Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Dong, M. Narrow- versus Wide-Ranged Ostrya Species. Encyclopedia. Available online: https://encyclopedia.pub/entry/15295 (accessed on 23 June 2026).
Dong M. Narrow- versus Wide-Ranged Ostrya Species. Encyclopedia. Available at: https://encyclopedia.pub/entry/15295. Accessed June 23, 2026.
Dong, Ming. "Narrow- versus Wide-Ranged Ostrya Species" Encyclopedia, https://encyclopedia.pub/entry/15295 (accessed June 23, 2026).
Dong, M. (2021, October 22). Narrow- versus Wide-Ranged Ostrya Species. In Encyclopedia. https://encyclopedia.pub/entry/15295
Dong, Ming. "Narrow- versus Wide-Ranged Ostrya Species." Encyclopedia. Web. 22 October, 2021.
Copy Citation
Four sympatric Ostrya species have different geographic range sizes. O. multinervis and O. rehderiana are narrow-ranged species with narrow potential geographical distributions. O. japonica and O. trichocarpa, both of which have wide potential geographical distributions, are wide-ranged species.
climate change
conservation management
geographic range size
1. Introduction
Geographic range size, the area in which a species occurred, is determined by intrinsic (e.g., propagule or body size, dispersal ability, population density) and extrinsic (e.g., local environmental conditions, topographical features) factors [1][2]. Among those complex closely related factors, the dominant factors are usually changing and may vary among organisms and species [3][4][5][6][7]. Numerous hypotheses have been proposed to explain the diversification in geographical range size among species and across space, such as geometric constraint [8], habitat area [9], climatic variability [3], dispersal ability [6], niche breadth [10].
Among the mentioned above, niche breadth–range size hypothesis states that geographic range size is positively associated with environmental niche breadth and has been supported in some species (e.g., Gaston and Spicer [11], Boulangeat et al. [12]; Botts et al. [13], Carrillo-Angeles et al. [14]; Yu et al. [15]; Vincent et al. [16]). Species with broad niche may have better adaptation to various environmental types than those with narrow niche [17][18][19][20]. If this hypothesis holds true, it can be inferred that species having wide distribution range currently will further widen their distribution ranges, while species having narrow distribution range currently will further narrow their distribution ranges, under changed climate in the future [21]. However, the positive relationship is still uncertain in different taxonomic groups; additional evidence is required to support this hypothesis [22][23][24].
Moreover, geographic range size is usually used to predict the extinction vulnerability or invasion risk of a given species [25][26]. Predicting the potential geographic distribution range size and identifying the dominant factors would underlie biodiversity conservation and management [27][28][29]. Species distribution models (SDMs) are practical to predict the potential geographic distribution under different climate scenarios [30][31], which have been widely used for the conservation of rare species [14][32][33][34]. Maximum entropy (Maxent) modeling is one of the most commonly used predictive methods in SDMs [35], which can achieve greater predictive accuracy than other methods with presence-only and biased sampling data [33][36][37][38]. Benefiting from the statistical mechanics, the Maxent approach is powerful modeling for geographic distributions of rare species with narrow ranges and presence-only data [39][40][41][42]. The Maxent approach also performed well for distribution prediction of widespread species [33][43][44].
2. Study Species
Genus Ostrya Scopoli (Betulaceae) contains 8 tree species, commonly named hop-hornbeam or ironwood due to their hard and heavy wood texture [45][46][47][48]. Ostrya contains some species having been regarded as threatened species, similar to Carpinus of the same family [34][42][48][49][50]. Ostrya species are distributed in the subtropical and temperate forests of China with five species recorded in Flora of China (http://www.efloras.org (accessed on 15 June 2020)) [49], including O. japonica, O. multinervis, O. rehderiana, O. trichocarpa, and O. yunnanensis. Based on the latest phylogenetic analysis [51], O. yunnanensis was treated as O. trichocarpa in data processing. Thus, four sympatric species of Ostrya were included in this study, i.e., O. japonica, O. multinervis, O. rehderiana, and O. trichocarpa [51].
