 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Ifeanyi D. Nwachukwu | + 6160 word(s) | 6160 | 2021-08-17 04:04:50 | | | |
| 2 | Bruce Ren | -21 word(s) | 6139 | 2021-08-19 07:39:29 | | |
Video Upload Options
Antioxidants remain interesting molecules of choice for suppression of the toxic effects of free radicals in foods and human systems. The current practice involves the use of mainly synthetic molecules as potent antioxidant agents. However, due to the potential negative impact on human health, there is an intensive effort within the research community to develop natural alternatives with similar antioxidant efficacy but without the negative side effects of synthetic molecules. Still, the successful development of new molecules depends on the use of reliable chemical or cell culture assays to screen antioxidant properties. Chemical antioxidant assays include the determination of scavenging ability against free radicals such as DPPH, superoxide anion radicals, hydroxyl radicals, hydrogen peroxide, and nitric oxide. Other antioxidant tests include the ability of compounds to bind and sequester prooxidant metal cations, reduce ferric iron, and attenuate the rate of lipid oxidation. Ex vivo tests utilize cell cultures to confirm entry of the molecules into cells and the ability to quench synthetic intracellular free radicals or to stimulate the increased biosynthesis of endogenous antioxidants.
1. Introduction
2. Principles of Chemical Antioxidant Assays
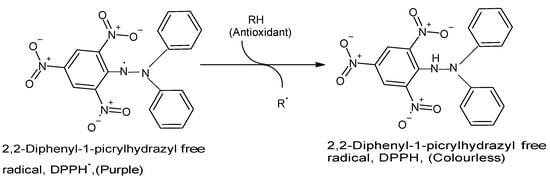
2.1. Free Radical (Synthetic DPPH) Scavenging
2.2. ROS Trapping (Hydrogen Peroxide, Nitric Oxide, Also Superoxide and Hydroxyl)
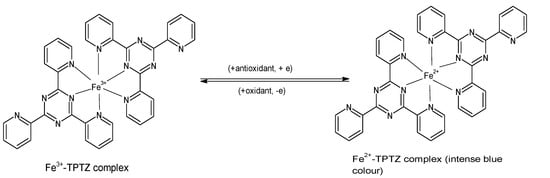
2.3. Ferric Reducing Antioxidant Power (FRAP)
2.4. Prooxidant Metal Chelation
2.5. Inhibition of Lipid Peroxidation including Lipoprotein Oxidation
3. Cell-Based Antioxidant Assays
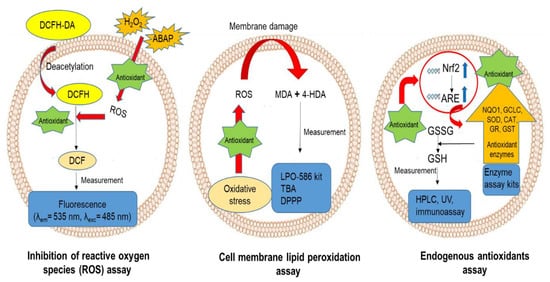
3.1. Inhibition of Intracellular ROS Production
| Ingredient | Cell-based Antioxidant Test | Cell Model | Antioxidant Effect | Reference |
|---|---|---|---|---|
| Lectin-free common bean (Phaseolus vulgaris L.) | CAA, haemolysis and antimutagenesis assay | Human erythrocytes, Saccharomyces cerevisiae | High dose-dependent antioxidant effect in yeast and human cells; attenuation of mutation induced by H2O2. | [59] |
| Turmeric leaf extract | CAA, lipid peroxidation and cell viability | Vero cells and zebrafish embryo model | Nuclear condensation, inhibition of ROS generation, cell death and lipid peroxidation. | [54] |
| Regular-darkening (RR) cranberry | Antioxidant enzyme activity | Caco-2 cells | Low permeability of flavanols; main influence was by surface adsorption and changes in cell signaling. Increase in level of SOD, CAT, GPx, GR, and GSH. | [66] |
| Fresh eating citrus fruits after in vitro digestion | CAA, cellular uptake assay | HepG2 cells | Digesta showed higher antioxidant activity than extract; strong correlation between naringenin and β-carotene absorption and antioxidant activity; detection of phenolic acids with hydroxybenzoic structure in cells showing permeability and no detection of hydroxycinnamic structure. | [67] |
| Flavonoid luteolin | GSH content, SOD activity, expression of antioxidant responsive-element (ARE) and Nrf2 | Caco-2 cells | Increase in expression of ARE and Nrf2; higher level of GSH and SOD inside the cells. | [68] |
