Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Maryam Nasiri Aghdam | + 3648 word(s) | 3648 | 2021-10-19 04:44:09 | | | |
| 2 | Peter Tang | Meta information modification | 3648 | 2021-10-25 04:03:08 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Nasiri Aghdam, M. CELF Family Proteins in Cancer. Encyclopedia. Available online: https://encyclopedia.pub/entry/15307 (accessed on 26 July 2026).
Nasiri Aghdam M. CELF Family Proteins in Cancer. Encyclopedia. Available at: https://encyclopedia.pub/entry/15307. Accessed July 26, 2026.
Nasiri Aghdam, Maryam. "CELF Family Proteins in Cancer" Encyclopedia, https://encyclopedia.pub/entry/15307 (accessed July 26, 2026).
Nasiri Aghdam, M. (2021, October 22). CELF Family Proteins in Cancer. In Encyclopedia. https://encyclopedia.pub/entry/15307
Nasiri Aghdam, Maryam. "CELF Family Proteins in Cancer." Encyclopedia. Web. 22 October, 2021.
Copy Citation
CELF (CUGBP Elav-like family) proteins are RBPs (RNA-binding proteins) with pleiotropic capabilities in RNA processing. Their responsibilities extend from alternative splicing and transcript editing in the nucleus to mRNA stability, and translation into the cytoplasm. In this way, CELF family members have been connected to global alterations in cancer proliferation and invasion, leading to their identification as potential tumor suppressors or even oncogenes. Notably, genetic variants, alternative splicing, phosphorylation, acetylation, subcellular distribution, competition with other RBPs, and ultimately lncRNAs, miRNAs, and circRNAs all impact CELF regulation.
CELF proteins
RNA-binding protein
noncoding RNA
cancer
1. Introduction
RNA-binding proteins (RBPs) orchestrate crucial aspects of RNA biology in many sets of targets. Global post-transcriptional modifications through RBPs, which are present in almost all organelles of a cell, can exert rapid cellular effects in response to external or internal stimuli. The widespread functions of RBPs range from splicing, localization, modification, stability, and translation of coding and noncoding RNAs [1]. In human cells, it is expected that approximately 1500 RBPs associate with over 33 million interactions in 3′UTR binding sites [2]. RBPs’ multifunctionality, their broad number of targets and subcellular location, as well as their tight interactions with other RBPs and noncoding RNAs, make them a challenging field to research, especially in cancer, where modifications in the expression, mutations, and copy number variations of RBPs can have a significant impact on their performance [3][4].
The CUGBP, ELAV-like family (CELF) of proteins are a kind of RBP that features cytoplasmic and nucleolar distribution and, like most other RBPs, they have broad and diverse roles in RNA regulation. The expression of CELF1 and CELF2 is detected in almost all tissues; however; their patterns can change in different developmental and differentiation stages. CELF3-6 are mostly present in neurons, and CELF6 can also be found in the kidneys and the testes [5]. All six members of this family feature three RRMs (RNA Recognition Motifs) with unique divergent linker domains, which let them attach to downstream targets [6]. CELF proteins are involved in RNA splicing through binding to the upstream or downstream intron of an alternative exon to mediate exon skipping or inclusion. They are also abundant in the 3′UTR of transcripts associating with mRNA decay or translation [7][8].
Non-coding RNAs of various kinds can directly or indirectly target CELF proteins; additionally, these RBPs can significantly alter non-coding RNAs in many ways, forming the noncoding RNA/CELFs regulatory axis. Long noncoding RNAs (lncRNAs), micro RNAs (miRNAs), and circular RNAs (circRNAs) are three forms of noncoding RNAs whose expression may be modified, and that can either promote or prevent carcinogenesis. LncRNAs with a length of longer than 200 nucleotides are transcribed by RNA polymerase II and preferentially stay in the nucleus to perform the regulation of gene expression and splicing [9]. MiRNAs are typically 20–23 nucleotides long, with the seed region for base pairing to target mRNAs located in seven or eight nucleotides at the 5′ end of the miRNA. The expression of a target mRNA is expected to bear a modest or strong reduction upon the binding of one miRNA or multiple miRNAs, respectively [10]. Lastly, circRNAs can be found broadly in almost all cells and tissues; they are mainly distributed in the cytoplasm and, rarely, in the nucleus. They can function as RNA sponges, nucleators of multiprotein complexes, or even transcriptional regulators [9].
2. General Characteristics of the CELF Protein Family
So far, six CELF RBPs have been discovered, with a nomenclature ranging from CELF1 to CELF6. Based on their phylogenetic structure and expression pattern, these members may be divided into two subfamilies: CELF1-2 and CELF3-6. CELF1 and CELF2 are extensively expressed in numerous tissues throughout the human body, but CELF3 through 5 are mostly found in the neurological system. CELF6 is most often found in the nervous system, the kidneys, and the testes [5][11][12]. All six members have a similar structure that includes three RRMs (RNA Recognition Motifs), two in the N-terminal and one in the C-terminal, as well as a divergent linker region that can improve RBP binding affinity through conformational variations. The nuclear localization signal, which comprises a lysine/arginine-rich region in the C terminal and a region important for nuclear export in the linker domain, are two other key structures [6][13][14]. A serine/threonine-rich phosphorylation area within the CELF1 and CELF2 linker domains could change their affinity for binding to target mRNAs [15][16]. However, there are no reports regarding the phosphorylation of CELF2 at this site. Apart from phosphorylation, the acetylation of the lysine436 residue inside the third RRM motif has been shown to influence CELF protein–protein interactions and mRNA affinity [17][18]. There are few studies in relation to the evolution of the CELF family members; however, their presence in xenopus, zebrafish, and mammals is clear evidence of their ontological importance and their conservation. During embryonic development CELF1 localizes in the vegetal pole of zebrafish and CELF2 in the animal pole [19][20]. This compartmentalization suggests that each protein performs different functions and, therefore, that they must have separated very early in evolutionary terms. Future works about the evolution and function of CELF proteins would clarify how and why these proteins arose.
By generating different isoforms, the alternative splicing process modifies the structure, expression, cellular location, and functional properties of CELF family members. For example, CELF2 features three promoters that create three distinct isoforms, each of which features a unique N-terminal sequence and 5′UTR [21]. CELF2 loses its RNA binding capacity when exon 14 is skipped; moreover, exon 9 skipping is reported for CELF4 [21][22]. Alternative splicing on 3′UTRs and C termini of CELF2 and CELF5 may affect their regulation and function [6]. Figure 1 illustrates the structure and conservation of all six proteins of the CELF family. Reported phosphorylation and acetylation sites are obtained from PhosphoSitePlus [23].