All of the Ostrya species are deciduous trees with scaly and rough barks. The male inflorescences would be formed from April to July, blooming in the spring of next year [49]. O. japonica is widespread from southwest to northeast China, while the other three Ostrya species have few occurrence records and narrow distributions in China. The Ostrya species differ in distribution regions, including O. japonica distributed in temperate deciduous forests (985–2800 m a.s.l.), O. multinervis in subtropical mixed forests (600–1300 m a.s.l.), O. rehderiana in subtropical evergreen broad-leaved forests (200–400 m a.s.l.), and O. trichocarpa in subtropical moist broadleaf forests (956–2600 m a.s.l.) [49][51]. O. rehderiana and O. multinervis had similar wild geographic distribution in the last glacial maximum (LGM), but then the habitat of the former was reduced dramatically while the latter maintained a stable population size [52]. Furthermore, O. rehderiana is on the International Union for Conservation of Nature (IUCN)’s Red List of Threatened Species as critically endangered [50]. It is also among the first-class state protection wild plants in China based on the latest report [53]. Ostrya species have different geographic range sizes, providing an opportunity to test the niche breadth–range size hypothesis through expectations of the models using narrowly distributed rare versus widespread congeneric species.
3. Predict the distribution of Ostrya species
We collected occurrence records of four Ostrya species in China. We used Maxent to predict the potential distributions under present and future climate scenarios, and calculated the niche breadth of each species. Potential distribution areas of wide niche breadth species (O. japonica and O. trichocarpa) were significantly wider than those of narrow niche breadth species (O. multinervis and O. rehderiana)(Figure 1). In the future scenarios of global climate change, wide-ranged O. japonica would have wider potential distribution than in the current scenario, even expand their geographic range (Figure 2). Conversely, suitable habitats of narrow-ranged O. multinervis and O. rehderiana would be reduced strikingly in future scenarios as compared with in current scenario, might subjected to a high risk of extinction. Potential distribution range sizes of the Ostrya species would positively correlate with their niche breadths in future scenarios and their niche breadths would determine their distribution variation with climate change.
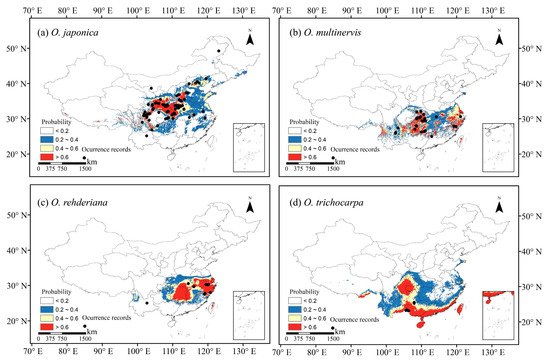
Figure 1. Occurrence records used in models and potential habitats in China of four species of Ostrya, i.e., O. japonica (a), O. multinervis (b), O. rehderiana (c), and O. trichocarpa (d), under present climate scenario. Note: Black dots represent the occurrence record locations of species in China. The values for species occurrence probability ranged between 0 and 1, i.e., least potential (<0.2), moderate potential (0.2–0.4), good potential (0.4–0.6), and high potential (>0.6).
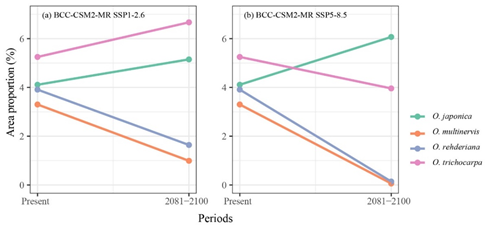
Figure 2. Great probability habitat proportion shifts of four Ostrya species under climate scenarios, i.e., BCC-CSM2-MR SSP1-2.6 (a), and BCC-CSM2-MR SSP5-8.5 (b), between the present and 2081–2100 periods.
4. Contributions of Climate Variables
The minimum temperature of the coldest month (BIO6) was the principal temperature factor impacting the distributions of four Ostrya species, achieving 59.87% and 70.12% in the contributions of O. rehderiana and O. trichocarpa, respectively. The lowest temperature in winter around ±5 °C was suitable for the growth of O. multinervis and O. rehderiana, while the other two species could adopt a wider range of BIO6 (Figure 3).