| Bryophyllum pinnatum leaf extracts | Lipid peroxidation, GSH content, SOD activity | Human red blood cells | Freeze-dried extract resulted in the lowest membrane destabilization, MDA formation, and highest GSH content and SOD activity. | [69] |
| Two bioactive peptides from brown rice hydrolysates | CAA and cell hemolysis | Human red blood cells | Dipeptide Leu-Tyr and tripeptide Tyr-Leu-Ala inhibited oxidation and decreased hemolysis rate. | [70] |
| Moroccan Zantaz honey rich in methyl syringate | CAA | Caco-2 cells | Methyl syringate containing more than 50% total polyphenols; antioxidant activity was mainly related to syringate and gallic acid contents. | [71] |
| Peptide fraction < 1 kDa from Dendrobium aphyllum | CAA in HepG2 cells, immune bioactivity in RAW 264.7 and cellular absorption in Caco2 cells | HepG2, Caco2, and RAW 264.7 cells | CAA in HepG2 cells was 63.46 µM quercetin equivalent/100 g of peptide; cytokine secretion increased in RAW 264.7 cells, uptake in Caco2 was 19.7–25.5%. | [72] |
| Milk-derived bioactive peptides | Lipid peroxidation, antioxidant enzyme and Nrf2 expression |
Caco-2 cells | Main antioxidant mechanism involved increase in Keep1-Nrfs expression; inhibition of lipid peroxidation. | [73] |
| WL15 peptide from cysteine and glycine-rich protein 2 | Lipid peroxidation, antioxidant enzyme activity and expression | Human erythrocyte and zebrafish embryos | Increase in SOD activity and gene expression of glutathione S-transferase, glutathione peroxidase, and γ-glutamyl cysteine synthetase. Decrease in caspase 3 expression and MDA production. |
[52] |
| Corn gluten peptide fractions | CAA, antioxidant enzyme activity | HepG2 cells | High antioxidant effect was related to fraction < 1 and 1–3 kDa; increase in SOD, CAT, GR, and total GSH level. | [56] |
3.2. Inhibition of Cell Membrane Lipid Peroxidation
3.3. Activation of the Endogenous Antioxidant System
References
- Verbon, E.H.; Post, J.A.; Boonstra, J. The influence of reactive oxygen species on cell cycle progression in mammalian cells. Gene 2012, 511, 1–6.
- Wang, L.; Ding, L.; Xue, C.; Ma, S.; Du, Z.; Zhang, T.; Liu, J. Corn gluten hydrolysate regulates the expressions of antioxidant defense and ROS metabolism relevant genes in H2O2-induced HepG2 cells. J. Funct. Foods. 2018, 42, 362–370.
- Harnedy, P.A.; O’Keeffe, M.B.; Fitzgerald, R.J. Fractionation and identification of antioxidant peptides from an enzymatically hydrolysed Palmaria palmata protein isolate. Food Res. Int. 2017, 100, 416–422.
- Wu, J.; Huo, J.; Huang, M.; Zhao, M.; Luo, X.; Sun, B. Structural characterization of a tetrapeptide from sesame flavor-type Baijiu and its preventive effects against AAPH-induced oxidative stress in HepG2 Cells. J. Agric. Food Chem. 2017, 65, 10495.
- Görlach, A.; Dimova, E.Y.; Petry, A.; Martínez-Ruiz, A.; Hernansanz-Agustín, P.; Rolo, A.P.; Palmeira, C.M.; Kietzmann, T. Reactive oxygen species, nutrition, hypoxia and diseases: Problems solved? Redox. Biol. 2015, 6, 372–385.
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126.
- Aluko, R.E. Bioactive peptides. In Functional Foods and Nutraceuticals; Springer Science+Business Media LLC: New York, NY, USA, 2012; pp. 36–71.
- Nwachukwu, I.D.; Aluko, R.E. Structural and functional properties of food protein-derived antioxidant peptides. J. Food Biochem. 2019, 43, e12761.
