 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Federica Rizzi | + 3350 word(s) | 3350 | 2021-08-06 11:25:24 | | | |
| 2 | Conner Chen | Meta information modification | 3350 | 2021-08-26 05:48:02 | | |
Video Upload Options
Tumorigenesis is characterized by an uncontrolled proliferation of cells that have usually undergone genetic mutations due to hereditary, environmental, and lifestyle factors. Cancer progression, persistent inflammation of the surrounding tissue, and extracellular matrix (ECM) remodeling are three highly interconnected processes. Indeed, a multifaceted network of inflammatory signals produced by cancer cells and innate immune cells recruited in the tumor microenvironment (TME) induce changes in the surrounding stroma that, in turn, influence the homeostasis of ECM, creating a “cancerized” microenvironment that supports tumor growth and metastasis. The ECM represents a well-organized and heterogeneous network of macromolecules that provides a mechanical scaffold to the cells and mediates the diffusion of signaling molecules to sustain cell functions.
1. Inflammation and ECM Remodeling, Close Allies in Cancer Progression
In 1986, Dvorak defined cancers as “wounds that do not heal”. This definition comes from the observation that tumor growth leads to a continuous injury of the surrounding stromal tissue that triggers, in turn, a chronic response in the attempt to repair the lesion and restore tissue homeostasis [1]. This response is very complex and multifaceted and includes the concerted activity of inflammatory molecules, immune cells, and fibroblasts, which together lead to a state of persistent inflammation.
When a tumor lesion occurs, immune cells, as leukocytes, macrophages, and/or bone marrow-derived myeloid precursors, are recruited at the injured site. These tumor-infiltrating cells, along with cancer cells, release in the TME high levels of transforming growth factor β (TGFβ), a cytokine that exerts pleiotropic effects on both cancer and normal cells adjacent to rupture through autocrine and paracrine signaling mechanisms [2].
In normal and premalignant stages of cell transformation, TGFβ exerts tumor-inhibiting functions by suppressing cancer cell growth through anti-proliferative and pro-apoptotic signaling [3]. Moreover, TGFβ limits the proliferation and differentiation of cells of the innate and adaptive immune system, thus suppressing tumorigenic inflammation. However, at more advanced stages of carcinogenesis, tumor cells acquire inactivating mutations of TGFβ receptors and/or alteration of their signaling transduction pathway (reviewed in [3]) and become resistant to the negative regulatory effects of this cytokine. As a consequence, TGFβ switches from a tumor suppressor factor to a metastasis promoter and drives changes in the TME that finally sustain tumor growth [4]. Indeed, high levels of TGFβ in the TME enable cancer cells to escape the immune surveillance leading to an over-production of cytokines and chemokines that contribute to boosting chronic inflammation [3][4]. Moreover, TGFβ produced by cancer cells acts in a paracrine way on stromal cells stimulating the secretion of growth factors and mitogens, as the platelet-derived growth factor (PDGF), and driving the trans-differentiation of stromal progenitors, including resident fibroblasts, endothelial cells, preadipocytes, and bone marrow-derived mesenchymal stem cells (MSCs) into “activated fibroblast” [5][6][7]. These “activated fibroblasts”, collectively named CAFs, bear a myofibroblast-like phenotype, express higher levels of αSMA, collagen 11-α1 (COL11A1), PDGF receptor (PDGFR) α/β, and lower levels of caveolin-1 than normal fibroblasts, and acquire greater contractile and proliferating capacities [8].
However, to date, specific markers to outline the molecular profile of CAFs are still lacking, making it difficult to better understand the biology of these heterogeneous classes of cells [9]. Once activated, CAFs themselves release TGFβ and feed an autocrine-positive signaling loop that sustains tumor progression by contributing to the generation of a permissive TME [10], as illustrated in Figure 1.
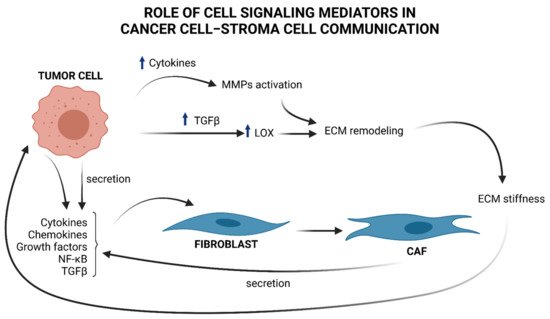
Figure 1. Brief overview of cell signaling mediators in cancer cell–stroma cell communication and ECM remodeling. Tumor cells produce cytokines, chemokines, NF-κB, and TGFβ, that “activate” stromal progenitors, such as fibroblasts, into CAFs and produce, in turn, cytokines and TGFβ, influencing tumor cell functions. TGFβ upregulation in cancer cells provokes an increment in LOX, which in turn remodels ECM by favoring collagen fibers cross-linking. As a consequence ECM stiffness increases and induces the mechanical activation of latent TGFβ. Cytokines produced by tumor cells contributes to MMPs activation. These enzymes take part to ECM remodeling and sustains tumor progress.
Under TGFβ stimulation, CAFs release high levels of cytokines and chemokines, contributing to attracting neutrophils, macrophages, lymphocytes, and natural killer (NK) cells at the site of the tumor lesion, thus boosting the inflammatory response that favors cancer progression [7]. One of the best characterized pro-tumorigenic cytokines is interleukin 6 (IL-6), known to be linked to increased risk of development of a large variety of cancers [11]. IL-6, secreted by CAFs, binds to its cognate receptor expressed by tumor cells and promotes their growth and the acquisition of an invasive phenotype by activating the IL-6–JAK–STAT axis and the notch pathway [12]. IL-6 secreted from cancer cells also acts in a paracrine manner to induce the differentiation of fibroblasts into CAFs. By these efferent (from tumor to stroma) and afferent (from stroma to tumor) signaling pathways, IL-6 is one of the molecules secreted in the TME that participates in the tumor–stroma cross-talk required for tumor growth [13].