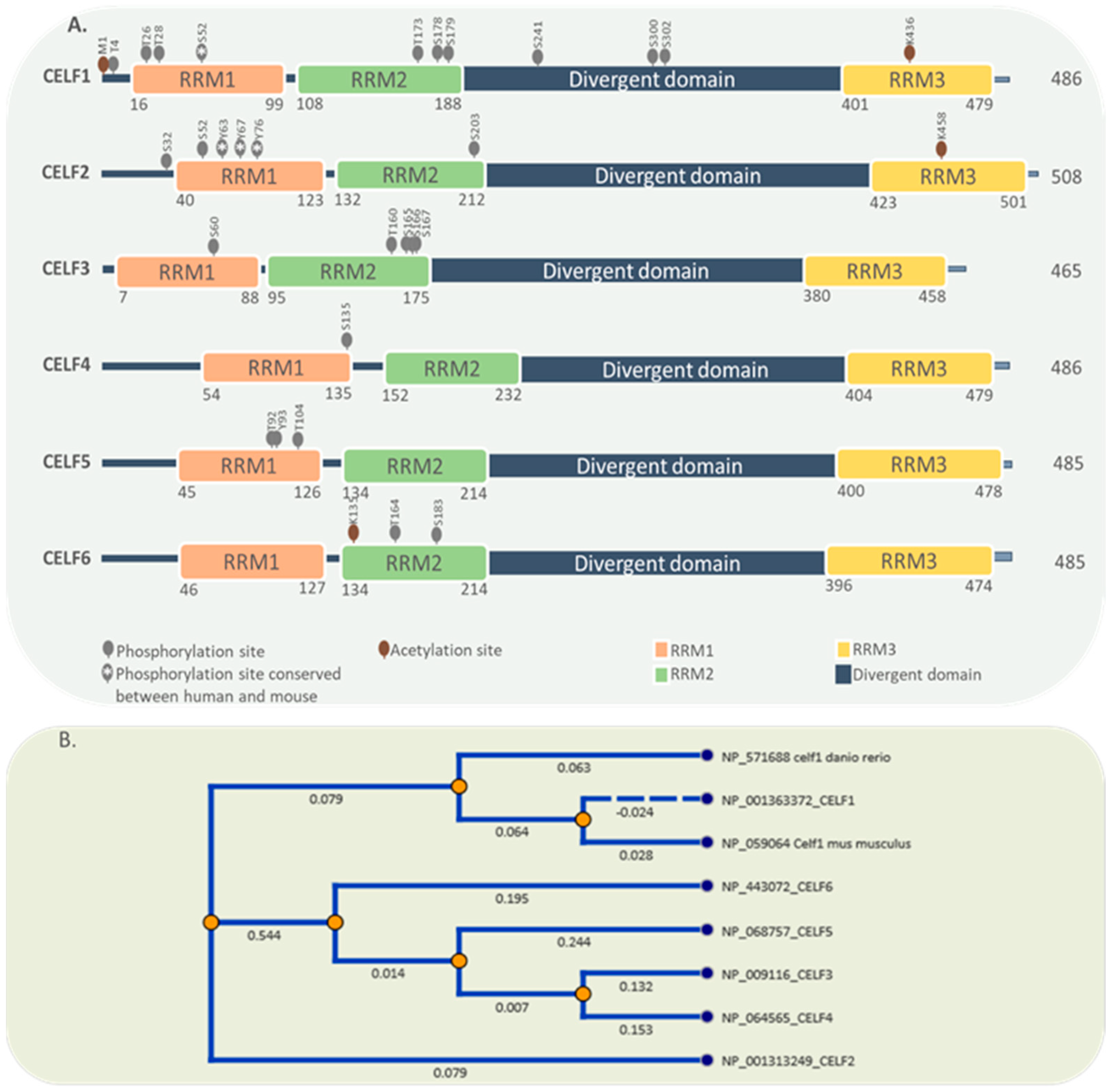
Figure 1. Protein domain structure and post-translational modifications of CELF family members. (A) CELF family members share the same domain structure comprising 3 RRMs and a divergent domain. Phosphorylation and acetylation sites are also shown. (B) Comparisons of the protein sequences suggest three clear branches for the evolution of CELF members. One branch for CELF1, other for CELF2 and a third for the group 3 to 6.
CELF proteins are found in the cytoplasm and nucleus, but they can also be found in stress granules, RNA neuronal granules, and on the PNC (peri-nuclear compartment) under specific circumstances [6][24][25]. As observed most clearly in skeletal muscle loss [26], CELF action remarkably depends on cellular location. CELF2 appears to be primarily or entirely nuclear, unlike CELF1, which is expressed in both the nucleus and the cytoplasm. As a result, despite the fact that they may bind to similar sequences, CELF2 and CELF1 are predicted to have distinct impacts on RNA processing [7][27].
CELF RBPs are primarily involved in alternative splicing and transcript editing in the nucleus. Members of this family are all potential global alternative splicing regulators by generally recognizing CUG and UG-repeat elements [28][29]. According to a genome-wide interaction analysis between CELF1 and its target RNAs, this RBP’s distribution at exon-intron borders, 3UTR, and even exonic regions can comprehensively affect alternative splicing events [8][29]. However, more complex CELF1 binding sites are occasionally required for more specialized splicing processes. An alternative 3′UTR splicing site with two CELF1 binding sites separated by a long intron is required to regulate the mPGES-1 isoforms [30]. Similarly, CELF2 has been identified as an alternative splicing factor for a high number of genes [31]. As observed for exon 6 of LEF1 (Lymphoid Enhancer Binding Factor) and exon 2 of MKK7 (Mitogen-Activated Protein kinase 7) transcripts, CELF2 binding to a downstream or upstream region of an alternate exon can result in that exon being either included or excluded, respectively [7]. CELF2 is additionally a regulator of 3′ UTR intron retention by competing with the U2-auxiliary factor (U2AF65) subunit. Notably, CELF2 mediates the 3′ UTR intron retention of its own transcript, as well as many others [32]. Lastly, CELF2 has been found to be involved in another nuclear function: blocking the C to U editing of apolipoprotein B (APOB) RNA, a process that may be stimulated by CELF2 knockdown [33].
In the cytoplasm, CELF proteins are involved in mRNA stability, translation, alternative polyadenylation (APA), and pre-miRNA maturation [12][32][34][35][36]. CELFs regulate mRNA stability across the genome, although only specific, well-studied events are discussed in this section. CELF1 controls mRNA stability by identifying “UGUUUGUUUGU” element (GU-rich element, GRE) or GU-repeat sequences in the 3′UTR, which mechanistically appear to rely on CELF1 interaction with ribonucleases, such as poly(A)-specific ribonuclease (PARN) [8][26]. PARN 3′-exoribonuclease destabilizes transcripts by removing poly(A) tails at the 3′ end [37]. On the other hand, CELF2 stabilizes the cytoxygenase-2 (COX2) transcript through a different method, which involves binding to A/U-rich regions in the 3′UTR of mRNA [5]. CELF6 has been shown to bind predominantly to certain 3′UTR regions, resulting in a substantial decrease in mRNA levels [12].
CELF proteins have been found to regulate widespread translational activation or inhibition by binding to target transcripts’ 5′UTR or 3′UTR [38]. For instance, the timed translation of Elavl4 (Embryonic Lethal, Abnormal Vision, Drosophila-Like 4) isoforms depends on distinct 5′UTRs controlled by CELF1 during neuronal development [39]. Moreover, CELF1 is able to regulate P21 mRNA stability by binding to its 5′UTR upon proteasome inhibition [40]. Binding through 3′UTR, CELF2 can inhibit the translation of MCL1 (Apoptosis Regulator, BCL2 Family Member) anti-apoptotic factor [41]. Another example is the circularization of SHMT1 (Serine Hydroxymethyltransferase 1) mRNA and its internal ribosome entry site (IRES)- mediated translation, which is provided with CELF1-hnRNPH (Heterogeneous Nuclear Ribonucleoprotein H1) proteins, implying a 5′/3′ interaction in the SHMT1 transcript [42]. Interestingly, following CELF2 depletion, hnRNPC (Heterogeneous Nuclear Ribonucleoprotein C) translation efficiency is shown to be reduced. This is mostly due to the hnRNPC mRNA’s lower translation elongation efficiency [34]. Further pathway study indicates that the knockdown of the CELF1 decreases the expression of some ribosomal proteins in the 40S and 60S subunits, indicating a broader function of the CELF1 in translation [29].