- Lim, C.S.H.; Lim, S.L. Ferric reducing capacity versus ferric reducing antioxidant power for measuring total antioxidant capacity. Lab. Med. 2013, 44, 51–55.
- Shahidi, F.; Zhong, Y. Measurement of antioxidant activity. J. Funct. Foods. 2015, 18, 757–781.
- Nwachukwu, I.D.; Aluko, R.E. Antioxidant properties of flaxseed protein hydrolysates: Influence of hydrolytic enzyme concentration and peptide size. J. Am. Oil Chem. Soc. 2018, 95, 1105–1118.
- Qiu, X.; Chen, S.; Dong, S. Effects of silver carp antioxidant peptide on the lipid oxidation of sierra fish fillets (Scomberomorus niphonius) during frozen storage. J. Food Biochem. 2014, 38, 167–174.
- He, R.; Ju, X.; Yuan, J.; Wang, L.; Girgih, A.T.; Aluko, R.E. Antioxidant activities of rapeseed peptides produced by solid state fermentation. Food Res. Int. 2012, 49, 432–438.
- Bibi Sadeer, N.; Montesano, D.; Albrizio, S.; Zengin, G.; Mahomoodally, M.F. The versatility of antioxidant assays in food science and safety-chemistry, applications, strengths, and limitations. Antioxidants 2020, 9, 709.
- Romulo, A. The principle of some in vitro antioxidant activity methods: Review. IOP Conf. Ser. Earth Environ. Sci. 2020, 426, 12177.
- Udenigwe, C.C.; Udechukwu, M.C.; Yiridoe, C.; Gibson, A.; Gong, M. Antioxidant mechanism of potato protein hydrolysates against in vitro oxidation of reduced glutathione. J. Funct. Foods. 2016, 20, 195–203.
- Schaich, K.M.; Tian, X.; Xie, J. Hurdles and pitfalls in measuring antioxidant efficacy: A critical evaluation of ABTS, DPPH, and ORAC assays. J. Funct. Foods. 2015, 14, 111–125.
- Huang, D.; Ou, B.; Prior, R. The chemistry behind antioxidant capacity assays. J. Agric. Food Chem. 2005, 53, 1841–1856.
- Platzer, M.; Kiese, S.; Herfellner, T.; Schweiggert-Weisz, U.; Miesbauer, O.; Eisner, P. Common trends and differences in antioxidant activity analysis of phenolic substances using single electron transfer based assays. Molecules 2021, 26, 1244.
- Radi, R. Oxygen radicals, nitric oxide, and peroxynitrite: Redox pathways in molecular medicine. Proc. Natl. Acad. Sci. USA 2018, 115, 5839–5848.
- Dawidowicz, A.L.; Wianowska, D.; Olszowy, M. On practical problems in estimation of antioxidant activity of compounds by DPPH method (Problems in estimation of antioxidant activity). Food Chem. 2012, 131, 1037–1043.
- Berker, K.I.; Demirata, B.; Apak, R. Determination of total antioxidant capacity of lipophilic and hydrophilic antioxidants in the same solution by using ferric–ferricyanide assay. Food Anal. Methods 2012, 5, 1150–1158.
- Moon, J.-K.; Shibamoto, T. Antioxidant assays for plant and food components. J. Agric. Food Chem. 2009, 57, 1655–1666.
- Aksoy, L.; Kolay, E.; Ağılönü, Y.; Aslan, Z.; Kargıoğlu, M. Free radical scavenging activity, total phenolic content, total antioxidant status, and total oxidant status of endemic Thermopsis turcica. Saudi J. Biol. Sci. 2013, 20, 235–239.
- Kedare, S.; Singh, R. Genesis and development of DPPH method of antioxidant assay. J. Food Sci. Technol. 2011, 48, 412–422.
- Foti, M.C.; Daquino, C.; Geraci, C. Electron-transfer reaction of cinnamic acids and their methyl esters with the DPPH• radical in alcoholic solutions. J. Org. Chem. 2004, 69, 2309–2314.
- Gupta, S.S.; Ghosh, M. In vitro antioxidative evaluation of α- and β-Carotene, isolated from crude palm oil. J. Anal. Methods Chem. 2013, 2013, 351671.
- Foti, M.C. Use and abuse of the DPPH• radical. J. Agric. Food Chem. 2015, 63, 8765–8776.