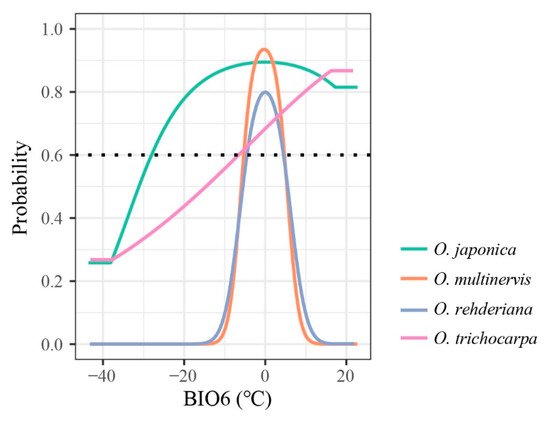
Figure 3. Response curves of four Ostrya species, i.e., O. japonica, O. multinervis, O. rehderiana, and O. trichocarpa, presence probability affected by BIO6 (minimum temperature of coldest month, °C). The black dotted line represents that the probability of species existing is 0.6; species would archive high potential adaptation above this line.
References
- Schwartz, M.W.; Iverson, L.R.; Prasad, A.M.; Matthews, S.N.; O’Connor, R.J. Predicting extinctions as a result of climate change. Ecology 2006, 87, 1611–1615.
- Hayes, K.R.; Barry, S.C. Are there any consistent predictors of invasion success? Biol. Invasions 2008, 10, 483–506.
- Stevens, G.C. The latitudinal gradient in geographical range: How so many species coexist in the tropics. Am. Nat. 1989, 133, 240–256.
- Brown, J.H.; Stevens, G.C.; Kaufman, D.M. The geographic range: Size, shape, boundaries, and internal structure. Annu. Rev. Ecol. Evol. Syst. 1996, 27, 597–623.
- Gaston, K.J. Species-range-size distributions: Patterns, mechanisms and implications. Trends Ecol. Evol. 1996, 11, 197–201.
- Lester, S.E.; Ruttenberg, B.I.; Gaines, S.D.; Kinlan, B.P. The relationship between dispersal ability and geographic range size. Ecol. Lett. 2007, 10, 745–758.
- Sheth, S.N.; Angert, A.L. The evolution of environmental tolerance and range size: A comparison of geographically restricted and widespread. Mimulus. Evol. 2014, 68, 2917–2931.
- Colwell, R.K.; Hurtt, G.C. Nonbiological gradients in species richness and a spurious Rapoport effect. Am. Nat. 1994, 144, 570–595.
- Ohlemüller, R.; Anderson, B.J.; Araujo, M.B.; Butchart, S.H.; Kudrna, O.; Ridgely, R.S.; Thomas, C.D. The coincidence of climatic and species rarity: High risk to small-range species from climate change. Biol. Lett. 2008, 4, 568–572.
- Brown, J.H. On the relationship between abundance and distribution of species. Am. Nat. 1984, 124, 255–279.
- Gaston, K.J.; Spicer, J.I. The relationship between range size and niche breadth: A test using five species of Gammarus (Amphipoda). Glob. Ecol. Biogeogr. 2001, 10, 179–188.
- Boulangeat, I.; Lavergne, S.; van Es, J.; Garraud, L.; Thuiller, W. Niche breadth, rarity and ecological characteristics within a regional flora spanning large environmental gradients. J. Biogeogr. 2012, 39, 204–214.
- Botts, E.A.; Erasmus, B.F.N.; Alexander, G.J.; Lawlor, J. Small range size and narrow niche breadth predict range contractions in South African frogs. Glob. Ecol. Biogeogr. 2013, 22, 567–576.
- Carrillo-Angeles, I.G.; Suzán-Azpiri, H.; Mandujano, M.C.; Golubov, J.; Martínez-Ávalos, J.G. Niche breadth and the implications of climate change in the conservation of the genus Astrophytum (Cactaceae). J. Arid Environ. 2016, 124, 310–317.