Tumor necrosis factor-α (TNFα) is another cytokine secreted at high levels in the TME during the early stages of tumorigenesis that has an established role in chronic inflammation, angiogenesis, tissue remodeling, tumor growth, and metastasis [14]. Similarly to TGFβ, the role played by TNFα in cancer progression is both tumor-inhibiting and tumor-promoting, depending on the cell context and the cancer stage. Indeed, although high levels of TNFα inhibit tumor growth [15], secretion of low levels of TNFα by cancer cells in a B16 mouse melanoma model furthers the recruitment of infiltrating myeloid cells, promoting cancer vascularization and progression [16]. Furthermore, in a mouse xenograft model of ovarian carcinoma, TNFα receptor 1 (TNFR1) expression on CD4+ T cells is necessary for IL-17 secretion and myeloid cell recruitment in tumors, sustaining inflammation and cancer progression [17].
Apart from participating in the complex signaling between tumor and surrounding stroma, CAFs are also responsible for the deposition of ECM components (see Figure 1), a physiological process that becomes highly unregulated in the early stages of cancer [18]. This dysregulation compromises abundance, concentration, structure, and architecture of the ECM, affecting tumor establishment and progression. For example, deposition of collagen by CAFs, together with changes in the expression of the enzymes responsible for its remodeling, leads to an impairment of the organization of collagen fibers and results in increased stiffness of the ECM [19]. In breast cancer, the collagen fibers become linearized and perpendicularly oriented at the tumor boundary, thus favoring the migration and invasion of cancer cells [20]. The lysyl oxidases (LOX) enzymes are responsible for the cross-linking of collagen and elastin fibers. The expression of LOX is upregulated in many cancers by a mechanism that is, at least in part, TGFβ-driven [21]. An unbalanced enhancement in LOX expression and activity increases ECM stiffness that induces the mechanical activation of latent TGFβ and fuels the onset of a vicious circle that maintains a tumor-promoting inflammatory environment [19]. The activity of LOX enzymes is therefore not restricted to the shaping of the architecture and the mechanical functions of the ECM but is part of a complex network of bi-directional signaling within the matrix and different cells of the microenvironment that contributes to establish and maintain an inflamed, immune-suppressive, and pro-carcinogenic TME [21].
The activity of ECM-degrading proteases, for example, releases matrix-bound growth factors and cytokines, which can mediate the activation of downstream effectors, such as various oncogenic transcription factors [2][18].
Among these, NF-κB is a prototypical inflammatory mediator involved in tumorigenesis. NF-κB transient activation is tightly controlled under physiological conditions that promote inflammation, as an adaptive, physiological response. However, in cancer proinflammatory cytokines, oncogenic growth factors and tyrosine kinases promote the constitutive activation of NF-κB through autocrine and paracrine ways [22]. This persistent activation yields to the transcriptional regulation of various ECM components and ECM degrading enzymes, such as MMPs, whose expression is under the control of NF-κB [23].
To degrade the ECM, different types of cells which constitute the tumor mass, i.e., the actual cancer cells, activated fibroblasts, and macrophages, produce a large number of enzymes, belonging to the metzincin protease superfamily of zinc-endopeptidases, which include MMPs, also known as matrixins, a disintegrin and metalloproteinases (ADAMs), and ADAMs with thrombospondin motifs (ADAMTSs).
MMPs induced by pro- and inflammatory factors and regulated by NF-κB [23], as mentioned above, have an important role in sustaining all six major hallmarks of cancer [24] highlighted within the complexity of cancer biology: (1) sustaining proliferative signaling; (2) evasion of apoptosis; (3) angiogenesis and lymphangiogenesis; (4) invasion and metastasis; (5) reprogramming of energy metabolism; and (6) evasion of the immune response. Indeed, there is a close relationship between the release of cytokines and the specific ECM-degrading enzymes MMPs, as MMPs may play a non-proteolytic and non-ECM role in cell–cell communication [25] (Figure 1). For a deeper description of the role of specific MMPs regulated by cytokines, see Review [25].
2. Extracellular Chaperones in the TME: Focus on Their Roles in Cancer-Related Inflammation and ECM Remodeling
The protein homeostasis of cancer cells is challenged by an increased rate of protein synthesis that is required to face the anabolic demand of fast-growing and proliferating cells. Oxygen and nutrients limitations drive changes in the metabolism of cancer cells that contribute to the maintenance of an acidified, oxidized, and inflamed TME favoring the unfolding of many extracellular proteins [26].
As a consequence, cancer cells upregulate the activity of their protein quality systems that collectively constitute the proteostasis network in the attempt to reduce the proteotoxic stress and keep the proteome as stable and functional as possible [27].
Molecular chaperones are first-line players of the proteostasis network due to their ubiquitous expression and their ability to stabilize proteins in their native conformation, re-fold denatured polypeptides, inhibit unfolded protein aggregation, and direct misfolded proteins to proteasomal and autophagic degradation [28][29]. The wide and heterogeneous family of molecular chaperones comprises, among others, the heat shock proteins (HSPs), a family of stress-proteins that are induced in response to physical, chemical, and biological insults to support cell survival [30]. According to their molecular weight, HSPs are currently classified into six groups. HSP100s, HSP90s, HSP70s, and HSP60s use the energy derived from ATP hydrolysis to assist protein folding. HSP40s and other small HSPs do not possess ATPase activity and act as a co-chaperone to HSP70s, binding to unfolded proteins preventing their aggregation and eventually delivering those irreparably damaged to the protein degradation systems [31].
It is now widely accepted that many HSPs, initially classified as intracellular proteins, can reach the extracellular space through various mechanisms, including exocytosis of lysosomal vesicles, translocation through the plasma membrane, and release of extracellular vesicles as exosomes [32][33][34][35].
The function of extracellular HSPs (eHSP) in the TME is not limited to the canonical function of stress proteins that counteract protein unfolding (reviewed in Richter K. et al.) [36]. More recent studies highlighted that eHSPs take part in the complex signaling network that influences the fate of cancer cells with different outcomes depending on the stage of the pathology [30]. They counteract tumor growth due to their ability to bind antigenic peptides and activate anti-tumor innate immunity [37], thus promoting immune surveillance and inhibiting inflammation [38]. However, they favor tumor progression and metastasis formation by sustaining cancer proliferation, cell migration and invasion, neo-angiogenesis, and immune escape [30].