CELF proteins are known to regulate alternative polyadenylation (APA) via two mechanisms. Firstly, CELF1 has been shown to bind to c-FOS (Fos Proto-Oncogene, AP-1 Transcription Factor Subunit) and TNFα (Tumor Necrosis Factor-Alpha) transcripts in a HeLa cell extract and promote deadenylation by interacting with PARN, indicating that CELF1 helps both deadenylation and mRNA stability [35]. Secondly, CELF2 prevents polyadenylation machinery from binding to RNA by competing with CFI (cleavage factor I) and CstF (cleavage stimulatory factor) components in a cell signal-dependent way [32].
Recently, CELF2 was reported as a potential regulator of pre-miR155 maturation. CELF2 is reported to be associated with IL10 (Interleukin-10) to inhibit the maturation of pre-miR155 by blocking KSRP (KH-Type Splicing Regulatory Protein). In this way, it can be considered as negative regulator of miR155 [36]. Similar results are reported for miR140, where CELF1/2 binding interferes with DICER binding to miR140 and thus inhibits its maturation [43].
Finally, CELF RBPs are present in stress granules, neuronal RNA granules, and the peri-nucleolar compartment (PNC). Stress granules (SGs) are dynamic tiny ribonucleoprotein condensates playing functional roles in mRNA processing and degradation and are found in all eukaryotes during cell stress conditions [44]. CELF1 and CELF2 have been found to be required for the trafficking, stability, and translation of p21/CDKN1A and COX-2 to SGs respectively [40][45]. Additionally, CELF4 associates with very large neuronal RNA granules to mediate neurotransmission by regulating the stability, translation, and/or localization of many downstream target mRNAs [25].
PNCs are specialized subnuclear organelles located adjacent to the nucleolus. They are associated with various solid malignant tumors. This region contains transcripts produced by RNA polymerase III, small nuclear RNAs, and many RBPs, including CELF1 [46][47]. In PNCs, CELF1 associates with the RNA component of RNAse MRP (mitochondrial RNA processing) and PTBP (polypyrimidine tract binding protein), interacting with newly synthesized RNA. However, CELF1 enrichment in PNCs may restrict it from performing its cytoplasmic functions [6][48].
In consequence, CELF members are highly divergent, pleiotropic RBPs that are responsible for global alterations in numerous cell types and disease situations [5]. However, CELF proteins are not the only multifunctional RBPs. In two previously published studies, various RBPs influencing different cancers were described [49][50]. The principal functions of CELF family proteins are depicted in Figure 2. Furthermore, they are controlled by numerous biological elements that are discussed specifically in relation to cancer in this article.
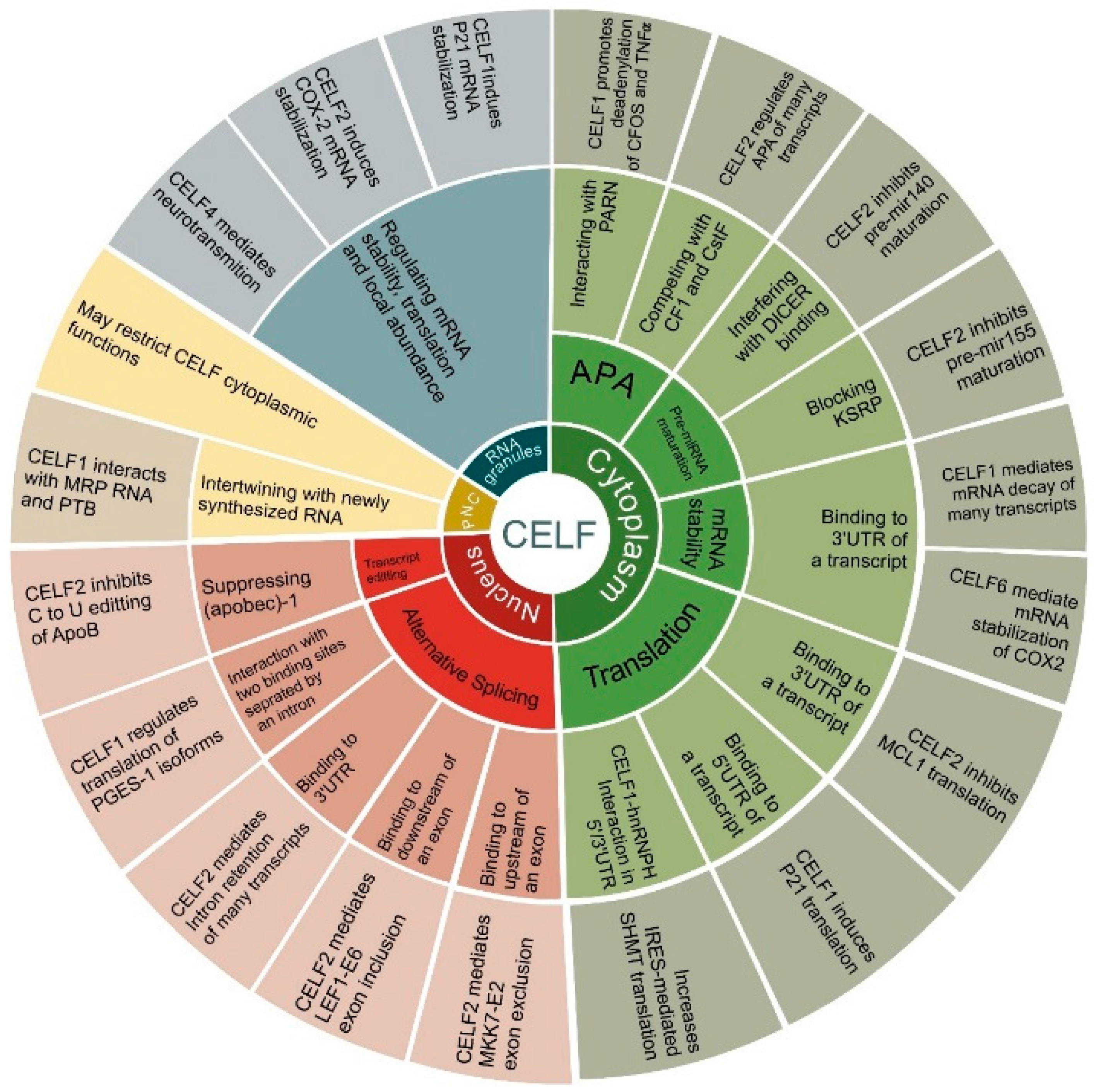
Figure 2. Cellular functions of CELF family members. The circular diagram is divided into 4 main branches illustrating CELF subcellular locations, including the cytoplasm, the nucleus, RNA granules and the peri-nuclear compartment (PNC). Next, each branch is divided into CELFs’ main functions, including translation, mRNA stability, pre-miRNA maturation and alternative polyadenylation (APA) in the cytoplasm, alternative splicing and transcript editing in the nucleus, regulating mRNA stability, translation, local abundance in RNA granules, and, finally, intertwining with newly synthesized RNA in PNCs. In each case, the main mechanism and some representative instances are included for each function. These are discussed in depth in the text.
3. CELF Targets in Cancer
The wide-ranging targets and diverse functions of CELF proteins in cancer have elevated them as prominent players with critical roles in divergent biological pathways. As a result, they might be regarded as prospective prognostic and therapeutic targets. However, present studies on the CELF family, particularly members 3–6, are quite sparse, and it appears that additional studies in this area are required in the future. Notably, despite the structural resemblance between CELF family members, particularly between CELF1 and CELF2 (>90% similarity in their RNA-binding domains), their functions are quite different, notably in cancer [6].