- Phaniendra, A.; Jestadi, D.B.; Periyasamy, L. Free radicals: Properties, sources, targets, and their implication in various diseases. Indian J. Clin. Biochem. 2015, 30, 11–26.
- Onuh, J.O.; Girgih, A.T.; Aluko, R.E.; Aliani, M. In vitro antioxidant properties of chicken skin enzymatic protein hydrolysates and membrane fractions. Food Chem. 2014, 150, 366–373.
- Li, H.; Fan, Y.; Zhi, H.; Zhu, Y.; Liu, Y.; Wang, Y. Influence of fruit stalk on reactive oxygen species metabolism and quality maintenance of peach fruit under chilling injury condition. Postharvest Biol. Technol. 2019, 148, 141–150.
- de Avellar, I.G.J.; Magalhães, M.M.M.; Silva, A.B.; Souza, L.L.; Leitão, A.C.; Hermes-Lima, M. Reevaluating the role of 1,10- phenanthroline in oxidative reactions involving ferrous ions and DNA damage. Biochim. Biophys. Acta Gen. Subj. 2004, 1675, 46–53.
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76.
- Benzie, I.F.F. An automated, specific, spectrophotometric method for measuring ascorbic acid in plasma (EFTSA). Clin. Biochem. 1996, 29, 111–116.
- Sethi, S.; Joshi, A.; Arora, B.; Bhowmik, A.; Sharma, R.R.; Kumar, P. Significance of FRAP, DPPH, and CUPRAC assays for antioxidant activity determination in apple fruit extracts. Eur. Food Res. Technol. 2020, 246, 591–598.
- Rezaie, M.; Farhoosh, R.; Sharif, A.; Asili, J.; Iranshahi, M. Chemical composition, antioxidant and antibacterial properties of Bene (Pistacia atlantica subsp. mutica) hull essential oil. J. Food Sci. Technol. 2015, 52, 6784–6790.
- Wang, Y.-K.; Zhang, X.; Chen, G.-L.; Yu, J.; Yang, L.-Q.; Gao, Y.-Q. Antioxidant property and their free, soluble conjugate and insoluble-bound phenolic contents in selected beans. J. Funct. Foods. 2016, 24, 359–372.
- Aryal, S.; Baniya, M.K.; Danekhu, K.; Kunwar, P.; Gurung, R.; Koirala, N. Total phenolic content, flavonoid content and antioxidant potential of wild vegetables from Western Nepal. Plants 2019, 8, 96.
- Rocchetti, G.; Lucini, L.; Rodriguez, J.M.L.; Barba, F.J.; Giuberti, G. Gluten-free flours from cereals, pseudocereals and legumes: Phenolic fingerprints and in vitro antioxidant properties. Food Chem. 2019, 271, 157–164.
- Pulido, R.; Bravo, L.; Saura-Calixto, F. Antioxidant activity of dietary polyphenols as determined by a modified ferric reducing/antioxidant power assay. J. Agric. Food Chem. 2000, 48, 3396–3402.
- Udenigwe, C.C.; Aluko, R.E. Chemometric analysis of the amino acid requirements of antioxidant food protein hydrolysates. Int. J. Mol. Sci. 2011, 12, 3148–3161.
- Pownall, T.L.; Udenigwe, C.C.; Aluko, R.E. Amino acid composition and antioxidant properties of pea seed (Pisum sativum L.) enzymatic protein hydrolysate fractions. J. Agric. Food Chem. 2010, 58, 4712–4718.
- Berker, K.I.; Güçlü, K.; Demirata, B.; Apak, R. A novel antioxidant assay of ferric reducing capacity measurement using ferrozine as the colour forming complexation reagent. Anal. Methods 2010, 2, 1770.
- Işıl Berker, K.; Güçlü, K.; Tor, İ.; Demirata, B.; Apak, R. Total antioxidant capacity assay using optimized ferricyanide/prussian blue method. Food Anal. Methods 2010, 3, 154–168.
- Cherrak, S.A.; Mokhtari-Soulimane, N.; Berroukeche, F.; Bensenane, B.; Cherbonnel, A.; Merzouk, H.; Elhabiri, M. In vitro antioxidant versus metal ion chelating properties of flavonoids: A structure-activity investigation. PLoS ONE 2016, 11, e0165575.