- Yu, F.; Groen, T.A.; Wang, T.; Skidmore, A.K.; Huang, J.; Ma, K. Climatic niche breadth can explain variation in geographical range size of alpine and subalpine plants. Int. J. Geogr. Inf. Sci. 2016, 31, 190–212.
- Vincent, H.; Bornand, C.N.; Kempel, A.; Fischer, M. Rare species perform worse than widespread species under changed climate. Biol. Invasions 2020, 246.
- Lynch, M.; Gabriel, W. Environmental tolerance. Am. Nat. 1987, 129, 283–303.
- Bolnick, D.I.; Svanbäck, R.; Fordyce, J.A.; Yang, L.H.; Davis, J.M.; Hulsey, C.D.; Forister, M.L. The ecology of individuals: Incidence and implications of individual specialization. Am. Nat. 2003, 161, 1–28.
- Schwilk, D.W.; Ackerly, D.D. Limiting similarity and functional diversity along environmental gradients. Ecol. Lett. 2005, 8, 272–281.
- Siqueira, T.; Bini, L.M.; Roque, F.O.; Marques Couceiro, S.R.; Trivinho-Strixino, S.; Cottenie, K. Common and rare species respond to similar niche processes in macroinvertebrate metacommunities. Ecography 2012, 35, 183–192.
- Carscadden, K.A.; Emery, N.C.; Arnillas, C.A.; Cadotte, M.W.; Afkhami, M.E.; Gravel, D.; Livingstone, S.W.; Wiens, J.J. Niche breadth: Causes and consequences for ecology, evolution, and conservation. Quart. Rev. Biol. 2020, 95, 179–214.
- Slatyer, R.A.; Hirst, M.; Sexton, J.P. Niche breadth predicts geographical range size: A general ecological pattern. Ecol. Lett. 2013, 16, 1104–1114.
- Hirst, M.J.; Griffin, P.C.; Sexton, J.P.; Hoffmann, A.A. Testing the niche-breadth–range-size hypothesis: Habitat specialization vs. performance in Australian alpine daisies. Ecology 2017, 98, 2708–2724.
- Cai, Q.; Welk, E.; Ji, C.; Fang, W.; Sabatini, F.M.; Zhu, J.; Zhu, J.; Tang, Z.; Attorre, F.; Campos, J.A.; et al. The relationship between niche breadth and range size of beech (Fagus) species worldwide. J. Biogeogr. 2021, 48, 1240–1253.
- Gaston, K.J.; Fuller, R.A. The sizes of species’ geographic ranges. J. Appl. Ecol. 2009, 46, 1–9.
- Xu, W.B.; Svenning, J.C.; Chen, G.K.; Zhang, M.G.; Huang, J.H.; Chen, B.; Ordonez, A.; Ma, K.P. Human activities have opposing effects on distributions of narrow-ranged and widespread plant species in China. Proc. Natl. Acad. Sci. USA 2019, 116, 26674–26681.
- Gaston, K.J. Species-range size distributions: Products of speciation, extinction and transformation. Philos. Trans. R. Soc. B 1998, 353, 219–230.
- Purvis, A.; Gittleman, J.L.; Cowlishaw, G.; Mace, G.M. Predicting extinction risk in declining species. Proc. R. Soc. B. Biol. Sci. 2000, 267, 1947–1952.
- Wan, J.Z.; Wang, C.J.; Yu, F.H. Spatial conservation prioritization for dominant tree species of Chinese forest communities under climate change. Clim. Chang. 2017, 144, 303–316.
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697.
- Zurell, D.; Franklin, J.; König, C.; Bouchet, P.J.; Dormann, C.F.; Elith, J.; Fandos, G.; Feng, X.; Guillera-Arroita, G.; Guisan, A.; et al. A standard protocol for reporting species distribution models. Ecography 2020, 43, 1261–1277.
- Franklin, J. Species distribution models in conservation biogeography: Developments and challenges. Divers. Distrib. 2013, 19, 1217–1223.
- Merow, C.; Smith, M.J.; Silander, J.A. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069.