4. CELF Regulation by Noncoding RNAs
Three types of noncoding RNA, including miRNAs, lncRNA, and circRNAs with protein-coding genes, can construct complicated networks regulating the expression of many downstream pathways [9]. Many noncoding RNAs that target CELF members have recently been discovered, and they appear to have a significant influence on the development of cancer. The regulatory networks for CELF1 and CELF2 are illustrated in Figure 3A,B.
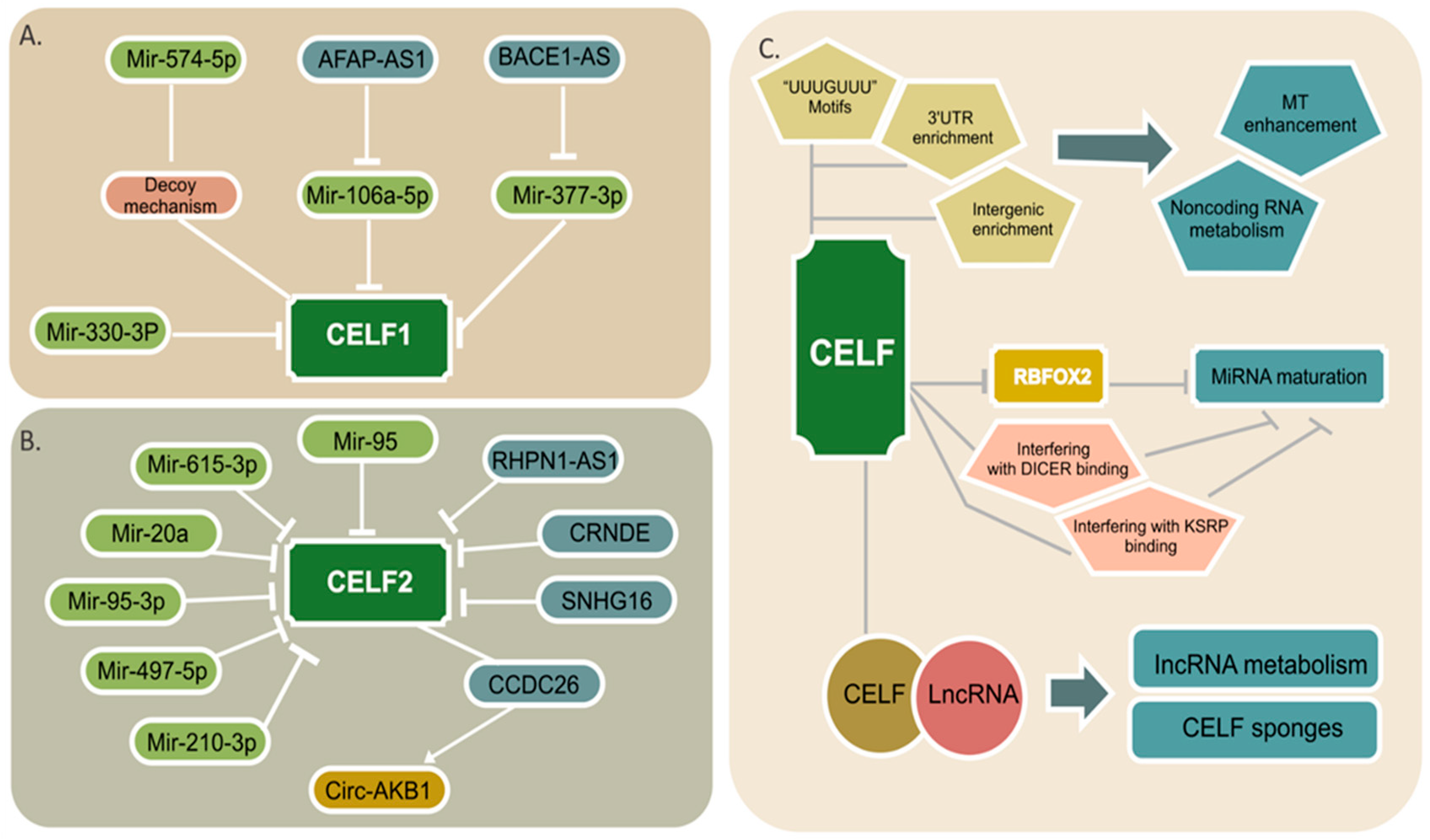
Figure 3. A schematic view of the noncoding RNA/CELF regulatory axis. Multiple miRNAs and lncRNAs targeting each of the CELF1 (A) and CELF2 (B) proteins are shown. The light green color is representative of miRNAs and the blue color is for lncRNAs. The only circular RNA is shown in mustard color. (C) CELF participation in miRNA targeting (MT), noncoding RNA metabolism, and miRNA maturation via both probable and well stablished paths are illustrated. CELF-lncRNA aggregates that are shown in attached circles might play a role in lncRNA metabolism or act as CELF sponges.
MicroRNAs are 21- to 24-nucleotide RNAs, tiny molecules that influence the stability and translation of over 18,000 potential mRNA targets favorably or negatively [51][52]. Mir-330-3p has been shown in certain studies to be an inhibitor of carcinogenesis [53]. This tumor suppressor is reported to have low expression in glioma tissues and cells, and it is able to prevent proliferation and migration by directly binding to CELF1 and suppressing its expression [54]. In human lung tumors, miR-574-5p upregulation promotes tumor development in vivo and increases mPGES-1 expression by preventing CUGBP1 binding to the mPGES-1 3′UTR (decoy mechanism), which leads to an enhanced alternative splicing mPGES-1 and the generation of a novel 3′UTR isoform [55]. The specificity of miR-574-5p/CELF1 regulation on mPGES-1 expression in human lung cancer cells is discovered in a recent proteomic analysis. Patients with lung cancer who have high miR-574-5p levels may benefit significantly from the pharmacological inhibition of PGE2 (Prostaglandin E Synthase 2) formation [30]. It is likely that miRNA-mediated CELF1 inhibition has tumor suppressor effects; however, additional research is needed for different malignancies.
According to numerous studies, CELF2, a tumor suppressor, is also downregulated by a number of miRNAs. In gastric cancer, the inhibition of CELF2 by miR-615-3p enhances proliferation and migration while blocking apoptosis [56]. However, depending on its expression levels, mir-615-3p can have tumor suppressive or carcinogenic effects in some other malignancies. [57]. In glioma cells, CELF2 is a potential target of miR-20a and miR-95-3p, in which the overexpression of these two miRNAs is positively correlated with cell proliferation and invasion [58][59]. In spinal cord glioma tissues, the overexpression of miR-106a-5p exerts its oncogenic function by decreasing CELF2 expression [60]. Exosomes are extracellular vesicles that transport information between healthy cells as well as in tumor microenvironments, allowing cell-to-cell communication. Exosomes from hypoxia colorectal cancer cells transport miR-210-3p to normoxic tumor cells, promoting G1-S cycle transition and proliferation while inhibiting apoptosis by downregulating CELF2 expression [61]. Brucein-D, a drug that targets miR-95, reduces hepatocellular carcinoma (HCC) cell proliferation in vitro and tumor growth in vivo. CELF2 has also been discovered to be a miR-95 downstream target [62].
The miRNA miR-375, which can regulate CELF6, is thought to be associated with pancreatic cancer and breast cancer [63][64][65][66]. In breast cancer, higher levels of miR-375 were expressed in ER-α-positive, where it was a critical factor in cell proliferation and an early event in tumorigenesis [67]. A single nucleotide variation, rs4777498, in CELF6, regulated by miR-375, was previously found to be associated with cervical cancer susceptibility [68]. The upregulation of miR-375 is a key driver of cell proliferation and patients with miR-375 overexpression have a higher probability of local relapse in early breast cancer [65].