- Adjimani, J.P.; Asare, P. Antioxidant and free radical scavenging activity of iron chelators. Toxicol. Rep. 2015, 2, 721–728.
- Xie, Z.; Huang, J.; Xu, X.; Jin, Z. Antioxidant activity of peptides isolated from alfalfa leaf protein hydrolysate. Food Chem. 2008, 111, 370–376.
- Girotti, A.W. Lipid hydroperoxide generation, turnover, and effector action in biological systems. J. Lipid Res. 1998, 39, 1529–1542.
- Girgih, A.; Udenigwe, C.; Aluko, R. Reverse-phase HPLC separation of hemp seed (Cannabis sativa L.) protein hydrolysate produced peptide fractions with enhanced antioxidant capacity. Plant Foods Hum. Nutr. 2013, 68, 39–46.
- Zhang, H.; Liu, R.; Tsao, R. Anthocyanin-rich phenolic extracts of purple root vegetables inhibit pro-inflammatory cytokines induced by H2O2 and enhance antioxidant enzyme activities in Caco-2 cells. J. Funct. Foods. 2016, 22, 363–375.
- López-Alarcón, C.; Denicola, A. Evaluating the antioxidant capacity of natural products: A review on chemical and cellular-based assays. Anal. Chim. Acta. 2013, 763, 1–10.
- Guru, A.; Lite, C.; Freddy, A.J.; Issac, P.K.; Pasupuleti, M.; Saraswathi, N.; Arasu, M.V.; Al-Dhabi, N.A.; Arshad, A.; Arockiaraj, J. Intracellular ROS scavenging and antioxidant regulation of WL15 from cysteine and glycine-rich protein 2 demonstrated in zebrafish in vivo model. Dev. Comp. Immunol. 2021, 114, 103863.
- Grzesik, M.; Naparło, K.; Bartosz, G.; Sadowska-Bartosz, I. Antioxidant properties of catechins: Comparison with other antioxidants. Food Chem. 2018, 241, 480–492.
- Kim, S.; Kim, M.; Kang, M.-C.; Lee, H.H.L.; Cho, C.H.; Choi, I.; Park, Y.; Lee, S.-H. Antioxidant effects of turmeric leaf extract against hydrogen peroxide-induced oxidative stress in vitro in vero cells and in vivo in zebrafish. Antioxidants 2021, 10, 112.
- Wolfe, K.L.; Liu, R.H. Cellular antioxidant activity (CAA) assay for assessing antioxidants, foods, and dietary supplements. J. Agric. Food Chem. 2007, 55, 8896–8907.
- Wang, L.; Ding, L.; Yu, Z.; Zhang, T.; Ma, S.; Liu, J. Intracellular ROS scavenging and antioxidant enzyme regulating capacities of corn gluten meal-derived antioxidant peptides in HepG2 cells. Food Res. Int. 2016, 90, 33–41.
- Ibrahim, H.R.; Nanbu, F.; Miyata, T. Potent antioxidant peptides derived from honey major protein enhance tolerance of eukaryotic cells toward oxidative stress. Food Prod. Process. Nutr. 2021, 3, 11.
- Kellett, M.E.; Greenspan, P.; Pegg, R.B. Modification of the cellular antioxidant activity (CAA) assay to study phenolic antioxidants in a Caco-2 cell line. Food Chem. 2018, 244, 359–363.
- Frassinetti, S.; Gabriele, M.; Caltavuturo, L.; Longo, V.; Pucci, L. Antimutagenic and antioxidant activity of a selected lectin-free common bean (Phaseolus vulgaris L.) in two cell-based models. Plant Foods Hum. Nutr. 2015, 70, 35–41.
- Wolfe, K.L.; Kang, X.; He, X.; Dong, M.; Zhang, Q.; Liu, R.H. Cellular antioxidant activity of common fruits. J. Agric. Food Chem. 2008, 56, 8418–8426.
- Dong, W.; Chen, D.; Chen, Z.; Sun, H.; Xu, Z. Antioxidant capacity differences between the major flavonoids in cherry (Prunus pseudocerasus) in vitro and in vivo models. LWT Food Sci. Technol. 2021, 141, 110938.