The eHSPs interact directly with cell surface receptors and consequently engage intracellular signaling pathways that influence cell behavior [30]. Low-density lipoprotein receptor-related protein 1 (LRP1) is a common receptor for many extracellular chaperones, both in normal and cancer cells where its expression is often upregulated [39]. eHSP90 following LRP1 binding activates both AKT and NF-κB signaling, promoting cell proliferation, EMT, and ECM remodeling [40][41]. Similarly, various eHSPs activate Toll-like receptors (TLRs), induce pro-inflammatory signaling cascades through NF-κB and ERK activation, and promote tumor establishment releasing pro-inflammatory, anti-apoptotic, proliferative, and pro-fibrogenic signals on cancer cells and TME cells [42].
Many ECM-degrading enzymes depend on eHSPs binding for their activity and stability. For instance, in breast cancer, eHSP90α mediates the proteolytic activation of MMP2 in cooperation with eHSP70 and other co-chaperones [43]. eHSP90α interacts with lysyl oxidase-like 2 (LOXL2) and allows the latter to assume its functional conformation favoring the migration of breast cancer cells [44] and with tissue plasminogen activator (tPA), favoring the conversion of plasminogen in plasmin, a process that favors cancer invasion [45]. In breast cancer cells, both HSP90α and HSP90β are closely associated in a complex with fibronectin, participating in its assembly and turnover [46][47].
Clusterin: A Prominent Extracellular Chaperone in the TME
Clusterin (CLU) is a heterodimeric glycoprotein of 80 kDa, secreted in almost all biological fluids, first identified in ram rete testis where it showed signs of clustering with rat sertoli cells and erythrocytes [48]. CLU is involved in a plethora of fundamental cellular processes and has been recently proposed as one of the most prominent extracellular ATP-independent chaperones, functionally related to the sHSPs, exhibiting anti-inflammatory functions [49][50].
Unlike other well-known HSPs, CLU is synthesized by ribosomes attached to the ER and reaches the outside of the cells by the canonical ER–Golgi route [51]. Once in the extracellular compartment, CLU binds to a variety of structurally unrelated client molecules through a dynamic disordered region containing amphipathic α-helices [52]. Of note, CLU not only prevents their aggregation but also commits them to lysosomal-degradation through endocytosis mediated by surface receptors [53]. A more recent study identified heparan sulfate (HS) as the main clearance receptor involved in CLU-client protein complex endocytosis and subsequent lysosomal degradation. Of note, the CLU-HS pathway may represent a ubiquitous and versatile degradation pathway for different extracellular damaged or unwanted ligands [54]. By these combinations of binding and scavenging properties, CLU participates in the reestablishment of the extracellular protein homeostasis [55].
According to its function, CLU has been involved in many diseases that are accompanied by an impairment in protein synthesis and protein quality maintenance, including neurodegeneration, inflammatory diseases, and cancer [56][57][58][59]. Many cancer- and inflammation-related transcription factors, as NF-κB, HIF1α, TGFβ, TNFα are involved in the transcriptional regulation of CLU during cancer progression [60]. CLU is down-regulated in the vast majority of primary and naïve cancers in comparison to normal tissue [59][60][61] by epigenetic mechanisms that include CpG islands hypermethylation and histone tail modification [62][63][64][65][66][67]. Conversely, CLU expression is upregulated in the late stages of carcinogenesis, especially in niches of cancer cells resistant to chemotherapy or hormonal therapy [68].
In line with the observations reported above, the abrogation of CLU expression in mouse models of neuroblastoma, prostate, lung, and skin carcinogenesis leads to a quicker progression of the neoplasia toward the invasive phenotype [69][70][71][72], while targeting CLU with siRNA or antisense oligonucleotides enhances the cytotoxicity of chemotherapeutic drugs, ionizing radiation, and hormonal therapy [73][74].
The apparent contradictory tumor limiting and promoting functions of CLU represents instead the two sides of the same coin, which are consistent with a cytoprotective role exerted by the protein, both under normal and pathological conditions. Actually, at early stages of carcinogenesis, CLU expression is required to counteract the proteotoxic stress after increased anabolism that in turn drives genomic instability onset. Differently, under the selective pressure of cytotoxic therapies and altered signal transduction, CLU is over-expressed by resistant cells and favors the onset of a permissive tumor microenvironment. Some of the functions played by CLU in the inflamed TME are illustrated in Figure 2.
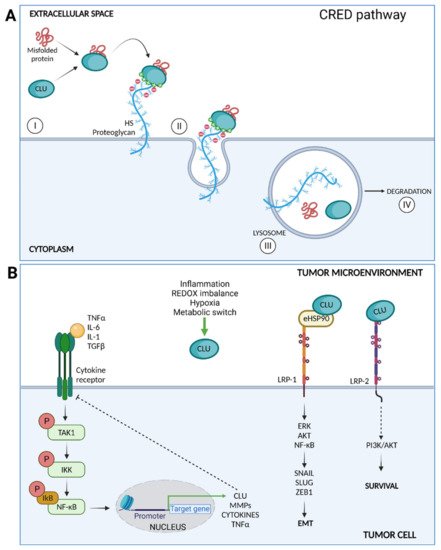
Figure 2. (A) CLU mediates clearance of misfolded proteins through the chaperone- and receptor-mediated extracellular protein degradation (CRED) pathway. CLU binds a misfolded protein and then interacts with the HS chains membrane PGs by electrostatic interactions (I); the CLU–client protein-receptor is endocytosed (II) and delivered to the lysosome (III) for intracellular degradation (IV). (B) Intracellular signaling pathways triggered by CLU after surface receptors binding. CLU inhibits the TAK1– NF-κB signaling axis triggered by inflammatory cytokines. CLU facilitates eHSP90 binding to LRP1, enhancing the activation of ERK, AKT, and NF-κB and promoting EMT. Finally, CLU binds LRP2 and promotes cell survival through PI3K/AKT activation.