LncRNAs are a class of RNA transcripts with more than 200 nucleotides, with secondary and three-dimensional structures that enable them to regulate their targets as cis or trans-acting elements. They are mainly present in the nucleus; however, they can also be found in cytoplasm. By regulating epigenetic, transcriptional, and post-transcriptional modifications, lncRNAs play a key role in a variety of cancers [69]. In hepatocellular carcinoma, the LncRNA BACE1-AS, which plays a crucial role in carcinogenesis, is overexpressed. BACE1-AS inhibits miR-377-3p, and CELF1 is a downstream target of miR-377-3p that serves as an oncogene [70]. In nasopharyngeal carcinoma, LncRNA AFAP1-AS1 can enhance cancer growth by affecting the miR-497-5p/CELF1 axis [71].
The RNA network also has CELF2 as a target. In acute lymphoblastic leukemia (ALL) and acute myeloid leukemia (AML) patients and cell lines, the lncRNA small nucleolar RNA host gene 16 (SNHG16) overexpresses, and by lowering CELF2 mRNA stability, this lncRNA enhances proliferation and migration [72]. In hepatocellular carcinoma, the lncRNA CRNDE (differentially expressed colorectal neoplasia) is considerably overexpressed and is associated with poor clinical outcomes. Mechanically, CRNDE is implied in the inhibition of tumor suppressor genes, including CELF2 and LATS2 (large tumor suppressor 2) [73]. Overexpression of the lncRNA RHPN1-AS1 is associated with enhanced cell viability, proliferation, migration, and invasion in nasopharyngeal carcinoma by targeting CELF2, resulting in an increased mTORC1 signaling pathway [74]. Finally, the LncRNA CCDC26 binds to CELF2, upregulating the expression of circRNA ANKIB1. By sponging miR-195-5p, the overexpression of circRNA ANKIB1 increases PRR11 (Proline Rich 11) protein production, activating the PI3K/AKT and NF-B pathways. The overexpression of the LncRNA CCDC26 in myeloid leukemia enhances cancer development in this way [75].
In summary, CELF proteins play a key role in the noncoding RNA/RBP regulatory axis as they transform noncoding RNA instructions into a variety of downstream signals.
5. Noncoding RNA Regulation by CELF
The flip side of the noncoding RNA/RBP coin is RBPs’ ability to influence noncoding RNA expression and functionality. This is summarized in Figure 3C. The 3′UTR elements play a regulatory role as the primary site of miRNA binding and function, as well as influencing mRNA destiny in terms of translation, stability, and subcellular localization [32]. Meanwhile, the global distribution of CELF members can be found in 3′UTR regions. In addition, intergenic regions that are involved in the process of non-coding RNA biogenesis are also enriched for CELF proteins [7][8][12]. RBPs have recently been found to improve miRNA targeting (MT) by opening mRNA secondary structures [2]. Notably, the “UUUGUUU” motifs, which are similar to CELF binding sites, are enriched adjacent to miRNA binding sites, and their presence tends to potentiate miRNA activity [76].
Cooperative or antagonistic RBP–RBP interactions should not be overlooked in this axis, especially in the case of miRNA regulation. CELF2 appears to play an important function in the pre-miRNA maturation process as it can potentially repress RBFOX2 (RNA Binding Fox-1 Homolog 2) mRNA and protein levels in human cells. These two RBPs have an evolutionary conserved antagonistic relationship that regulates splicing processes in a variety of signaling pathways and transcription factors [31]. The Rbfox proteins’ binding site is assumed to be the conserved GCAUG sequence, which allows them to attach to the terminal loops of miR-20b and miR-107 precursors and inhibit their nuclear processing. MiR-107, in turn, targets Dicer mRNA’s 3′UTR and downregulates it in a way that correlates to the invasiveness of breast cancer cell lines [77]. Rbfox RBPs can influence the expression of downstream targets in cancer by regulating mature miRNA levels. Additionally, RBFOX2 was shown to be one of the specific miRNA hairpin binding proteins binding at multiple targets [43][78].
CELF2 was recently discovered to be a possible regulator of pre-miR155 maturation, with CELF2 knockdown reducing IL10’s ability to block pre-miR155 processing. CELF2 binding to pre-miR155 may hamper DICER’s capacity to bind to and cleave pre-miR155 [36]. Other proteins required for pre-miR155 processing, such as KH-type splicing regulatory protein (KSRP), may also be blocked by CELF2 binding to pre-miR155 [79]. Similar findings have been reported for pre-miR140, in which CELF1/2 binding inhibits pre-miR140 processing by interfering with DICER binding [43]. Another RBP, HuR (also known as ELAV Like RNA Binding Protein 1), is thought to enhance miRNA targeting by attracting AGO (Argonaute) or opening the secondary structure to make AGO more accessible. This RBP can also prevent miRNA targeting by competing against AGO [80]. Competition between CELF1 and HuR for binding to 3′UTR is possible, although this association has only been observed in Myc mRNA translation [81].
The nuclear aggregates of CELF1 and the tumorigenic lincRNAs (long intergenic non-coding RNAs) NEAT1 and NEAT2/MALAT1 may also help to explain how CELF members interact with noncoding RNAs. These aggregates might indicate a function for CELF1 in lincRNA biology, although lincRNAs could also operate as “CELF1 sponges” [82][83]. CELF1/lncRNA combinations with strong affinity have also been seen in colorectal cancer, where CELF1-TNBL aggregates occur in non-perinucleolar areas upon demethylation events [84].
As a result, evaluating all the methods through which CELF members can widely control noncoding RNAs makes it impossible to determine who has the upper hand in this game.
References
- Sternburg, E.L.; Karginov, F.V. Global Approaches in Studying RNA-Binding Protein Interaction Networks. Trends Biochem. Sci. 2020, 45, 593–603.
- Kim, S.; Kim, S.; Chang, H.R.; Kim, D.; Park, J.; Son, N.; Park, J.; Yoon, M.; Chae, G.; Kim, Y.-K.; et al. The regulatory impact of RNA-binding proteins on microRNA targeting. Nat. Commun. 2021, 12, 1–15.
- Sebestyén, E.; Singh, B.; Miñana, B.; Pagès, A.; Mateo, F.; Pujana, M.A.; Valcárcel, J.; Eyras, E. Large-scale analysis of genome and transcriptome alterations in multiple tumors unveils novel cancer-relevant splicing networks. Genome Res. 2016, 26, 732–744.
- Kechavarzi, B.; Janga, S.C. Dissecting the expression landscape of RNA-binding proteins in human cancers. Genome Biol. 2014, 15, R14.
- Dasgupta, T.; Ladd, A.N. The importance of CELF control: Molecular and biological roles of the CUG-BP, Elav-like family of RNA-binding proteins. Wiley Interdiscip. Rev. RNA 2011, 3, 104–121.
- Louis, I.V.-S.; Dickson, A.M.; Bohjanen, P.; Wilusz, C.J. CELFish ways to modulate mRNA decay. Biochim. Biophys. Acta Bioenerg. 2013, 1829, 695–707.
- Ajith, S.; Gazzara, M.R.; Cole, B.S.; Shankarling, G.; Martinez, N.; Mallory, M.J.; Lynch, K.W. Position-dependent activity of CELF2 in the regulation of splicing and implications for signal-responsive regulation in T cells. RNA Biol. 2016, 13, 569–581.
- Xia, H.; Chen, D.; Wu, Q.; Wu, G.; Zhou, Y.; Zhang, Y.; Zhang, L. CELF1 preferentially binds to exon-intron boundary and regulates alternative splicing in HeLa cells. Biochim. Biophys. Acta Bioenerg. 2017, 1860, 911–921.