- Sato, Y.; Itagaki, S.; Kurokawa, T.; Ogura, J.; Kobayashi, M.; Hirano, T.; Sugawara, M.; Iseki, K. In vitro and in vivo antioxidant properties of chlorogenic acid and caffeic acid. Int. J. Pharm. 2011, 403, 136–138.
- Nallathambi, R.; Poulev, A.; Zuk, J.B.; Raskin, I. Proanthocyanidin-rich grape seed extract reduces inflammation and oxidative stress and restores tight junction barrier function in Caco-2 colon cells. Nutrients 2020, 12, 1623.
- Pan, Y.; Deng, Z.-Y.; Chen, X.; Zhang, B.; Fan, Y.; Li, H. Synergistic antioxidant effects of phenolic acids and carotenes on H2O2-induced H9c2 cells: Role of cell membrane transporters. Food Chem. 2021, 341, 128000.
- Wongaem, A.; Reamtong, O.; Srimongkol, P.; Sangtanoo, P.; Saisavoey, T.; Karnchanatat, A. Antioxidant properties of peptides obtained from the split gill mushroom (Schizophyllum commune). J. Food Sci. Technol. 2021, 58, 680–691.
- Yang, C.; Shahidi, F.; Tsao, R. Biomarkers of oxidative stress and cellular-based assays of indirect antioxidant measurement. In Measurement of Antioxidant Activity & Capacity: Recent Trends and Applications; Apak, R., Capanoglu, E., Shahidi, F., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2018; pp. 165–186.
- Reyes-Gonzales, M.C.; Esteban-Zubero, E.; López-Pingarrón, L.; Soria, M.S.; Pereboom, D.; Tan, D.X.; Reiter, R.J.; García, J.J. Antioxidant activity of pineal methoxyindoles on hepatocyte plasmatic membrane. Melatonin Res. 2019, 2, 161–174.
- Shi, Y.; Kovacs-Nolan, J.; Jiang, B.; Tsao, R.; Mine, Y. Peptides derived from eggshell membrane improve antioxidant enzyme activity and glutathione synthesis against oxidative damage in Caco-2 cells. J. Funct. Foods. 2014, 11, 571–580.
- Kang, M.-C.; Cha, S.H.; Wijesinghe, W.; Kang, S.-M.; Lee, S.-H.; Kim, E.-A.; Song, C.B.; Jeon, Y.-J. Protective effect of marine algae phlorotannins against AAPH-induced oxidative stress in zebrafish embryo. Food Chem. 2013, 138, 950–955.
- Kim, Y.-S.; Kim, E.-K.; Jeon, N.-J.; Ryu, B.-I.; Hwang, J.-W.; Choi, E.-J.; Moon, S.-H.; Jeon, B.-T.; Park, P.-J. Antioxidant effect of taurine-rich paroctopus dofleini extracts through inhibiting ROS production against LPS-induced oxidative stress in vitro and in vivo model. Adv. Exp. Med. Biol. 2017, 975, 1165–1177.
- Nishikawa, H.; Tsutsumi, J.; Kitani, S. Anti-inflammatory and anti-oxidative effect of curcumin in connective tissue type mast cell. J. Funct. Foods. 2013, 5, 763–772.
- Cropotova, J.; Rustad, T. A new fluorimetric method for simultaneous determination of lipid and protein hydroperoxides in muscle foods with the use of diphenyl-1-pyrenylphosphine (DPPP). LWT Food Sci. Technol. 2020, 128, 109467.
- Zhang, Q.; Tong, X.; Sui, X.; Wang, Z.; Qi, B.; Li, Y.; Jiang, L. Antioxidant activity and protective effects of Alcalase-hydrolyzed soybean hydrolysate in human intestinal epithelial Caco-2 cells. Food Res. Int. 2018, 111, 256–264.
- Bartolomei, M.; Bollati, C.; Bellumori, M.; Cecchi, L.; Cruz-Chamorro, I.; Santos-Sánchez, G.; Ranaldi, G.; Ferruzza, S.; Sambuy, Y.; Arnoldi, A. Extra virgin olive oil phenolic extract on human hepatic HepG2 and intestinal caco-2 cells: Assessment of the antioxidant activity and intestinal trans-epithelial transport. Antioxidants 2021, 10, 118.