- He, Q.; Zhao, R.; Zhu, Z. Geographical distribution simulation and comparative analysis of Carpinus viminea and C. londoniana. Glob. Ecol. Conserv. 2020, 21, e00825.
- Elith, J.; Graham, C.H. Do they? How do they? Why do they differ? On finding reasons for differing performances of species distribution models. Ecography 2009, 32, 66–77.
- Pearson, R.G.; Raxworthy, C.J.; Nakamura, M.; Peterson, A.T. Predicting species distributions from small numbers of occurrence records: A test case using cryptic geckos in Madagascar. J. Biogeogr. 2006, 34, 102–117.
- Tsoar, A.; Allouche, O.; Steinitz, O.; Rotem, D.; Kadmon, R. A comparative evaluation of presence-only methods for modelling species distribution. Divers. Distrib. 2007, 13, 397–405.
- Kaky, E.; Nolan, V.; Alatawi, A.; Gilbert, F. A comparison between Ensemble and MaxEnt species distribution modelling approaches for conservation: A case study with Egyptian medicinal plants. Ecol. Inform. 2020, 60, 101150.
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudik, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57.
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893.
- Rhoden, C.M.; Peterman, W.E.; Taylor, C.A. Maxent-directed field surveys identify new populations of narrowly endemic habitat specialists. PeerJ 2017, 5, e3632.
- Zhao, R.; Chu, X.; He, Q.; Tang, Y.; Song, M.; Zhu, Z. Modeling current and future potential geographical distribution of Carpinus tientaiensis, a critically endangered species from China. Forests 2020, 11, 774.
- Visger, C.J.; Germain-Aubrey, C.C.; Patel, M.; Sessa, E.B.; Soltis, P.S.; Soltis, D.E. Niche divergence between diploid and autotetraploid Tolmiea. Am. J. Bot. 2016, 103, 1396–1406.
- Yu, F.; Wang, T.; Groen, T.A.; Skidmore, A.K.; Yang, X.; Ma, K.; Wu, Z. Climate and land use changes will degrade the distribution of Rhododendrons in China. Sci. Total. Environ. 2019, 659, 515–528.
- Bozkurt, A.; Erdin, N. Wood Material Technology Handbook; Istanbul University Publication, Faculty of Forestry Publication: Istanbul, Turkey, 1997.
- Korkut, S.; Guller, B. Physical and mechanical properties of European Hophornbeam (Ostrya carpinifolia Scop.) wood. Bioresour. Technol. 2008, 99, 4780–4785.
- Chen, Z.D.; Manchester, S.R.; Sun, H.Y. Phylogeny and evolution of the Betulaceae as inferred from DNA sequences, morphology, and paleobotany. Am. J. Bot. 1999, 86, 1168–1181.
- Holstein, N.; Weigend, M. No taxon left behind?—A critical taxonomic checklist of Carpinus and Ostrya (Coryloideae, Betulaceae). Eur. J. Taxon. 2017, 375, 1–52.
- Fang, Z.; Zhao, S.; Skvortsov, A. Flora of China. Harv. Pap. Bot. 1999, 4, 300–301.
- Shaw, K.; Roy, S.; Wilson, B. The IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2014.
- Lu, Z.; Zhang, D.; Liu, S.; Yang, X.; Liu, X.; Liu, J. Species delimitation of Chinese hop-hornbeams based on molecular and morphological evidence. Ecol. Evol. 2016, 6, 4731–4740.
- Yang, Y.; Ma, T.; Wang, Z.; Lu, Z.; Li, Y.; Fu, C.; Chen, X.; Zhao, M.; Olson, M.S.; Liu, J. Genomic effects of population collapse in a critically endangered ironwood tree Ostrya rehderiana. Nat. Commun. 2018, 9, 5449.
- National Forestry and Grassland Administration, National Key Protected Wild Plant List. Available online: https://www.forestry.gov.cn/main/153/20200710/085720879652689.html (accessed on 9 July 2020).
More
Information
Subjects:
Ecology; Area Studies
Contributor
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.1K
Entry Collection:
Environmental Sciences
Revisions:
3 times
(View History)
Update Date:
25 Oct 2021
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No