- Goodall, G.J.; Wickramasinghe, V.O. RNA in cancer. Nat. Rev. Cancer 2020, 21, 22–36.
- Cursons, J.; Pillman, K.A.; Scheer, K.G.; Gregory, P.A.; Foroutan, M.; Hediyeh-Zadeh, S.; Toubia, J.; Crampin, E.J.; Goodall, G.J.; Bracken, C.P.; et al. Combinatorial Targeting by MicroRNAs Co-ordinates Post-transcriptional Control of EMT. Cell Syst. 2018, 7, 77–91.e7.
- Maloney, S.E.; Khangura, E.; Dougherty, J.D. The RNA-binding protein Celf6 is highly expressed in diencephalic nuclei and neuromodulatory cell populations of the mouse brain. Brain Struct. Funct. 2015, 221, 1809–1831.
- Rieger, M.A.; King, D.M.; Crosby, H.; Liu, Y.; Cohen, B.A.; Dougherty, J.D. CLIP and Massively Parallel Functional Analysis of CELF6 Reveal a Role in Destabilizing Synaptic Gene mRNAs through Interaction with 3′ UTR Elements. Cell Rep. 2020, 33, 108531.
- Ladd, A.; Cooper, T.A. Multiple domains control the subcellular localization and activity of ETR-3, a regulator of nuclear and cytoplasmic RNA processing events. J. Cell Sci. 2004, 117, 3519–3529.
- Itai, T.; Hamanaka, K.; Sasaki, K.; Wagner, M.; Kotzaeridou, U.; Brösse, I.; Ries, M.; Kobayashi, Y.; Tohyama, J.; Kato, M.; et al. De novo variants in CELF2 that disrupt the nuclear localization signal cause developmental and epileptic encephalopathy. Hum. Mutat. 2020, 42, 66–76.
- Salisbury, E.; Sakai, K.; Schoser, B.; Huichalaf, C.; Schneider-Gold, C.; Nguyen, H.; Wang, G.-L.; Albrecht, J.H.; Timchenko, L.T. Ectopic expression of cyclin D3 corrects differentiation of DM1 myoblasts through activation of RNA CUG-binding protein, CUGBP. Exp. Cell Res. 2008, 314, 2266–2278.
- Beisang, D.; Rattenbacher, B.; Louis, I.A.V.-S.; Bohjanen, P.R. Regulation of CUG-binding Protein 1 (CUGBP1) Binding to Target Transcripts upon T Cell Activation. J. Biol. Chem. 2012, 287, 950–960.
- Choudhary, C.; Kumar, C.; Gnad, F.; Nielsen, M.L.; Rehman, M.; Walther, T.C.; Olsen, J.V.; Mann, M. Lysine Acetylation Targets Protein Complexes and Co-Regulates Major Cellular Functions. Science 2009, 325, 834–840.
- Gnad, F.; Gunawardena, J.; Mann, M. PHOSIDA 2011: The posttranslational modification database. Nucleic Acids Res. 2010, 39, D253–D260.
- Suzuki, H.; Maegawa, S.; Nishibu, T.; Sugiyama, T.; Yasuda, K.; Inoue, K. Vegetal localization of the maternal mRNA encoding an EDEN-BP/Bruno-like protein in zebrafish. Mech. Dev. 2000, 93, 205–209.
- Choi, D.-K.; Yoo, K.-W.; Hong, S.-K.; Rhee, M.; Sakaki, Y.; Kim, C.-H. Isolation and expression of Napor/CUG-BP2 in embryo development. Biochem. Biophys. Res. Commun. 2003, 305, 448–454.
- Ramalingam, S.; Natarajan, G.; Schafer, C.; Subramaniam, D.; May, R.; Ramachandran, I.; Queimado, L.; Houchen, C.W.; Anant, S. Novel intestinal splice variants of RNA-binding protein CUGBP2: Isoform-specific effects on mitotic catastrophe. Am. J. Physiol. Liver Physiol. 2008, 294, G971–G981.
- Suzuki, H.; Takeuchi, M.; Sugiyama, A.; Alam, A.K.; Vu, L.T.; Sekiyama, Y.; Dam, H.C.; Ohki, S.-Y.; Tsukahara, T. Alternative splicing produces structural and functional changes in CUGBP. BMC Biochem. 2012, 13, 6.
- Hornbeck, P.V.; Zhang, B.; Murray, B.; Kornhauser, J.M.; Latham, V.; Skrzypek, E. PhosphoSitePlus, 2014: Mutations, PTMs and recalibrations. Nucleic Acids Res. 2014, 43, D512–D520.
- Fujimura, K.; Kano, F.; Murata, M. Dual localization of the RNA binding protein CUGBP-1 to stress granule and perinucleolar compartment. Exp. Cell Res. 2008, 314, 543–553.
- Wagnon, J.L.; Briese, M.; Sun, W.; Mahaffey, C.L.; Curk, T.; Rot, G.; Ule, J.; Frankel, W.N. CELF4 Regulates Translation and Local Abundance of a Vast Set of mRNAs, Including Genes Associated with Regulation of Synaptic Function. PLoS Genet. 2012, 8, e1003067.
- Cox, D.C.; Guan, X.; Xia, Z.; Cooper, T.A. Increased nuclear but not cytoplasmic activities of CELF1 protein leads to muscle wasting. Hum. Mol. Genet. 2020, 29, 1729–1744.
- Mallory, M.J.; Jackson, J.; Weber, B.; Chi, A.; Heyd, F.; Lynch, K.W. Signal- and Development-Dependent Alternative Splicing of LEF1 in T Cells Is Controlled by CELF. Mol. Cell. Biol. 2011, 31, 2184–2195.
- Nikonova, E.; Kao, S.-Y.; Ravichandran, K.; Wittner, A.; Spletter, M.L. Conserved functions of RNA-binding proteins in muscle. Int. J. Biochem. Cell Biol. 2019, 110, 29–49.
- Blech-Hermoni, Y.; Dasgupta, T.; Coram, R.J.; Ladd, A.N. Identification of Targets of CUG-BP, Elav-Like Family Member 1 (CELF1) Regulation in Embryonic Heart Muscle. PLoS ONE 2016, 11, e0149061.
- Emmerich, A.C.; Wellstein, J.; Ossipova, E.; Baumann, I.; Lengqvist, J.; Kultima, K.; Jakobsson, P.-J.; Steinhilber, D.; Saul, M.J. Proteomics-Based Characterization of miR-574-5p Decoy to CUGBP1 Suggests Specificity for mPGES-1 Regulation in Human Lung Cancer Cells. Front. Pharmacol. 2020, 11, 196.
- Gazzara, M.R.; Mallory, M.J.; Roytenberg, R.; Lindberg, J.P.; Jha, A.; Lynch, K.W.; Barash, Y. Ancient antagonism between CELF and RBFOX families tunes mRNA splicing outcomes. Genome Res. 2017, 27, 1360–1370.
- Chatrikhi, R.; Mallory, M.J.; Gazzara, M.R.; Agosto, L.M.; Zhu, W.S.; Litterman, A.J.; Ansel, K.M.; Lynch, K.W. RNA Binding Protein CELF2 Regulates Signal-Induced Alternative Polyadenylation by Competing with Enhancers of the Polyadenylation Machinery. Cell Rep. 2019, 28, 2795–2806.e3.
- Chen, Z.; Eggerman, T.L.; Patterson, A.P. ApoB mRNA editing is mediated by a coordinated modulation of multiple apoB mRNA editing enzyme components. Am. J. Physiol. Liver Physiol. 2007, 292, G53–G65.