In this scenario, different studies have shown that increased levels of CLU correlate with the induction of EMT in various cancer cell lines [75][76][77][78][79]. Coherently, CLU silencing is accompanied by an increase in epithelial markers as E-cadherin and junctional zona occludens-1 (ZO-1) concomitantly with the loss of the mesenchymal markers fibronectin, N-cadherin, and vimentin [75][76][77][78][79]. CLU is one of the most significantly upregulated genes in the murine breast cancer cell line BRI-JM01, undergoing TGFβ-induced EMT, where its neutralization with an anti-CLU antibody blocks the acquisition of the mesenchymal phenotype induced by TGFβ [75]. The positive modulatory effects of TGFβ on CLU transcription is mediated by Twist-1, an EMT-promoting transcription factor [77]. In hepatocellular carcinoma cells, CLU silencing leads to a reduction in Smad3 phosphorylation in concomitance with the inhibition of EMT markers [78]. This finding suggests that CLU in the meantime is a transcriptional target of TGFβ and positively modulates the TGFβ signaling transduction through the canonical Smad pathway. In breast cancer cells, CLU favors the binding of eHSP-90α to the surface receptor LRP-1 and potentiates the transduction of EMT-promoting signaling [79].
In prostate, breast, and colon cancer, the expression of CLU changes with disease progression both in the epithelial cancer cells and in the surrounding stromal tissue [80][81][82]. Of note, only clusterin stromal staining significantly correlates with disease recurrence and clinical outcome following surgical intervention or chemotherapy [82][83].
These results highlight the concept that stromal non-cancer cells contribute to shaping the biological behavior of cancer cells, strongly suggesting a role of CLU in ECM remodeling. Disappointingly, so far only a limited number of studies have specifically addressed this issue. In vitro studies conducted in breast, renal, nasopharyngeal, and hepatocellular cancer cell lines found that MMP-9 and MMP-2 expression is reduced following CLU silencing, concomitantly with a decrease in cell motility and invasion [84][85][86][87].
Collectively, all these studies limited their investigation to evaluating the changes of MMPs expression in response to CLU gene manipulation (up- or down-regulation), without paralleling gene expression data with measures of the enzymatic activity. In contrast, experiments performed in normal kidney cells, in neutrophils, and in breast cancer cell line MCF-7 found that CLU binds to and inhibits the activity of MT6-MMP, a member of the membrane-type MMPs [88]. An inhibiting effect of CLU on MMPs activity was also confirmed in the context of dry eye syndrome, an inflammatory disease caused by inadequate hydration and lubrication of the ocular surface. In a mouse model of dry eye disease, the induction of the pathology is accompanied by a decrease in CLU expression and a concomitant increase in MMPs expression. The authors demonstrated that CLU prevented stress-induced MMP-9 aggregation and activation, both inside and outside human epithelial cells, by direct binding with the catalytic domain. The inhibition of the enzymatic activity was also demonstrated for MMP-2, MMP-3, and MMP-7 [89]. In addition, TNFα stimulation of corneal cultured cells induced CLU upregulation, along with a decrease in MMP-9 expression, while the topical administration of CLU protected the ocular surface barrier from functional disruption after dry eye induction in a mouse model [90]. These data suggest a role for CLU in preventing the inflammatory status of the eye maintaining the normal fluid/epithelial interface homeostasis.
In support of the anti-inflammatory function of CLU, our laboratory recently showed that CLU limits NF-κB hyperactivation in prostate cancer cells and pre-clinical models of prostate cancer [91]. More precisely, CLU overexpression in PC3 cells is accompanied by a significant reduction of the phosphorylation of the p65 subunit of NF-κB. This led to reduced nuclear translocation of NF-κB along with a reduction of MMP-2 and MMP-9, while CLU silencing in the same cells produced opposite effects. In line with these data, we found a significant increase of MMP-2 and MMP-9 gelatinase activity in a transgenic murine model of prostate cancer knock-out for CLU (TRAMP/CLUKO) compared with age-matched prostate cancer-prone mice (TRAMP) expressing CLU [91]. Recently, another study showed that CLU inactivation leads to the constitutive activation of the TAK1–NF-κB signaling axis in experimental models of non-small cell lung cancer and promotes EMT in vitro and metastasis formation in vivo [70].
Altogether, these data suggest that CLU expression may be required to limit the excessive activation of NF-κB that occurs during chronic inflammation associated with tumor growth. The inhibition of the MMPs activity, therefore, would be part of an array of anti-inflammatory properties that CLU shares with other members of the sHSPs. In this context, the abrogation of CLU expression at early stages of carcinogenesis would fail to dampen NF-κB activity resulting in the generation of a TME conducive to the process of tumor invasion.
References
- Dvorak, H.F. Tumors: Wounds that do not heal. Similarities between tumor stroma generation and wound healing. N. Engl. J. Med. 1986, 315, 1650–1659.
- Grivennikov, S.I.; Greten, F.R.; Karin, M. Immunity, inflammation, and cancer. Cell 2010, 140, 883–899.
- Massague, J. Tgfbeta in Cancer. Cell 2008, 134, 215–230.
- Hargadon, K.M. Dysregulation of TGFbeta1 Activity in Cancer and Its Influence on the Quality of Anti-Tumor Immunity. J. Clin. Med. 2016, 5, 76.
- Pietras, K.; Ostman, A. Hallmarks of cancer: Interactions with the tumor stroma. Exp. Cell Res. 2010, 316, 1324–1331.
- Polanska, U.M.; Orimo, A. Carcinoma-associated fibroblasts: Non-neoplastic tumour-promoting mesenchymal cells. J. Cell. Physiol. 2013, 228, 1651–1657.
- Kalluri, R. The biology and function of fibroblasts in cancer. Nat. Rev. Cancer 2016, 16, 582–598.
- Olumi, A.F.; Grossfeld, G.D.; Hayward, S.W.; Carroll, P.R.; Tlsty, T.D.; Cunha, G.R. Carcinoma-associated fibroblasts direct tumor progression of initiated human prostatic epithelium. Cancer Res. 1999, 59, 5002–5011.
