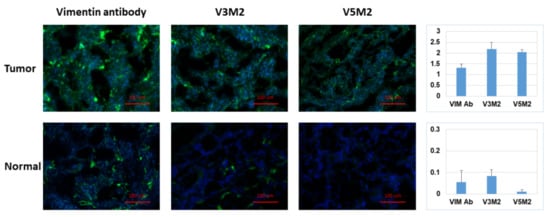
[21]. The secondary structure prediction suggests that those aptamers would form 3–4 stem-loop structures. The originally selected V3 and V5 aptamers were 74-mer in length and had high affinities to vimentin (Kd = 42.46 nM, Kd = 95.22 nM, respectively). As the recognition-based binding activity between the DNA aptamer and vimentin protein is dependent on the secondary structure of the vimentin aptamer, we further analyzed predictive secondary structures of the aptamers to identify sequence features and patterns by Mfold
[1], and conducted a few truncations on the stem-loop regions to achieve improved binding affinity with the truncated aptamers.
 +1 credit
+1 credit