- Mallory, M.J.; McClory, S.P.; Chatrikhi, R.; Gazzara, M.R.; Ontiveros, R.J.; Lynch, K.W. Reciprocal regulation of hnRNP C and CELF2 through translation and transcription tunes splicing activity in T cells. Nucleic Acids Res. 2020, 48, 5710–5719.
- Moraes, K.C.; Wilusz, C.J.; Wilusz, J. CUG-BP binds to RNA substrates and recruits PARN deadenylase. RNA 2006, 12, 1084–1091.
- Yoon, J.S.J.; Wu, M.K.; Zhu, T.H.; Zhao, H.; Cheung, S.T.; Chamberlain, T.; Mui, A.L.-F. Interleukin-10 control of pre-miR155 maturation involves CELF. PLoS ONE 2020, 15, e0231639.
- Dejene, E.A.; Li, Y.; Showkatian, Z.; Ling, H.; Seto, E. Regulation of poly(a)-specific ribonuclease activity by reversible lysine acetylation. J. Biol. Chem. 2020, 295, 10255–10270.
- Chaudhury, A.; Cheema, S.; Fachini, J.M.; Kongchan, N.; Lu, G.; Simon, L.M.; Wang, T.; Mao, S.; Rosen, D.G.; Ittmann, M.M.; et al. CELF1 is a central node in post-transcriptional regulatory programmes underlying EMT. Nat. Commun. 2016, 7, 13362.
- Popovitchenko, T.; Park, Y.; Page, N.F.; Luo, X.; Krsnik, Z.; Liu, Y.; Salamon, I.; Stephenson, J.D.; Kraushar, M.L.; Volk, N.L.; et al. Translational derepression of Elavl4 isoforms at their alternative 5′ UTRs determines neuronal development. Nat. Commun. 2020, 11, 1674.
- Gareau, C.; Fournier, M.-J.; Filion, C.; Coudert, L.; Martel, D.; Labelle, Y.; Mazroui, R. p21WAF1/CIP1 Upregulation through the Stress Granule-Associated Protein CUGBP1 Confers Resistance to Bortezomib-Mediated Apoptosis. PLoS ONE 2011, 6, e20254.
- Subramaniam, D.; Natarajan, G.; Ramalingam, S.; Ramachandran, I.; May, R.; Queimado, L.; Houchen, C.W.; Anant, S. Translation inhibition during cell cycle arrest and apoptosis: Mcl-1 is a novel target for RNA binding protein CUGBP. Am. J. Physiol. Liver Physiol. 2008, 294, G1025–G1032.
- Fox, J.T.; Stover, P.J. Mechanism of the Internal Ribosome Entry Site-mediated Translation of Serine Hydroxymethyltransferase 1. J. Biol. Chem. 2009, 284, 31085–31096.
- Treiber, T.; Treiber, N.; Plessmann, U.; Harlander, S.; Daiß, J.-L.; Eichner, N.; Lehmann, G.; Schall, K.; Urlaub, H.; Meister, G. A Compendium of RNA-Binding Proteins that Regulate MicroRNA Biogenesis. Mol. Cell 2017, 66, 270–284.e13.
- Amen, T.; Guihur, A.; Zelent, C.; Ursache, R.; Wilting, J.; Kaganovich, D. Resveratrol and related stilbene-derivatives induce Stress Granules with distinct clearance kinetics. Mol. Biol. Cell 2021. mbc.E21–02.
- Moraes, K.C.M.; Monteiro, C.J.; Pacheco-Soares, C. A novel function for CUGBP2 in controlling the pro-inflammatory stimulus in H9c2 cells: Subcellular trafficking of messenger molecules. Cell Biol. Int. 2013, 37, 1129–1138.
- Qian, Y.; Lv, S.; Dong, X. Detection and Significance of PNC in Breast Cancer Tissues. Clin. Lab. 2021, 67.
- Vilimas, T.; Wang, A.Q.; Patnaik, S.; Hughes, E.A.; Singleton, M.D.; Knotts, Z.; Li, D.; Frankowski, K.; Schlomer, J.J.; Guerin, T.M.; et al. Pharmacokinetic evaluation of the PNC disassembler metarrestin in wild-type and Pdx1-Cre;LSL-KrasG12D/+;Tp53R172H/+ (KPC) mice, a genetically engineered model of pancreatic cancer. Cancer Chemother. Pharmacol. 2018, 82, 1067–1080.
- Pollock, C.; Daily, K.; Nguyen, V.T.; Wang, C.; Lewandowska, M.A.; Bensaude, O.; Huang, S. Characterization of MRP RNA–protein interactions within the perinucleolar compartment. Mol. Biol. Cell 2011, 22, 858–866.
- Pereira, B.; Billaud, M.; Almeida, R. RNA-Binding Proteins in Cancer: Old Players and New Actors. Trends Cancer 2017, 3, 506–528.
- Schuschel, K.; Helwig, M.; Hüttelmaier, S.; Heckl, D.; Klusmann, J.-H.; Hoell, J.I. RNA-Binding Proteins in Acute Leukemias. Int. J. Mol. Sci. 2020, 21, 3409.
- Kozomara, A.; Griffiths-Jones, S. miRBase: Annotating high confidence microRNAs using deep sequencing data. Nucleic Acids Res. 2013, 42, D68–D73.
- Matsuyama, H.; Suzuki, H.I. Systems and Synthetic microRNA Biology: From Biogenesis to Disease Pathogenesis. Int. J. Mol. Sci. 2019, 21, 132.
- Qin, G.; Wu, X. Hsa_circ_0032463 acts as the tumor promoter in osteosarcoma by regulating the miR-330-3p/PNN axis. Int. J. Mol. Med. 2021, 47, 1–14.
- Wang, H.; Liu, G.; Li, T.; Wang, N.; Wu, J.; Zhi, H. MiR-330-3p functions as a tumor suppressor that regulates glioma cell proliferation and migration by targeting CELF. Arch. Med Sci. 2020, 16, 1166–1175.
- Saul, M.J.; Baumann, I.; Bruno, A.; Emmerich, A.C.; Wellstein, J.; Ottinger, S.M.; Contursi, A.; Dovizio, M.; Donnini, S.; Tacconelli, S.; et al. miR-574-5p as RNA decoy for CUGBP1 stimulates human lung tumor growth by mPGES-1 induction. FASEB J. 2019, 33, 6933–6947.
- Wang, J.; Liu, L.; Sun, Y.; Xue, Y.; Qu, J.; Pan, S.; Li, H.; Qu, H.; Wang, J.; Zhang, J. miR-615-3p promotes proliferation and migration and inhibits apoptosis through its potential target CELF2 in gastric cancer. Biomed. Pharmacother. 2018, 101, 406–413.
- Wu, H.; Wei, H.; Chen, Q. Long noncoding RNA HOTTIP promotes the metastatic potential of ovarian cancer through the regulation of the miR -615-3p/ SMARCE1 pathway. Kaohsiung J. Med Sci. 2020, 36, 973–982.
- Liao, C.; Chen, W.; Wang, J. MicroRNA-20a Regulates Glioma Cell Proliferation, Invasion, and Apoptosis by Targeting CUGBP Elav-Like Family Member. World Neurosurg. 2019, 121, e519–e527.
- Fan, B.; Jiao, B.-H.; Fan, F.-S.; Lu, S.-K.; Song, J.; Guo, C.-Y.; Yang, J.-K.; Yang, L. Downregulation of miR-95-3p inhibits proliferation, and invasion promoting apoptosis of glioma cells by targeting CELF. Int. J. Oncol. 2015, 47, 1025–1033.