- Yoshida, G.J.; Azuma, A.; Miura, Y.; Orimo, A. Activated Fibroblast Program Orchestrates Tumor Initiation and Progression; Molecular Mechanisms and the Associated Therapeutic Strategies. Int. J. Mol. Sci. 2019, 20, 2256.
- Foster, D.S.; Jones, R.E.; Ransom, R.C.; Longaker, M.T.; Norton, J.A. The evolving relationship of wound healing and tumor stroma. JCI Insight 2018, 3.
- Heikkila, K.; Ebrahim, S.; Lawlor, D.A. Systematic review of the association between circulating interleukin-6 (IL-6) and cancer. Eur. J. Cancer 2008, 44, 937–945.
- Landskron, G.; De la Fuente, M.; Thuwajit, P.; Thuwajit, C.; Hermoso, M.A. Chronic inflammation and cytokines in the tumor microenvironment. J. Immunol. Res. 2014, 2014, 149185.
- Hugo, H.J.; Lebret, S.; Tomaskovic-Crook, E.; Ahmed, N.; Blick, T.; Newgreen, D.F.; Thompson, E.W.; Ackland, M.L. Contribution of Fibroblast and Mast Cell (Afferent) and Tumor (Efferent) IL-6 Effects within the Tumor Microenvironment. Cancer Microenviron. 2012, 5, 83–93.
- Balkwill, F. TNF-alpha in promotion and progression of cancer. Cancer Metastasis Rev. 2006, 25, 409–416.
- Balkwill, F.; Mantovani, A. Inflammation and cancer: Back to Virchow? Lancet 2001, 357, 539–545.
- Li, B.; Vincent, A.; Cates, J.; Brantley-Sieders, D.M.; Polk, D.B.; Young, P.P. Low levels of tumor necrosis factor alpha increase tumor growth by inducing an endothelial phenotype of monocytes recruited to the tumor site. Cancer Res. 2009, 69, 338–348.
- Charles, K.A.; Kulbe, H.; Soper, R.; Escorcio-Correia, M.; Lawrence, T.; Schultheis, A.; Chakravarty, P.; Thompson, R.G.; Kollias, G.; Smyth, J.F.; et al. The tumor-promoting actions of TNF-alpha involve TNFR1 and IL-17 in ovarian cancer in mice and humans. J. Clin. Investig. 2009, 119, 3011–3023.
- Winkler, J.; Abisoye-Ogunniyan, A.; Metcalf, K.J.; Werb, Z. Concepts of extracellular matrix remodelling in tumour progression and metastasis. Nat. Commun. 2020, 11, 5120.
- Girigoswami, K.; Saini, D.; Girigoswami, A. Extracellular Matrix Remodeling and Development of Cancer. Stem Cell Rev. Rep. 2020, 739–747.
- Conklin, M.W.; Eickhoff, J.C.; Riching, K.M.; Pehlke, C.A.; Eliceiri, K.W.; Provenzano, P.P.; Friedl, A.; Keely, P.J. Aligned collagen is a prognostic signature for survival in human breast carcinoma. Am. J. Pathol. 2011, 178, 1221–1232.
- Tenti, P.; Vannucci, L. Lysyl oxidases: Linking structures and immunity in the tumor microenvironment. Cancer Immunol. Immunother. 2020, 69, 223–235.
- Ben-Neriah, Y.; Karin, M. Inflammation meets cancer, with NF-kappaB as the matchmaker. Nat. Immunol. 2011, 12, 715–723.
- Bassères, D.S.; Baldwin, A.S. Nuclear factor-kappaB and inhibitor of kappaB kinase pathways in oncogenic initiation and progression. Oncogene 2006, 25, 6817–6830.
- Caon, I.; Bartolini, B.; Parnigoni, A.; Carava, E.; Moretto, P.; Viola, M.; Karousou, E.; Vigetti, D.; Passi, A. Revisiting the hallmarks of cancer: The role of hyaluronan. Semin. Cancer Biol. 2020, 62, 9–19.
- Young, D.; Das, N.; Anowai, A.; Dufour, A. Matrix Metalloproteases as Influencers of the Cells’ Social Media. Int. J. Mol. Sci. 2019, 20, 3847.
- Ciocca, D.R.; Arrigo, A.P.; Calderwood, S.K. Heat shock proteins and heat shock factor 1 in carcinogenesis and tumor development: An update. Arch. Toxicol. 2013, 87, 19–48.
- Hartl, F.U.; Bracher, A.; Hayer-Hartl, M. Molecular chaperones in protein folding and proteostasis. Nature 2011, 475, 324–332.
- Wyatt, A.R.; Yerbury, J.J.; Ecroyd, H.; Wilson, M.R. Extracellular Chaperones and Proteostasis. Annu. Rev. Biochem. 2013, 82, 295–322.
- Clausen, L.; Abildgaard, A.B.; Gersing, S.K.; Stein, A.; Lindorff-Larsen, K.; Hartmann-Petersen, R. Protein Stability and Degradation in Health and Disease. Adv. Protein Chem. Struct. Biol. 2019, 114, 61–83.
- Secli, L.; Fusella, F.; Avalle, L.; Brancaccio, M. The Dark-Side of the Outside: How Extracellular Heat Shock Proteins Promote Cancer. Cell Mol. Life Sci. 2021, 4069–4083.
- Kampinga, H.H.; Hageman, J.; Vos, M.J.; Kubota, H.; Tanguay, R.M.; Bruford, E.A.; Cheetham, M.E.; Chen, B.; Hightower, L.E. Guidelines for the Nomenclature of the Human Heat Shock Proteins. Cell Stress Chaperones 2009, 14, 105–111.
- Basu, S.; Binder, R.J.; Suto, R.; Anderson, K.M.; Srivastava, P.K. Necrotic but Not Apoptotic Cell Death Releases Heat Shock Proteins, Which Deliver a Partial Maturation Signal to Dendritic Cells and Activate the Nf-Kappa B Pathway. Int. Immunol. 2000, 12, 1539–1546.