- Cilla, A.; Rodrigo, M.J.; Zacarías, L.; De Ancos, B.; Sánchez-Moreno, C.; Barberá, R.; Alegría, A. Protective effect of bioaccessible fractions of citrus fruit pulps against H2O2-induced oxidative stress in Caco-2 cells. Food Res. Int. 2018, 103, 335–344.
- Martins, C.C.; Bagatini, M.D.; Simões, J.L.B.; Cardoso, A.M.; Baldissarelli, J.; Dalenogare, D.P.; Dos Santos, D.L.; Schetinger, M.R.C.; Morsch, V.M. Increased oxidative stress and inflammatory markers contrasting with the activation of the cholinergic anti-inflammatory pathway in patients with metabolic syndrome. Clin. Biochem. 2021, 89, 63–69.
- Armstrong, D. Oxidative Stress Biomarkers and Antioxidant Protocols; Humana Press: Totowa, NJ, USA, 2002.
- Xie, C.; Kang, J.; Ferguson, M.E.; Nagarajan, S.; Badger, T.M.; Wu, X. Blueberries reduce pro-inflammatory cytokine TNF-α and IL-6 production in mouse macrophages by inhibiting NF-κB activation and the MAPK pathway. Mol. Nutr. Food Res. 2011, 55, 1587–1591.
- Biswas, S.K.; McClure, D.; Jimenez, L.A.; Megson, I.L.; Rahman, I. Curcumin induces glutathione biosynthesis and inhibits NF-κB activation and interleukin-8 release in alveolar epithelial cells: Mechanism of free radical scavenging activity. Antioxid. Redox Signal. 2005, 7, 32–41.
- Tonolo, F.; Folda, A.; Cesaro, L.; Scalcon, V.; Marin, O.; Ferro, S.; Bindoli, A.; Rigobello, M.P. Milk-derived bioactive peptides exhibit antioxidant activity through the Keap1-Nrf2 signaling pathway. J. Funct. Foods. 2020, 64, 103696.
- Chen, P.X.; Zhang, H.; Marcone, M.F.; Pauls, K.P.; Liu, R.; Tang, Y.; Zhang, B.; Renaud, J.B.; Tsao, R. Anti-inflammatory effects of phenolic-rich cranberry bean (Phaseolus vulgaris L.) extracts and enhanced cellular antioxidant enzyme activities in Caco-2 cells. J. Funct. Foods. 2017, 38, 675–685.
- Liu, H.; Ye, H.; Sun, C.; Xi, H.; Ma, J.; Lai, F.; Wu, H. Antioxidant activity in HepG2 cells, immunomodulatory effects in RAW 264.7 cells and absorption characteristics in Caco-2 cells of the peptide fraction isolated from Dendrobium aphyllum. Int. J. Food Sci. Technol. 2018, 53, 2027–2036.
- Sun, Y.; Tao, W.; Huang, H.; Ye, X.; Sun, P. Flavonoids, phenolic acids, carotenoids and antioxidant activity of fresh eating citrus fruits, using the coupled in vitro digestion and human intestinal HepG2 cells model. Food Chem. 2019, 279, 321–327.
- Yuan, J.; Che, S.; Zhang, L.; Ruan, Z. Reparative effects of ethanol-induced intestinal barrier injury by flavonoid luteolin via MAPK/NF-κB/MLCK and Nrf2 signaling pathways. J. Agric. Food Chem. 2021, 69, 4101–4110.
- Omojokun, O.S.; Oboh, G.; Ademiluyi, A.O.; Oladele, J.O.; Boligon, A.A. Impact of drying processes on Bryophyllum pinnatum phenolic constituents and its anti-inflammatory and antioxidative activities in human erythrocytes. J. Food Biochem. 2020, 45, e13298.
- Du, R.; Liu, K.; Zhao, S.; Chen, F. Changes in antioxidant activity of peptides identified from brown rice hydrolysates under different conditions and their protective effects against AAPH-induced oxidative stress in human erythrocytes. ACS Omega 2020, 5, 12751–12759.
- Elamine, Y.; Lyoussi, B.; Miguel, M.G.; Anjos, O.; Estevinho, L.; Alaiz, M.; Girón-Calle, J.; Martín, J.; Vioque, J. Physicochemical characteristics and antiproliferative and antioxidant activities of Moroccan Zantaz honey rich in methyl syringate. Food Chem. 2021, 339, 128098.