- Xu, H.; Wang, F.; Wang, L. Suppression of miR-106a-5p expression inhibits tumorigenesis via increasing CELF-2 expression in spinal cord glioma. Oncol. Lett. 2021, 22, 1–9.
- Ge, L.; Zhou, F.; Nie, J.; Wang, X.; Zhao, Q. Hypoxic colorectal cancer-secreted exosomes deliver miR-210-3p to normoxic tumor cells to elicit a protumoral effect. Exp. Biol. Med. 2021.
- Xiao, Z.; Chow, S.C.; Li, C.H.; Tang, S.C.; Tsui, S.K.; Lin, Z.; Chen, Y. Role of microRNA-95 in the anticancer activity of Brucein D in hepatocellular carcinoma. Eur. J. Pharmacol. 2014, 728, 141–150.
- Lee, K.H.; Lee, J.K.; Choi, D.W.; Do, I.-G.; Sohn, I.; Jang, K.-T.; Jung, S.-H.; Heo, J.S.; Choi, S.H.; Lee, K.T. Postoperative Prognosis Prediction of Pancreatic Cancer With Seven MicroRNAs. Pancreas 2015, 44, 764–768.
- Zehentmayr, F.; Hauser-Kronberger, C.; Zellinger, B.; Hlubek, F.; Schuster, C.; Bodenhofer, U.; Fastner, G.; Deutschmann, H.; Steininger, P.; Reitsamer, R.; et al. Hsa-miR-375 is a predictor of local control in early stage breast cancer. Clin. Epigenetics 2016, 8, 28.
- Zellinger, B.; Bodenhofer, U.; Engländer, I.A.; Kronberger, C.; Strasser, P.; Grambozov, B.; Fastner, G.; Stana, M.; Reitsamer, R.; Sotlar, K.; et al. Hsa-miR-375/RASD1 Signaling May Predict Local Control in Early Breast Cancer. Genes 2020, 11, 1404.
- Tang, W.; Li, G.-S.; Li, J.-D.; Pan, W.-Y.; Shi, Q.; Xiong, D.-D.; Mo, C.-H.; Zeng, J.-J.; Chen, G.; Feng, Z.-B.; et al. The role of upregulated miR-375 expression in breast cancer: An in vitro and in silico study. Pathol.-Res. Pr. 2019, 216, 152754.
- Jonsdottir, K.; Janssen, S.R.; Da Rosa, F.C.; Gudlaugsson, E.; Skaland, I.; Baak, J.P.A.; Janssen, E.A.M. Validation of Expression Patterns for Nine miRNAs in 204 Lymph-Node Negative Breast Cancers. PLoS ONE 2012, 7, e48692.
- Fang, J.; Li, Y.; Zhang, J.; Yan, M.; Li, J.; Bao, S.; Jin, T. Correlation between polymorphisms in microRNA-regulated genes and cervical cancer susceptibility in a Xinjiang Uygur population. Oncotarget 2017, 8, 31758–31764.
- Chi, Y.; Wang, J.; Yu, W.; Yang, J. Long Non-Coding RNA in the Pathogenesis of Cancers. Cells 2019, 8, 1015.
- Liu, C.; Wang, H.; Tang, L.; Huang, H.; Xu, M.; Lin, Y.; Zhou, L.; Ho, L.; Lu, J.; Ai, X. LncRNA BACE1-AS enhances the invasive and metastatic capacity of hepatocellular carcinoma cells through mediating miR-377-3p/CELF1 axis. Life Sci. 2021, 275, 119288.
- Jin, H.; Liang, G.; Yang, L.; Liu, L.; Wang, B.; Yan, F. SP1-induced AFAP1-AS1 contributes to proliferation and invasion by regulating miR-497-5p/CELF1 pathway in nasopharyngeal carcinoma. Hum. Cell 2021, 34, 491–501.
- Shi, M.; Yang, R.; Lin, J.; Wei, Q.; Chen, L.; Gong, W.; Guo, X. LncRNA-SNHG16 promotes proliferation and migration of acute myeloid leukemia cells via PTEN/PI3K/AKT axis through suppressing CELF2 protein. J. Biosci. 2021, 46, 1–13.
- Xie, S.-C.; Zhang, J.-Q.; Jiang, X.-L.; Hua, Y.-Y.; Xie, S.-W.; Qin, Y.-A.; Yang, Y.-J. LncRNA CRNDE facilitates epigenetic suppression of CELF2 and LATS2 to promote proliferation, migration and chemoresistance in hepatocellular carcinoma. Cell Death Dis. 2020, 11, 1–17.
- Zhao, Y.; Zhou, H.; Dong, W. LncRNA RHPN1-AS1 promotes the progression of nasopharyngeal carcinoma by targeting CELF2 expression. Exp. Mol. Pathol. 2021, 122, 104671.
- Li, C.; Mu, J.; Shi, Y.; Xin, H. LncRNA CCDC26 Interacts with CELF2 Protein to Enhance Myeloid Leukemia Cell Proliferation and Invasion via the circRNA_ANKIB1/miR-195-5p/PRR11 Axis. Cell Transplant. 2021, 30.
- Jacobsen, A.; Wen, J.; Marks, D.S.; Krogh, A. Signatures of RNA binding proteins globally coupled to effective microRNA target sites. Genome Res. 2010, 20, 1010–1019.
- Chen, Y.; Zubovic, L.; Yang, F.; Godin, K.; Pavelitz, T.; Castellanos, J.; Macchi, P.; Varani, G. Rbfox proteins regulate microRNA biogenesis by sequence-specific binding to their precursors and target downstream Dicer. Nucleic Acids Res. 2016, 44, 4381–4395.
- Nussbacher, J.K.; Yeo, G.W. Systematic Discovery of RNA Binding Proteins that Regulate MicroRNA Levels. Mol. Cell 2018, 69, 1005–1016.e7.
- Ruggiero, T.; Trabucchi, M.; De Santa, F.; Zupo, S.; Harfe, B.D.; McManus, M.T.; Rosenfeld, M.G.; Briata, P.; Gherzi, R. LPS induces KH-type splicing regulatory protein-dependent processing of microRNA-155 precursors in macrophages. FASEB J. 2009, 23, 2898–2908.
- Ahuja, D.; Goyal, A.; Ray, P.S. Interplay between RNA-binding protein HuR and microRNA-125b regulates p53 mRNA translation in response to genotoxic stress. RNA Biol. 2016, 13, 1152–1165.
- Liu, L.; Ouyang, M.; Rao, J.N.; Zou, T.; Xiao, L.; Chung, H.K.; Wu, J.; Donahue, J.M.; Gorospe, M.; Wang, J.-Y. Competition between RNA-binding proteins CELF1 and HuR modulates MYC translation and intestinal epithelium renewal. Mol. Biol. Cell 2015, 26, 1797–1810.
- Le Tonquèze, O.; Gschloessl, B.; Legagneux, V.; Paillard, L.; Audic, Y. Identification of CELF1 RNA targets by CLIP-seq in human HeLa cells. Genom. Data 2016, 8, 97–103.
- Ebert, M.S.; Sharp, P.A. Emerging Roles for Natural MicroRNA Sponges. Curr. Biol. 2010, 20, R858–R861.
- Dumbovic, G.; Biayna, J.; Banús, J.; Samuelsson, J.; Roth, A.; Diederichs, S.; Alonso, S.; Buschbeck, M.; Perucho, M.; Forcales, S.-V. A novel long non-coding RNA from NBL2 pericentromeric macrosatellite forms a perinucleolar aggregate structure in colon cancer. Nucleic Acids Res. 2018, 46, 5504–5524.
More
Information
Contributor
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
897
Revisions:
2 times
(View History)
Update Date:
25 Oct 2021
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No