- Santos, T.G.; Martins, V.R.; Hajj, G.N.M. Unconventional Secretion of Heat Shock Proteins in Cancer. Int. J. Mol. Sci. 2017, 18, 946.
- Mambula, S.S.; Calderwood, S.K. Heat Shock Protein 70 Is Secreted from Tumor Cells by a Nonclassical Pathway Involving Lysosomal Endosomes. J. Immunol. 2006, 177, 7849–7857.
- Albakova, Z.; Siam, M.K.S.; Sacitharan, P.K.; Ziganshin, R.H.; Ryazantsev, D.Y.; Sapozhnikov, A.M. Extracellular Heat Shock Proteins and Cancer: New Perspectives. Transl. Oncol. 2021, 14, 100995.
- Richter, K.; Haslbeck, M.; Buchner, J. The heat shock response: Life on the verge of death. Mol. Cell 2010, 40, 253–266.
- Suto, R.; Srivastava, P.K. A Mechanism for the Specific Immunogenicity of Heat Shock Protein-Chaperoned Peptides. Science 1995, 269, 1585–1588.
- Taha, E.A.; Ono, K.; Eguchi, T. Roles of Extracellular Hsps as Biomarkers in Immune Surveillance and Immune Evasion. Int. J. Mol. Sci. 2019, 20, 4588.
- Basu, S.; Binder, R.J.; Ramalingam, T.; Srivastava, P.K. Cd91 Is a Common Receptor for Heat Shock Proteins Gp96, Hsp90, Hsp70, and Calreticulin. Immunity 2001, 14, 303–313.
- Tsen, F.; Bhatia, A.; O’Brien, K.; Cheng, C.F.; Chen, M.; Hay, N.; Stiles, B.; Woodley, D.T.; Li, W. Extracellular Heat Shock Protein 90 Signals through Subdomain Ii and the Npvy Motif of Lrp-1 Receptor to Akt1 and Akt2: A Circuit Essential for Promoting Skin Cell Migration in Vitro and Wound Healing in Vivo. Mol. Cell Biol. 2013, 33, 4947–4959.
- Chen, W.S.; Chen, C.C.; Chen, L.L.; Lee, C.C.; Huang, T.S. Secreted Heat Shock Protein 90alpha (Hsp90alpha) Induces Nuclear Factor-Kappab-Mediated Tcf12 Protein Expression to Down-Regulate E-Cadherin and to Enhance Colorectal Cancer Cell Migration and Invasion. J. Biol. Chem. 2013, 288, 9001–9010.
- Pradere, J.P.; Dapito, D.H.; Schwabe, R.F. The Yin and Yang of Toll-Like Receptors in Cancer. Oncogene 2014, 33, 3485–3495.
- Sims, J.D.; McCready, J.; Jay, D.G. Extracellular Heat Shock Protein (Hsp)70 and Hsp90alpha Assist in Matrix Metalloproteinase-2 Activation and Breast Cancer Cell Migration and Invasion. PLoS ONE 2011, 6, e18848.
- McCready, J.; Wong, D.S.; Burlison, J.A.; Ying, W.; Jay, D.G. An Impermeant Ganetespib Analog Inhibits Extracellular Hsp90-Mediated Cancer Cell Migration That Involves Lysyl Oxidase 2-Like Protein. Cancers 2014, 6, 1031–1046.
- McCready, J.; Sims, J.D.; Chan, D.; Jay, D.G. Secretion of extracellular hsp90alpha via exosomes increases cancer cell motility: A role for plasminogen activation. BMC Cancer 2010, 10, 294.
- Hunter, M.C.; O’Hagan, K.L.; Kenyon, A.; Dhanani, K.C.; Prinsloo, E.; Edkins, A.L. Hsp90 Binds Directly to Fibronectin (Fn) and Inhibition Reduces the Extracellular Fibronectin Matrix in Breast Cancer Cells. PLoS ONE 2014, 9, e86842.
- Chakraborty, A.; Boel, N.M.; Edkins, A.L. Hsp90 Interacts with the Fibronectin N-Terminal Domains and Increases Matrix Formation. Cells 2020, 9, 272.
- Blaschuk, O.; Burdzy, K.; Fritz, I.B. Purification and Characterization of a Cell-Aggregating Factor (Clusterin), the Major Glycoprotein in Ram Rete Testis Fluid. J. Biol. Chem. 1983, 258, 7714–7720.
- Poon, S.; Treweek, T.M.; Wilson, M.R.; Easterbrook-Smith, S.B.; Carver, J.A. Clusterin Is an Extracellular Chaperone That Specifically Interacts with Slowly Aggregating Proteins on Their Off-Folding Pathway. FEBS Lett. 2002, 513, 259–266.
- Wilson, M.R.; Easterbrook-Smith, S.B. Clusterin Is a Secreted Mammalian Chaperone. Trends Biochem. Sci. 2000, 25, 95–98.
- Rohne, P.; Prochnow, H.; Koch-Brandt, C. The Clu-Files: Disentanglement of a Mystery. Biomol. Concepts 2016, 7, 1–15.
- Bailey, R.W.; Dunker, A.K.; Brown, C.J.; Garner, E.C.; Griswold, M.D. Clusterin, a Binding Protein with a Molten Globule-Like Region. Biochemistry 2001, 40, 11828–11840.
- Wyatt, A.R.; Yerbury, J.J.; Berghofer, P.; Greguric, I.; Katsifis, A.; Dobson, C.M.; Wilson, M.R. Clusterin Facilitates in Vivo Clearance of Extracellular Misfolded Proteins. Cell Mol. Life Sci. 2011, 68, 3919–3931.
- Itakura, E.; Chiba, M.; Murata, T.; Matsuura, A. Heparan Sulfate Is a Clearance Receptor for Aberrant Extracellular Proteins. J. Cell Biol. 2020, 219.
- Trougakos, I.P. The Molecular Chaperone Apolipoprotein J/Clusterin as a Sensor of Oxidative Stress: Implications in Therapeutic Approaches—A Mini-Review. Gerontology 2013, 59, 514–523.
- Foster, E.M.; Dangla-Valls, A.; Lovestone, S.; Ribe, E.M.; Buckley, N.J. Clusterin in Alzheimer’s Disease: Mechanisms, Genetics, and Lessons from Other Pathologies. Front. Neurosci. 2019, 13, 164.
- Fini, M.E.; Bauskar, A.; Jeong, S.; Wilson, M.R. Clusterin in the Eye: An Old Dog with New Tricks at the Ocular Surface. Exp. Eye Res. 2016, 147, 57–71.
- Niforou, K.; Cheimonidou, C.; Trougakos, I.P. Molecular Chaperones and Proteostasis Regulation During Redox Imbalance. Redox Biol. 2014, 2, 323–332.
- Rizzi, F.; Bettuzzi, S. The Clusterin Paradigm in Prostate and Breast Carcinogenesis. Endocr. Relat. Cancer 2010, 17, R1–R17.
- Sala, A.; Bettuzzi, S.; Pucci, S.; Chayka, O.; Dews, M.; Thomas-Tikhonenko, A. Regulation of Clu Gene Expression by Oncogenes and Epigenetic Factors Implications for Tumorigenesis. Adv. Cancer Res. 2009, 105, 115–132.
- Cheimonidi, C.; Grivas, I.N.; Sesti, F.; Kavrochorianou, N.; Gianniou, D.D.; Taoufik, E.; Badounas, F.; Papassideri, I.; Rizzi, F.; Tsitsilonis, O.E.; et al. Clusterin Overexpression in Mice Exacerbates Diabetic Phenotypes but Suppresses Tumor Progression in a Mouse Melanoma Model. Aging 2021, 13, 6485–6505.
- Wang, C.; Liu, Z.; Woo, C.W.; Li, Z.; Wang, L.; Wei, J.S.; Marquez, V.E.; Bates, S.E.; Jin, Q.; Khan, J.; et al. Ezh2 Mediates Epigenetic Silencing of Neuroblastoma Suppressor Genes Casz1, Clu, Runx3, and Ngfr. Cancer Res. 2012, 72, 315–324.
- Bonacini, M.; Coletta, M.; Ramazzina, I.; Naponelli, V.; Modernelli, A.; Davalli, P.; Bettuzzi, S.; Rizzi, F. Distinct Promoters, Subjected to Epigenetic Regulation, Drive the Expression of Two Clusterin Mrnas in Prostate Cancer Cells. Biochim. Biophys. Acta 2015, 1849, 44–54.
- Rauhala, H.E.; Porkka, K.P.; Saramaki, O.R.; Tammela, T.L.; Visakorpi, T. Clusterin Is Epigenetically Regulated in Prostate Cancer. Int. J. Cancer 2008, 123, 1601–1609.
- Serrano, A.; Redondo, M.; Tellez, T.; Castro-Vega, I.; Roldan, M.J.; Mendez, R.; Rueda, A.; Jimenez, E. Regulation of Clusterin Expression in Human Cancer Via DNA Methylation. Tumour Biol. 2009, 30, 286–291.
- Corvetta, D.; Chayka, O.; Gherardi, S.; D’Acunto, C.W.; Cantilena, S.; Valli, E.; Piotrowska, I.; Perini, G.; Sala, A. Physical Interaction between Mycn Oncogene and Polycomb Repressive Complex 2 (Prc2) in Neuroblastoma: Functional and Therapeutic Implications. J. Biol. Chem. 2013, 288, 8332–8341.
- Hellebrekers, D.M.; Melotte, V.; Vire, E.; Langenkamp, E.; Molema, G.; Fuks, F.; Herman, J.G.; van Criekinge, W.; Griffioen, A.W.; van Engeland, M. Identification of Epigenetically Silenced Genes in Tumor Endothelial Cells. Cancer Res. 2007, 67, 4138–4148.
- Zhou, J.; Chen, X.; Gilvary, D.L.; Tejera, M.M.; Eksioglu, E.A.; Wei, S.; Djeu, J.Y. Hmgb1 Induction of Clusterin Creates a Chemoresistant Niche in Human Prostate Tumor Cells. Sci. Rep. 2015, 5, 15085.
- Bettuzzi, S.; Davalli, P.; Davoli, S.; Chayka, O.; Rizzi, F.; Belloni, L.; Pellacani, D.; Fregni, G.; Astancolle, S.; Fassan, M.; et al. Genetic Inactivation of Apoj/Clusterin: Effects on Prostate Tumourigenesis and Metastatic Spread. Oncogene 2009, 28, 4344–4352.
- Chen, Z.; Fan, Z.; Dou, X.; Zhou, Q.; Zeng, G.; Liu, L.; Chen, W.; Lan, R.; Liu, W.; Ru, G.; et al. Inactivation of Tumor Suppressor Gene Clusterin Leads to Hyperactivation of Tak1-Nf-Kappab Signaling Axis in Lung Cancer Cells and Denotes a Therapeutic Opportunity. Theranostics 2020, 10, 11520–11534.
- Chayka, O.; Corvetta, D.; Dews, M.; Caccamo, A.E.; Piotrowska, I.; Santilli, G.; Gibson, S.; Sebire, N.J.; Himoudi, N.; Hogarty, M.D.; et al. Clusterin, a Haploinsufficient Tumor Suppressor Gene in Neuroblastomas. J. Natl. Cancer Inst. 2009, 101, 663–677.
- Thomas-Tikhonenko, A.; Viard-Leveugle, I.; Dews, M.; Wehrli, P.; Sevignani, C.; Yu, D.; Ricci, S.; el-Deiry, W.; Aronow, B.; Kaya, G.; et al. Myc-Transformed Epithelial Cells Down-Regulate Clusterin, Which Inhibits Their Growth in Vitro and Carcinogenesis in Vivo. Cancer Res. 2004, 64, 3126–3136.
- Djeu, J.Y.; Wei, S. Clusterin and Chemoresistance. Adv. Cancer Res. 2009, 105, 77–92.
- Wilson, M.R.; Zoubeidi, A. Clusterin as a Therapeutic Target. Expert Opin. Targets 2017, 21, 201–213.
- Lenferink, A.E.; Cantin, C.; Nantel, A.; Wang, E.; Durocher, Y.; Banville, M.; Paul-Roc, B.; Marcil, A.; Wilson, M.R.; O’Connor-McCourt, M.D. Transcriptome Profiling of a Tgf-Beta-Induced Epithelial-to-Mesenchymal Transition Reveals Extracellular Clusterin as a Target for Therapeutic Antibodies. Oncogene 2010, 29, 831–844.
- Chou, T.Y.; Chen, W.C.; Lee, A.C.; Hung, S.M.; Shih, N.Y.; Chen, M.Y. Clusterin Silencing in Human Lung Adenocarcinoma Cells Induces a Mesenchymal-to-Epithelial Transition through Modulating the Erk/Slug Pathway. Cell Signal. 2009, 21, 704–711.
- Shiota, M.; Zardan, A.; Takeuchi, A.; Kumano, M.; Beraldi, E.; Naito, S.; Zoubeidi, A.; Gleave, M.E. Clusterin Mediates Tgf-Beta-Induced Epithelial-Mesenchymal Transition and Metastasis Via Twist1 in Prostate Cancer Cells. Cancer Res. 2012, 72, 5261–5272.
- Wang, C.; Jiang, K.; Kang, X.; Gao, D.; Sun, C.; Li, Y.; Sun, L.; Zhang, S.; Liu, X.; Wu, W.; et al. Tumor-Derived Secretory Clusterin Induces Epithelial-Mesenchymal Transition and Facilitates Hepatocellular Carcinoma Metastasis. Int. J. Biochem. Cell Biol. 2012, 44, 2308–2320.
- Tian, Y.; Wang, C.; Chen, S.; Liu, J.; Fu, Y.; Luo, Y. Extracellular Hsp90alpha and Clusterin Synergistically Promote Breast Cancer Epithelial-to-Mesenchymal Transition and Metastasis Via Lrp1. J. Cell Sci. 2019, 132, jcs228213.
- Scaltriti, M.; Brausi, M.; Amorosi, A.; Caporali, A.; D’Arca, D.; Astancolle, S.; Corti, A.; Bettuzzi, S. Clusterin (Sgp-2, Apoj) Expression Is Downregulated in Low- and High-Grade Human Prostate Cancer. Int. J. Cancer 2004, 108, 23–30.
- Andersen, C.L.; Schepeler, T.; Thorsen, K.; Birkenkamp-Demtroder, K.; Mansilla, F.; Aaltonen, L.A.; Laurberg, S.; Orntoft, T.F. Clusterin Expression in Normal Mucosa and Colorectal Cancer. Mol. Cell Proteom. 2007, 6, 1039–1048.
- Wang, Y.; Brodsky, A.S.; Xiong, J.; Lopresti, M.L.; Yang, D.; Resnick, M.B. Stromal Clusterin Expression Predicts Therapeutic Response to Neoadjuvant Chemotherapy in Triple Negative Breast Cancer. Clin. Breast Cancer 2018, 18, e373–e379.
- Pins, M.R.; Fiadjoe, J.E.; Korley, F.; Wong, M.; Rademaker, A.W.; Jovanovic, B.; Yoo, T.K.; Kozlowski, J.M.; Raji, A.; Yang, X.J.; et al. Clusterin as a Possible Predictor for Biochemical Recurrence of Prostate Cancer Following Radical Prostatectomy with Intermediate Gleason Scores: A Preliminary Report. Prostate Cancer Prostatic Dis. 2004, 7, 243–248.
- Li, J.; Jia, L.; Zhao, P.; Jiang, Y.; Zhong, S.; Chen, D. Stable Knockdown of Clusterin by Vectorbased Rna Interference in a Human Breast Cancer Cell Line Inhibits Tumour Cell Invasion and Metastasis. J. Int. Med. Res. 2012, 40, 545–555.
- Wang, X.; Luo, L.; Dong, D.; Yu, Q.; Zhao, K. Clusterin Plays an Important Role in Clear Renal Cell Cancer Metastasis. Urol. Int. 2014, 92, 95–103.
- Zheng, W.; Yao, M.; Wu, M.; Yang, J.; Yao, D.; Wang, L. Secretory Clusterin Promotes Hepatocellular Carcinoma Progression by Facilitating Cancer Stem Cell Properties Via Akt/Gsk-3beta/Beta-Catenin Axis. J. Transl. Med. 2020, 18, 81.
- Zhong, J.; Yu, X.; Dong, X.; Lu, H.; Zhou, W.; Li, L.; Li, Z.; Sun, P.; Shi, X. Therapeutic Role of Meloxicam Targeting Secretory Clusterin-Mediated Invasion in Hepatocellular Carcinoma Cells. Oncol. Lett. 2018, 15, 7191–7199.
- Matsuda, A.; Itoh, Y.; Koshikawa, N.; Akizawa, T.; Yana, I.; Seiki, M. Clusterin, an Abundant Serum Factor, Is a Possible Negative Regulator of Mt6-Mmp/Mmp-25 Produced by Neutrophils. J. Biol. Chem. 2003, 278, 36350–36357.
- Jeong, S.; Ledee, D.R.; Gordon, G.M.; Itakura, T.; Patel, N.; Martin, A.; Fini, M.E. Interaction of Clusterin and Matrix Metalloproteinase-9 and Its Implication for Epithelial Homeostasis and Inflammation. Am. J. Pathol. 2012, 180, 2028–2039.
- Bauskar, A.; Mack, W.J.; Mauris, J.; Argueso, P.; Heur, M.; Nagel, B.A.; Kolar, G.R.; Gleave, M.E.; Nakamura, T.; Kinoshita, S.; et al. Clusterin Seals the Ocular Surface Barrier in Mouse Dry Eye. PLoS ONE 2015, 10, e0138958.
- Bonacini, M.; Negri, A.; Davalli, P.; Naponelli, V.; Ramazzina, I.; Lenzi, C.; Bettuzzi, S.; Rizzi, F. Clusterin Silencing in Prostate Cancer Induces Matrix Metalloproteinases by an Nf-Kappab-Dependent Mechanism. J. Oncol. 2019, 2019, 4081624.

